伏隔核區Jak/Stat3抑制劑注射調控布氏田鼠的能量代謝
2018-01-30 01:58:00閆立新張知彬宋銘晶
四川動物 2018年1期
關鍵詞:實驗
閆立新, 張知彬, 宋銘晶*
(1.中國醫學科學院醫學實驗動物研究所,北京協和醫學院比較醫學中心,衛生部人類疾病比較醫學重點實驗室,北京100021; 2.中國科學院動物研究所,北京100101)
瘦素主要由白色脂肪細胞分泌,它有助于減少脂肪積累、降低體質量、增強能量代謝。瘦素缺乏或瘦素抵抗是導致肥胖的重要發病機制之一。迄今為止,已經發現瘦素有8種突變方式,突變的瘦素蛋白不能和瘦素受體結合,無法發揮減少攝食和降低體質量的作用,最終導致極度肥胖(Strobeletal.,1998;Chekhranovaetal.,2008;Mazenetal.,2009;Fischer-Posovszkyetal.,2010;Fatimaetal.,2011;Thakuretal.,2014;Wabitschetal.,2015)。最近發現的瘦素的一種突變是在2015年,研究者發現一個2歲小男孩出現了早發性極端肥胖,原因是其瘦素的第100個氨基酸由天冬氨酸突變為酪氨酸,瘦素失去了生物學活性,在使用人重組瘦素(美曲普汀)治療后,病人的飲食行為恢復正常,且體質量迅速下降(Wabitschetal.,2015)。瘦素是下丘腦發育過程中的神經營養因子,促進下丘腦瘦素傳導到大腦腦區通路的形成,從而達到控制腦區、調控飲食與能量消耗的功能(Bouretetal.,2004)。信號轉導與轉錄激活因子3(signal transducer and activator of transcription,Stat3)作為轉錄因子可以通過調控基因轉錄來控制細胞增殖、生長、凋亡等過程,進而參與到免疫、腫瘤生成、肥胖等生命進程中。瘦素通過誘導Stat3的磷酸化調控中樞神經系統,從而達到抑制肥胖的功能(Pattersonetal.,2009)。研究發現,高脂飲食肥胖小鼠的leptin-Stat3(瘦素-Stat3)信號通路幾乎被抑制,磷酸化Stat3(P-Stat3)的表達量明顯下降,但經過12周自主轉輪運動實驗后,高脂飲食肥胖小鼠的leptin-Stat3信號通路恢復,P-Stat3表達量提高,進而減少高脂飲食對小鼠體質量的影響(Laingetal.,2016)。
布氏田鼠Lasiopodomysbrandtii是研究“健康性肥胖”的潛在模型動物,雖然長光照在35 d后會導致布氏田鼠體質量在短時間顯著增加,但是它會重新建立一種穩態,與短光照的布氏田鼠相比,長光照布氏田鼠白色脂肪的總含量顯著增高,血清瘦素含量顯著增高,飲食量顯著下降,沒有出現瘦素抵抗與胰島素抵抗的現象(Liuetal.,2016)。此外,在懷孕和哺乳期間給予布氏田鼠高蛋白飲食可以降低后代的消化能力,并降低其肥胖抵抗以及糖耐受現象(Louetal.,2015;Zhangetal.,2017)。因此,我們認為布氏田鼠在研究肥胖與代謝方面具有很高的潛在應用價值。迄今為止,沒有大腦伏隔核區與代謝類疾病相關的報道,而且瘦素在伏隔核區高表達,因此選擇伏隔核作為本實驗代謝研究的腦區。通過在布氏田鼠伏隔核區定點注射Jak/Stat3抑制劑(WP1066)阻斷leptin-Stat3信號通路,探索伏隔核區瘦素通路與能量代謝的關系。
1 材料和方法
1.1 材料
1.1.1實驗動物50 g左右雄性布氏田鼠(中國醫學科學院醫學實驗動物研究所,實驗動物使用批準號:ILAS-PG-2014-011)共30只,并在獨立送排風凈化動物籠中飼養,照明晝夜節律為14∶10。動物飲水、墊料和籠具均經高壓滅菌處理,飼料經Co60照射消毒達到SPF級動物飼料標準。
1.1.2實驗器材腦立體定位儀(Stoelting,USA),腦區套管(深圳市瑞沃德生命科技有限公司),TSE動物代謝測量分析系統-PhenoMaster/LabMaster(TSE Systems)。
1.1.3試劑Jak/Stat3抑制劑WP1066(Abcam),二甲基亞砜(DMSO),Western Blot用瘦素抗體(Abcam),免疫組化用Phospho-Stat3(Tyr705)抗體(Cell Signaling),山羊抗兔抗體(北京中杉金橋生物技術有限公司),麻醉劑(2.5%水合氯醛)。
1.2 實驗方法
1.2.1腦區埋管手術用麻醉劑將布氏田鼠麻醉后固定在腦立體定位儀上,在頭部剪開1 cm左右露出頭蓋骨,并確定左右頭骨、前后囟在同一水平線上,根據伏隔核腦區坐標(AP:1.6 mm,ML:±1.3 mm,DV:-4.3 mm)雙側埋管。手術后,給予正常飲食。1周后,待其體質量恢復后進行藥物注射。
1.2.2腦區注射對照組注射二甲基亞砜,實驗組注射WP1066(10 mmol·L-1),注射速度是每側1 μL/3 min,注射結束后留針2 min,防止液體倒流并使其充分擴散。每隔一天注射1次藥物,共注射4次。
1.2.3動物代謝測量分析實驗先將布氏田鼠放入代謝籠適應2 d,再收集其之后12 d的本底代謝水平,然后開始給布氏田鼠注射藥物,注射藥物結束4 h后收集接下來2 d的實驗數據。
1.2.4動物體質量在注射藥物之前稱量布氏田鼠的體質量,再稱量其第10天的體質量。
1.2.5WesternBlot實驗實驗結束后,獲取布氏田鼠新鮮腦組織,冰凍切片摳取伏隔核腦區,將摳取的腦組織放在同一個離心管里,提取蛋白,-80 ℃儲存,Western Blot檢測瘦素的表達量。
1.2.6免疫組化實驗實驗結束后,在布氏田鼠的心臟處灌流,先用生理鹽水沖洗,再用4%多聚甲醛灌注固定,最后將腦組織儲存在4%多聚甲醛中,冰凍切片厚40 μm,浸泡在10%疊氮化鈉中儲存,免疫組化實驗檢測P-Stat3表達量。
1.2.7血清瘦素濃度和血糖濃度測定實驗結束后麻醉布氏田鼠,用1 mL注射器在其腹主動脈抽取血樣放在血液促凝管中,離心得血清(3 500 r·min-1,10 min),并檢測血清瘦素濃度(放射免疫,北京北方生物技術研究所有限公司)和血糖濃度(葡萄糖氧化法,北京北方生物技術研究所有限公司)。
1.3 數據分析
數據統計使用SPSS 20.0,代謝籠數據(總飲水量、總能量消耗量、總耗氧量、呼吸交換率)用重復測量方差分析,其他數據用獨立樣本T檢驗法分析,P<0.05表示差異有統計學意義,數據以平均值±標準誤(Mean±SE)表示,最后用GraphPad Prism 6作圖。
2 實驗結果
2.1 免疫組化檢測伏隔核區P-Stat3表達
免疫組化結果顯示,布氏田鼠伏隔核區P-Stat3(Tyr705)表達顯著下降(圖1)。
2.2 Western Blot實驗檢測伏隔核區瘦素表達
Western Blot結果顯示,布氏田鼠伏隔核區瘦素表達水平顯著下降(圖2)。
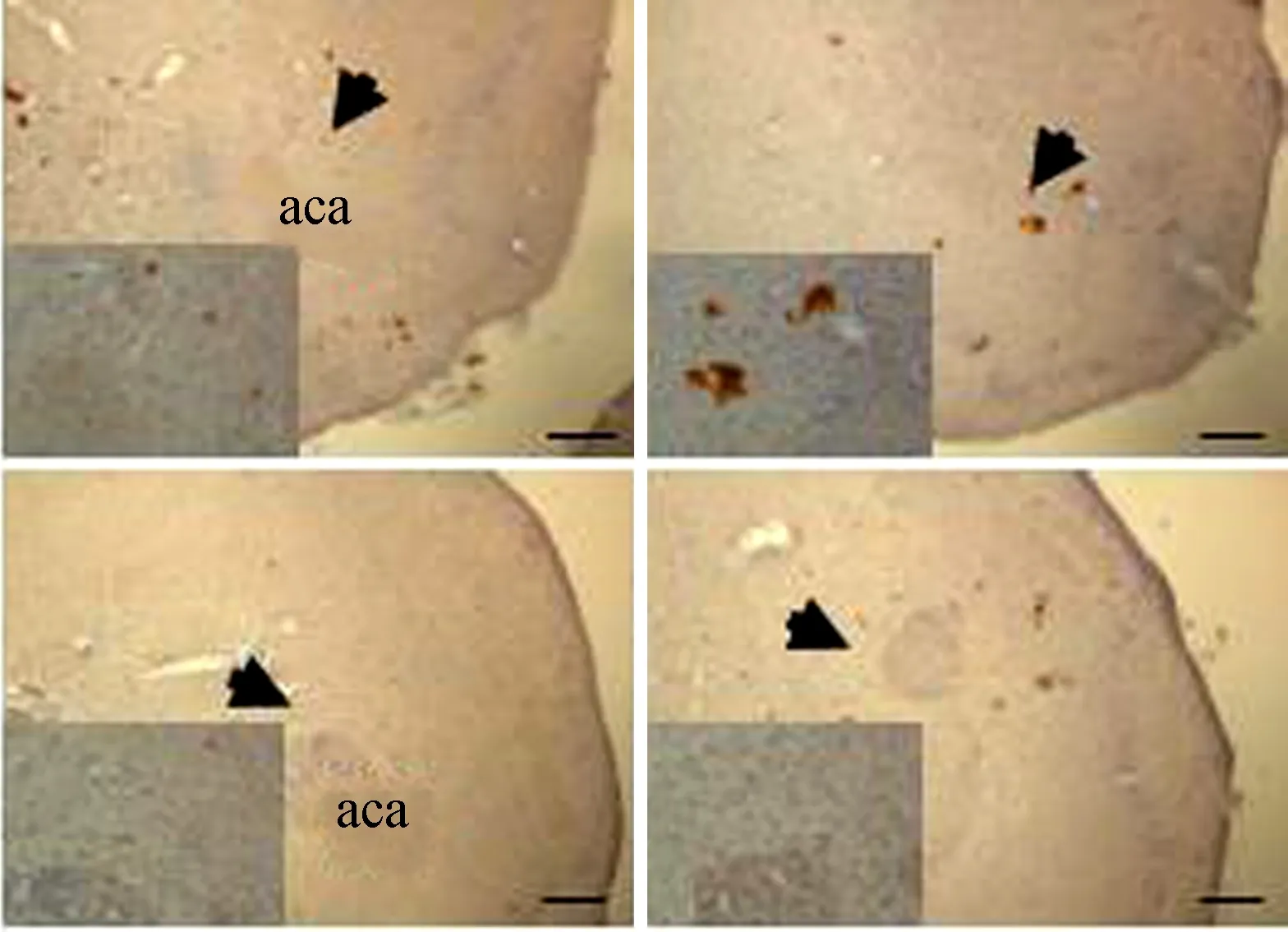
圖1 對照組(上)與實驗組(下)伏隔核區P-Stat3的表達Fig. 1 Protein expression of P-Stat3 in nucleus accumbens of control group (upper) and experimental group (bottom)
aca. 伏隔核區的一部分,用于伏隔核的定位; 左下角為箭頭所指處的放大; 比例尺=200 μm
aca. the core of nudeus accumbens, which is used to locate nudeus accumbens in the brain; the small pictures in the lower-left are amplified area directed by the arrow in the same figure; scale bar=200 μm
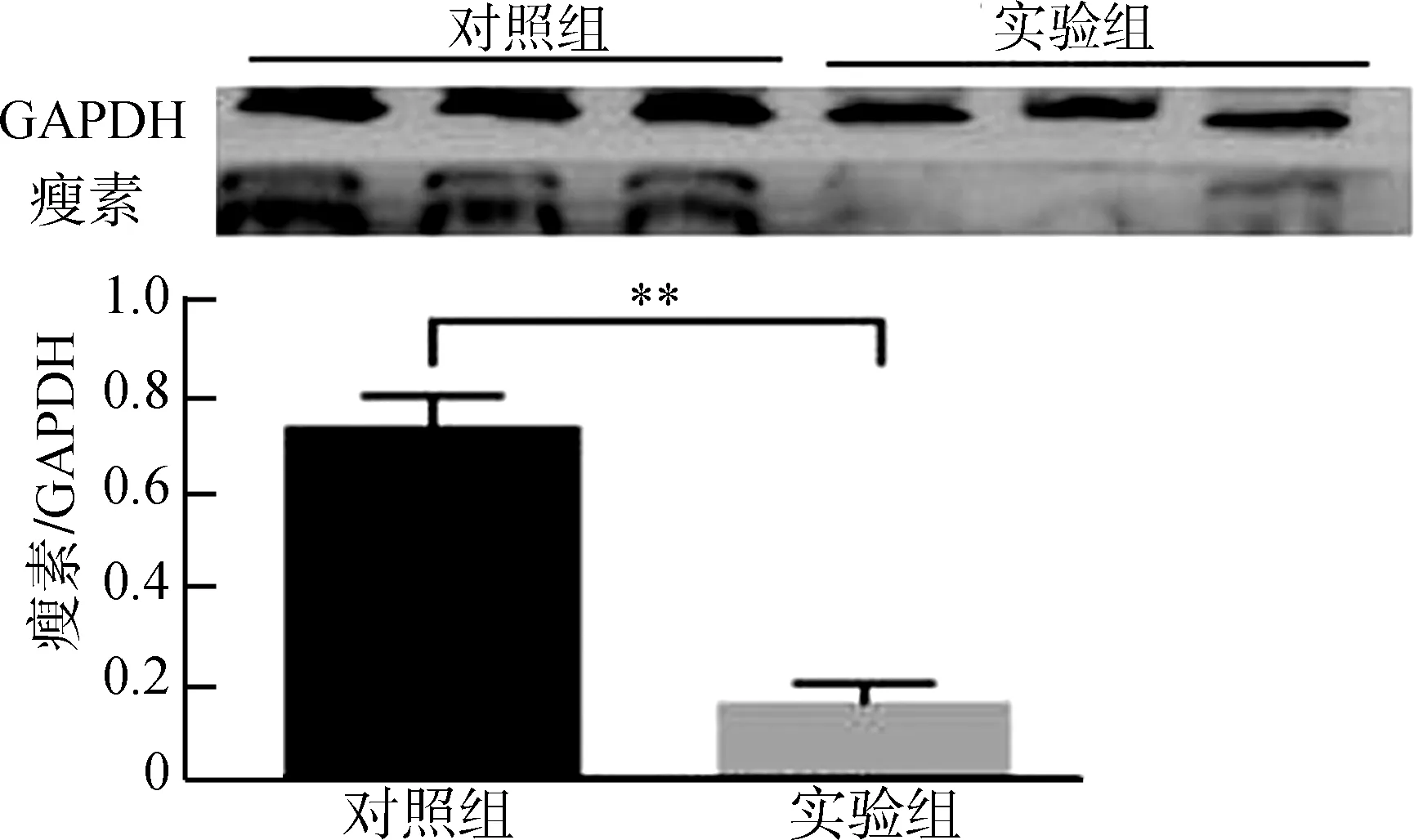
圖2 實驗組與對照組伏隔核區的瘦素表達量
Fig. 2 Protein expression of leptin in control group and experimental group
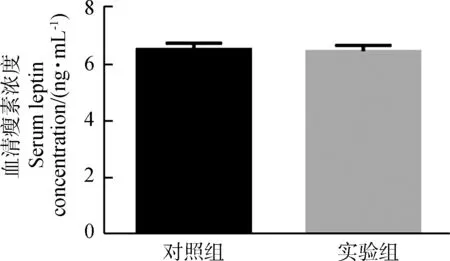
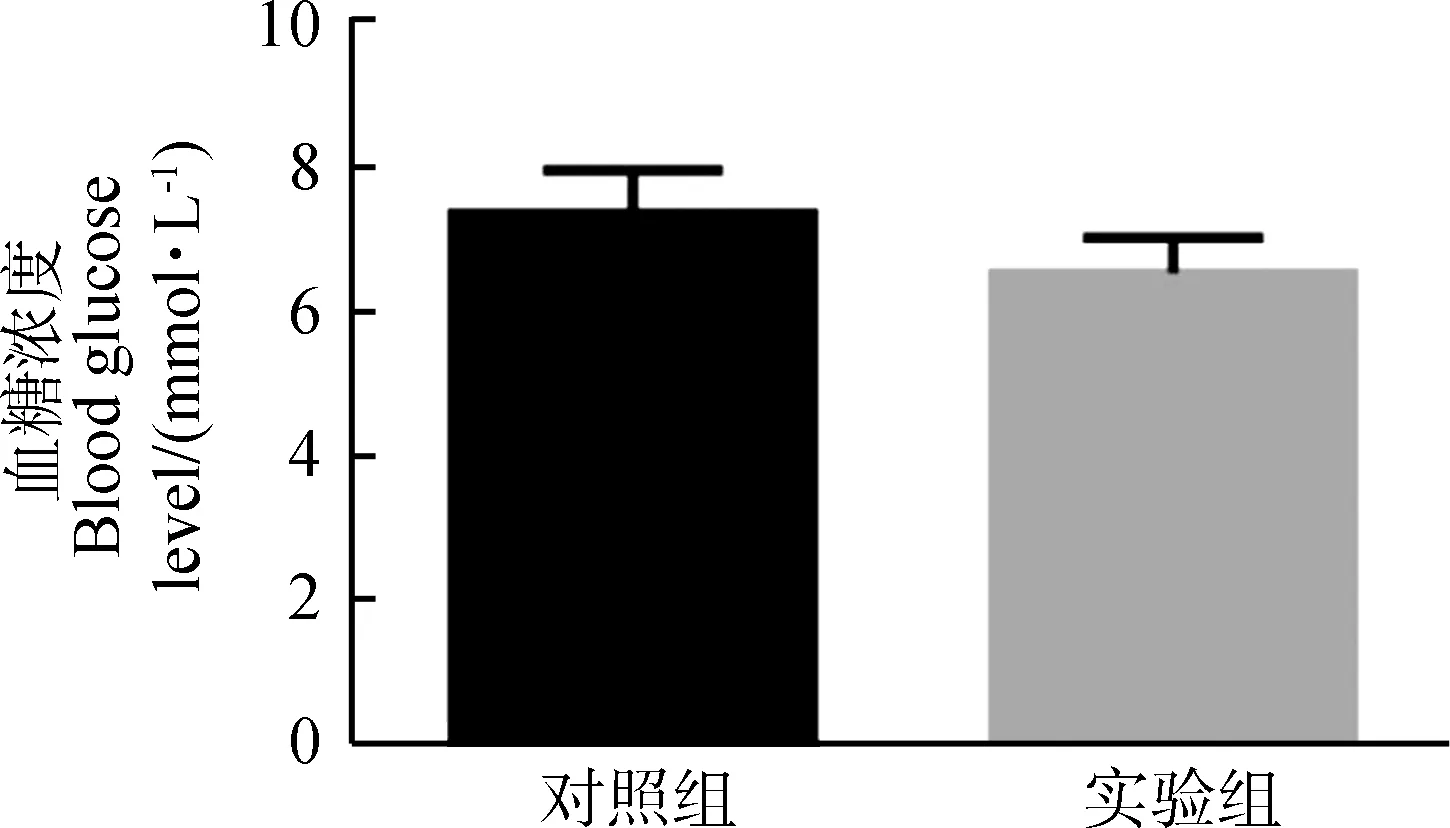
**0.001 實驗組與對照組血清瘦素濃度之間的差異無統計學意義(P>0.05,實驗組n=9,對照組n=8;圖3),血糖濃度差異也無統計學意義(P>0.05,實驗組n=7,對照組n=6;圖3)。 圖3 對照組和實驗組血清瘦素濃度與血糖濃度比較Fig. 3 Comparison of serum leptin concentration and blood glucose level between control group and experimental group 與對照組相比,實驗組布氏田鼠總飲水量極顯著增加(P<0.01,實驗組:n=6,對照組:n=6,時間=2 700 min;圖4:A)、總能量消耗量顯著下降(P<0.05,實驗組:n=6,對照組:n=6,時間=2 700 min;圖4:B)、總耗氧量顯著下降(P<0.05,實驗組:n=6,對照組:n=6,時間=2 700 min;圖4:C)、呼吸交換率(respiratory exchange rate, RER;RER=CO2產出量/O2消耗量)顯著提高(P<0.05,實驗組:n=6,對照組:n=6,時間=1 800 min;圖4:D)、總自主運動量顯著下降(P<0.05,實驗組:n=6,對照組:n=6,時間=2 700 min;圖4:E)、體質量顯著增加(P<0.05,實驗組:n=6,對照組:n=6,時間=10 d;圖4:F)。 圖4 對照組和實驗組的基礎代謝率比較 *0.01 呼吸交換率是相同時間內釋放CO2與吸收O2體積或摩爾數的比值,且有研究表明,O2消耗量也是影響體質量與脂肪儲存的機制之一。研究發現,高呼吸交換率表明脂肪的高氧化率以及碳水化合物的低氧化率(Schutz,1995);同時研究發現,呼吸交換率與體質量的增加有關,呼吸交換率提高會導致體質量增加(Haineretal.,2000;Marraetal.,2004)。本實驗結果表明,與對照組相比,實驗組布氏田鼠的呼吸交換率顯著提高,說明實驗組傾向于減少脂肪的消耗而增加糖的消耗,且實驗組的能量消耗、活動量以及耗氧量顯著下降,為脂肪積累創造了條件,最終導致肥胖。 在伏隔核區注射Jak/Stat3抑制劑后布氏田鼠體質量顯著增加,而血糖與血清瘦素水平卻沒有顯著變化,從而驗證了布氏田鼠可以作為研究“健康性肥胖”的模型動物(Liuetal.,2016)。實驗組布氏田鼠的飲水量顯著增加,由此可見,布氏田鼠是一種可應用于研究代謝與飲食的潛在實驗模型動物。 瘦素在控制能量代謝以及飲食功能上的重要性毋庸置疑,但是不同腦區瘦素水平的差異及其對代謝和神經調控的影響還不確定。目前的研究結果顯示,瘦素抵抗主要分為外周性瘦素抵抗和中樞性瘦素抵抗。本實驗研究發現,在布氏田鼠伏隔核區注射Jak/Stat3抑制劑后,P-Stat3表達量和瘦素水平顯著下降,而血清瘦素濃度卻無顯著變化,由此可見,使用抑制劑后只是導致中樞性瘦素抵抗,沒有造成外周性瘦素抵抗。造成中樞性瘦素抵抗的原因可能有2種:一是伏隔核區Jak/Stat3信號通路被抑制,導致leptin-Stat3信號通路傳導障礙,從而直接導致中樞性瘦素抵抗;二是伏隔核區Jak/Stat3信號通路被抑制后,間接降低血清瘦素的血腦屏障轉運功能,從而降低腦組織中瘦素水平,最終導致中樞性瘦素抵抗(Banks,2001)。由此可見,leptin-Stat3信號通路被抑制會導致中樞性瘦素抵抗,調控布氏田鼠伏隔核區的leptin-Stat3信號通路能夠調控基礎代謝,進而在短期內改變其體質量。需要進一步實驗證明瘦素和P-Stat3調控下游通路的具體機制。 致謝:感謝孫秀萍老師對實驗的指導,以及張楠師兄和田靜的幫助。 Banks WA. 2001. Enhanced leptin transport across the blood-brain barrier by alpha 1-adrenergic agents[J]. Brain Research, 899(1-2): 209-217. Bouret SG, Draper SJ, Simerly RB. 2004. Trophic action of leptin on hypothalamic neurons that regulate feeding[J]. Science, 304(5667): 108-110. Chekhranova MK, Karpova SK, Iatsyshina SB,etal. 2008. A new mutation c.422C>G (p.S141C) in homo- and heterozygous forms of the human leptin gene[J]. Bioorganicheskaia Khimiia, 34(6): 854-856. Fatima W, Shahid A, Imran M,etal. 2011. Leptin deficiency and leptin gene mutations in obese children from Pakistan[J]. International Journal of Pediatric Obesity, 6(5-6): 419-427. Fischer-Posovszky P, von Schnurbein J, Moepps B,etal. 2010. A new missense mutation in the leptin gene causes mild obesity and hypogonadism without affecting T cell responsiveness[J]. The Journal of Clinical Endocrinology & Metabolism, 95(6): 2836-2840. Hainer V, Kunesova M, Parizkova J,etal. 2000. Respiratory quotient in obesity: its association with an ability to retain weight loss and with parental obesity[J]. Sborník Lékarsky, 101(1): 99-104. Laing BT, Do K, Matsubara T,etal. 2016. Voluntary exercise improves hypothalamic and metabolic function in obese mice[J]. Journal of Endocrinology, 229(2): 109-122. Liu XY, Yang DB, Xu YC,etal. 2016. Photoperiod induced obesity in the Brandt’s vole (Lasiopodomysbrandtii): a model of 'healthy obesity?[J]. Disease Models & Mechanisms, 9(11): 1357-1366. Lou MF, Shen W, Fu RS,etal. 2015. Maternal dietary protein supplement confers long-term sex-specific beneficial consequences of obesity resistance and glucose tolerance to the offspring in Brandt’s voles[J]. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 182: 38-44. Marra M, Scalfi L, Contaldo F,etal. 2004. Fasting respiratory quotient as a predictor of long-term weight changes in non-obese women[J]. Annals Nutrition Metabolism, 48(3): 189-192. Mazen I, El-Gammal M, Abdel-Hamid M,etal. 2009. A novel homozygous missense mutation of the leptin gene (N103K) in an obese Egyptian patient[J]. Molecular Genetics Metabolism, 97(4): 305-308. Patterson CM, Bouret SG, Dunn-Meynell A,etal. 2009. Three weeks of postweaning exercise in DIO rats produces prolonged increases in central leptin sensitivity and signaling[J]. American Journal of Physiology Regulatory Integrative Comparative Physiology, 296(3): R537-R548. Schutz Y. 1995. Abnormalities of fuel utilization as predisposing to the development of obesity in humans[J]. Obesity, 3(S2): 173S-178S. Strobel A, Issad T, Camoin L,etal. 1998. A leptin missense mutation associated with hypogonadism and morbid obesity[J]. Nature Genetics, 18(3): 213-215. Thakur S, Kumar A, Dubey S,etal. 2014. A novel mutation of the leptin gene in an Indian patient[J]. Clinal Genetics, 86(4): 391-393. Wabitsch M, Funcke JB, Lennerz B,etal. 2015. Biologically inactive leptin and early-onset extreme obesity[J]. The New England Journal of Medicine, 372(1): 48-54. Zhang XY, Lou MF, Shen W,etal. 2017. A maternal low-fiber diet predisposes offspring to improved metabolic phenotypes in adulthood in an herbivorous rodent[J]. Physiological Biochemical Zoology, 90(1): 75-84.2.3 血清瘦素濃度與血糖濃度變化
2.4 代謝實驗
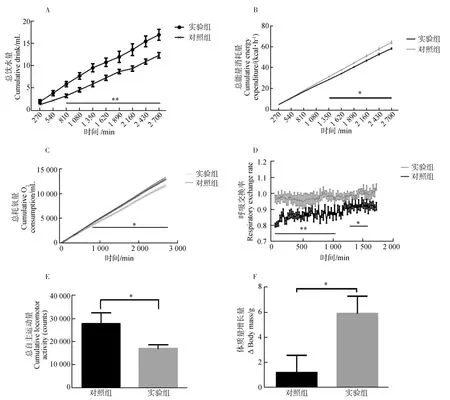
Fig.4 Comparison of basal metabolic rate between control group and experimental group3 討論
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00作文·小學低年級(2024年2期)2024-04-29 00:00:00作文·小學低年級(2023年3期)2023-04-29 00:00:00小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42小主人報(2022年4期)2022-08-09 08:52:06中學生數理化·中考版(2022年11期)2022-02-16 07:01:20小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50發明與創新(2016年38期)2016-08-22 03:02:52太空探索(2016年5期)2016-07-12 15:17:55
