小桐子MYB基因家族的鑒定及JcMYB308基因的克隆與低溫表達分析
2018-01-31 03:45:24辛胡顏岳輝丁雪梅劉潮高永代冬琴唐利洲王海波
生物技術(shù)通報 2018年1期
辛胡 顏岳輝 丁雪梅 劉潮 高永 代冬琴唐利洲 王海波
(1. 西南林業(yè)大學林學院,昆明 650224;2. 曲靖師范學院 云南高原生物資源保護與利用研究中心 云南省高校云貴高原動植物遺傳多樣性及生態(tài)適應性重點實驗室,曲靖 655011)
小桐子(Jatropha curcas L.)又名麻瘋樹、黃腫樹(廣東)、膏桐(云南)、桐油樹(臺灣)、假花生(廣西),發(fā)源于加勒比海的海島,現(xiàn)廣泛分布和栽培于熱帶、亞熱帶等地區(qū)[1-2],在我國主要分布于廣西西南部和西部、云南南部和東南部等地區(qū)[3]。它屬大戟科(Euphorbiaceae)麻瘋樹屬(Jatropha)多年生木本油料植物,其種子含油量高,流動性好,是加工生物柴油的優(yōu)質(zhì)原料,且耐干旱貧瘠,是干熱河谷地區(qū)理想的生態(tài)造林樹種[4-5],被世界公認為生物能源樹種。其油被改性后適用于各種柴油機,并在閃點、凝固點、硫含量、一氧化碳排量及顆粒值等關(guān)鍵技術(shù)均優(yōu)于國內(nèi)零號柴油,達到歐洲二號排放標準。另外,其種仁可作為傳統(tǒng)的肥皂及潤滑油原料[6],有瀉下和催吐功效[7-8],油麩可作農(nóng)藥及肥料[9],兼具經(jīng)濟價值及良好的開發(fā)前景[10]。
生物與非生物逆境脅迫下,植物通過一系列信號轉(zhuǎn)導途徑激活相應的轉(zhuǎn)錄因子,并結(jié)合靶基因上游特定順式作用元件從而激活或抑制抗逆相關(guān)基因以特定的時空與強度進行轉(zhuǎn)錄表達,進而抵抗不利環(huán)境[11]。目前,已經(jīng)鑒定的植物抗逆轉(zhuǎn)錄因子包括 MYB、WRKY、ZFP、AP2/ERF、bHLH、NAC、VOZ、CAMTA及 EIN3等[12-13]。其中,MYB基因家族及MYB結(jié)構(gòu)域基因進化時間較長,且存在于所有真核生物中,是植物界最大的轉(zhuǎn)錄因子家族之一[14],參與植物細胞分化、葉片等器官的形態(tài)建成、次生物質(zhì)代謝、細胞周期調(diào)節(jié)及環(huán)境因子的應答等過程[15]。根據(jù)MYB結(jié)構(gòu)域的數(shù)量及分布,MYB基因一般分為4種類型,其中,R2R3-MYB是植物中分布最為廣泛的一種MYB基因[16-17],而R1R2R3-MYB(3R-MYB)主要存在于動物體中,參與動物細胞周期的調(diào)節(jié)等過程[18-19],但在植物中數(shù)量較少,一般含有4-5個[20],4R-MYB在大部分動植物中僅含有1個,而且人們對其功能的了解甚少。本研究基于小桐子基因組數(shù)據(jù)庫,對MYB基因家族在全基因組水平進行了鑒定,并克隆到一個MYB基因家族基因JcMYB308,順式作用元件與qRT-PCR分析顯示其可能參與小桐子多種非生物脅迫如低溫的應答過程,本研究為進一步對該基因進行功能驗證及轉(zhuǎn)基因研究奠定了基礎(chǔ)。
1 材料與方法
1.1 材料
1.1.1 實驗材料及處理 本實驗選用的小桐子種子取自云南省楚雄州元謀縣。選取飽滿的小桐子種子,1.5% CuSO4溶液消毒15 min后,用無菌水漂洗3-5次,置于26℃的恒溫培養(yǎng)箱中吸漲24 h。隨后,將吸漲的種子于無菌水中漂洗2-3次后,置于墊有5層無菌水濕潤濾紙的托盤中,在相對濕度(RH)為75%、26/20℃、16/8 h光周期的恒溫培養(yǎng)箱中萌發(fā)5 d。選取發(fā)芽較好的種子播于消毒的培養(yǎng)土中,并于相同條件的恒溫培養(yǎng)箱中生長約15 d,直至第2片真葉展開。隨后將生長15 d后的小桐子幼苗置于相對濕度(RH)為75%、12℃、16/8 h光周期的低溫培養(yǎng)箱中進行低溫脅迫處理,分別取低溫脅迫0.5、3、12、24、48 h與對照(正常培養(yǎng))的第2片真葉、莖及根,置于貼好標簽的自封袋中,液氮速凍后存于-80℃冰箱中用于RNA的提取。
1.1.2 實驗的主要試劑 本研究中大腸桿菌 Trans1-T1(DH5α)、TransZol Up、DNase I、TransStart Taq DNAPolymerase、氨芐青霉素(Amp)、X-gal、IPTG、2× Easy Taq PCR SuperMix(+dye)、EasyPure Plasmid MiniPrep Kit、pEASY-T1 Cloning Kit、Trans 2K Plus II DNA Marker、TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix、TransStart Top Green qPCR SuperMix等購自北京全式金生物技術(shù)有限公司;引物合成及測序由華大基因有限公司完成。
1.2 方法
1.2.1 小桐子MYB基因家族全基因組成員鑒定及序列分析 從GenBank下載小桐子最新注釋蛋白質(zhì)數(shù)據(jù)庫,通過Pfam(http://pfam.xfam.org/)下載MYB結(jié)構(gòu)域的隱馬可夫模型(PF02701),利用Hmmer3.0軟件的Hmmsearch程序?qū)π⊥┳拥鞍踪|(zhì)數(shù)據(jù)庫進行比對(閾值E<1e-10,序列相似性>50%),得到候選的小桐子MYB蛋白質(zhì)序列,并手工去除重復序列。將非冗余的候選序列利用Pfam與CDD在線工具分析MYB蛋白結(jié)構(gòu)域做進一步篩選,得到最終的小桐子MYB家族蛋白序列。將鑒定的小桐子MYB蛋白序列利用ClustalX(Version2.0)進行序列相似性比對,然后用MEGA6.0軟件通過鄰接法(NJ)構(gòu)建系統(tǒng)進化樹,并采用自展法(Bootstrap)進行檢驗。
1.2.2 總RNA的提取及cDNA第一鏈的合成 利用兩步裂解法提取小桐子對照與12℃低溫脅迫0.5、3、12、24、48 h的根、莖及葉片的總RNA,并利用DNase I消化RNA中的殘余基因組DNA,得到純化的總RNA。分別取3 μg總RNA,以Anchored Oligo(dT)為逆轉(zhuǎn)錄引物,利用 TransScript One-Step gDNA Removal and cDNA Synthesis SuperMix合成第一鏈 cDNA。
1.2.3 小桐子JcMYB308基因cDNA片段的克隆 以1.2.2反轉(zhuǎn)錄cDNA為模板,使用雙封閉熱啟動DNA聚合酶TransStart Taq DNA Polymerase進行PCR擴增,引物序列為:JcMYB308_F1:5'-AACCCATTTGCCTTGTCT-3';JcMYB308_R1:5'-CTCCATCGCCAGTCTTCA-3'。擴增條件為:94℃預變性5 min;94℃變性30 s,51.8℃退火30 s,72℃延伸1 min,35個循環(huán);72℃后延伸20 min。切膠回收目的條帶(約700 bp),與克隆載體pEASY-T1連接后,轉(zhuǎn)化大腸桿菌感受態(tài)細胞,涂Amp抗性LB平板,過夜培養(yǎng),進行藍白斑篩選。挑取單菌落進行菌落PCR驗證陽性克隆,提取重組質(zhì)粒命名為pEASY-T1-JcMYB308。
1.2.4 小桐子JcMYB308基因的序列分析 利用BioEdit軟件將克隆的小桐子JcMYB308基因cDNA序列翻譯成氨基酸序列,用在線工具ProtParam計算其理論分子量、等電點等基本參數(shù)。將獲得的小桐子JcMYB308基因?qū)enBank小桐子基因組序列(Annotation release 101)進行tblastn檢索得到其基因與CDS序列,同時下載其它植物的MYB308基因序列、CDS序列及氨基酸序列。利用在線軟件GSDS(Gene Structure Display Server,http://gsds.cbi.pku.edu.cn/)進行CDS序列與基因序列比對以確定基因內(nèi)含子與外顯子的結(jié)構(gòu)并繪制基因結(jié)構(gòu)圖。將小桐子JcMYB308蛋白序列與其它植物MYB308進行序列相似性比對,然后用MEGA6.0軟件通過鄰接法(NJ)構(gòu)建系統(tǒng)進化樹。利用GenBank的CDD工具對比對文件進行結(jié)構(gòu)域分析并利用GenDOC軟件繪圖。
1.2.5 小桐子JcMYB308基因的熒光定量表達分析 利用1.2.2中獲得的小桐子各器官低溫脅迫條件下的cDNA為模板,以18S rRNA為內(nèi) 參(GenBank登 錄 號:AY823528), 利 用TransStart Top Green qPCR SuperMix進 行 小 桐子JcMYB308基因的qRT-PCR擴增,使用儀器為 Roche Lightcycler 96,20 μL反 應 體 系, 每 個樣品重復3次。JcMYB308基因擴增引物序列JcMYB308_F2(5'-AACCCATTTGCCTTGT-3'),JcMYB308_R2(5'-CAGCAGCCTTCACCAT-3');內(nèi)參基因18S rRNA擴增引物序列18S rRNA_F(5'-AGAAACGGCTACCACATC-3'),18S rRNA_R(5'-CCAAGGTCCAACTACGAG-3')。 擴 增 條 件 為 :94℃預變性30 s;94℃變性10 s,50.8℃退火15 s,72℃延伸15 s,45個循環(huán),之后增加溶解曲線程序:95℃ 10 s,65℃ 60 s,97℃ 1 s,連續(xù)檢測信號。采用2-ΔΔCt方法進行基因差異表達分析。組織差異表達分析以葉片的表達量為基準,低溫差異表達分析以對照(CK)的表達量為基準。
2 結(jié)果
2.1 小桐子MYB基因家族全基因組成員鑒定及序列分析
自GenBank下載小桐子最新注釋蛋白質(zhì)數(shù)據(jù)庫,結(jié)合Pfam與CDD結(jié)構(gòu)域分析共鑒定到213個小桐子MYB基因家族成員,其中1R-MYB 86個、R2R3-MYB 122個、3R-MYB 4個、4R-MYB 1個,JcMYB-308基因?qū)儆赗2R3-MYB亞家族中的一員。同時,對小桐子MYB家族進行理化性質(zhì)分析表明:小桐子213個MYB轉(zhuǎn)錄因子蛋白質(zhì)序列長度分布在88-2 039 aa之間,等電點分布在4.49-10.33之間。聚類分析顯示小桐子MYB家族分為6個亞家族(圖1)。
2.2 小桐子JcMYB308基因片段cDNA的克隆
以小桐子低溫脅迫48 h的葉片提取總RNA并反轉(zhuǎn)錄的cDNA為模板擴增JcMYB308基因片段。結(jié)果表明,擴增產(chǎn)物約713 bp,與預期設計引物結(jié)果一致(圖2-A)。將目的條帶切膠回收后與T/A克隆載體pEASY-T1連接、轉(zhuǎn)化大腸桿菌,經(jīng)菌落PCR 鑒定陽性克隆(圖2-B),之后陽性克隆過夜培養(yǎng),提取重組質(zhì)粒(圖2-C)。
2.3 小桐子JcMYB308基因的序列分析
利用BioEdit 軟件將克隆的小桐子 JcMYB308 基因的部分 cDNA序列翻譯成氨基酸序列(圖 3)。

圖1 通過ClustalX序列比對MEGA6.0鄰接法構(gòu)建的小桐子MYB基因家族進化樹
從GenBank下載其它植物的MYB308蛋白序列,通過ClustalX2.0序列比對,并利用MEGA5.0構(gòu)建系統(tǒng)進化樹顯示,MYB308蛋白在N端序列保守性較高,分別鑒定到2個MYB結(jié)構(gòu)域,而中部與C端的序列一致性相對較小(圖4)。單子葉植物和雙子葉植物單獨聚類為兩個分支,而小桐子與同屬大戟科的蓖麻親緣關(guān)系最近,序列一致性為62.7%(圖5)。
為了分析小桐子JcMYB308基因的結(jié)構(gòu)特征,構(gòu)建了JcMYB308與其它物種MYB308基因內(nèi)含子-外顯子結(jié)構(gòu)圖和系統(tǒng)發(fā)育樹,結(jié)果表明,所有考察的植物MYB308基因都含有2個外顯子,且都包含5'和3'非編碼區(qū)(5'-UTR和3'-UTR)。其中,小桐子JcMYB308基因5'-UTR區(qū)域長度約為1 791 bp,與其它植物MYB308基因結(jié)構(gòu)差異較為顯著(圖6)。
2.4 小桐子JcMYB308基因的表達分析
據(jù)文獻報道,MYB類轉(zhuǎn)錄因子中包含與植物抗逆性相關(guān)的基因[21]。通過實時熒光定量PCR對小桐子JcMYB308基因進行組織差異與低溫脅迫表達分析。結(jié)果表明,JcMYB308基因在根、莖、葉器官中均有表達,但存在組織差異性。其中,在小桐子根中表達量較高;其次是莖,而在葉片中表達量相對較低(圖7-A)。另外,小桐子JcMYB308基因在葉片、莖及根中都屬于低溫誘導表達基因,在葉中,在低溫脅迫的0.5 h-12 h持續(xù)誘導高表達,并在低溫脅迫12 h達到最大表達量,較對照提高6.73倍(圖7-B),而在莖與根中,均在低溫脅迫24 h達到最大表達量,分別較對照提高18.17倍與10.93倍(圖7-C、D),說明JcMYB308基因與小桐子的抗冷性形成直接相關(guān)。
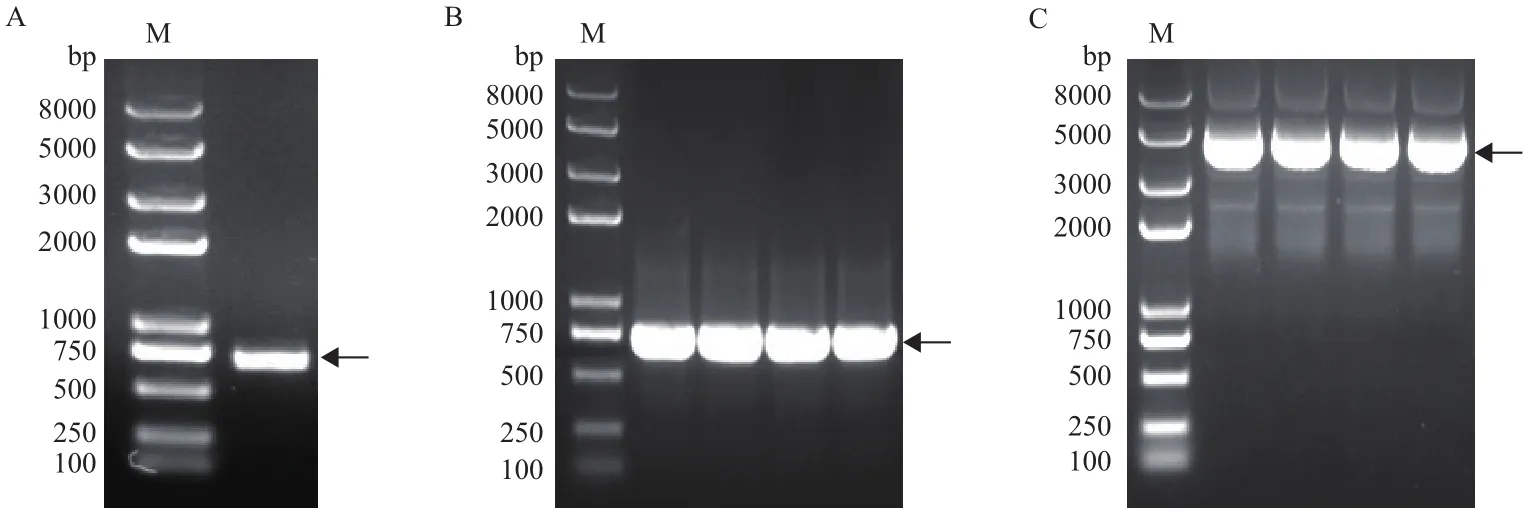
圖2 小桐子JcMYB308基因cDNA片段克隆

圖3 小桐子JcMYB308基因的部分cDNA及對應的氨基酸序列
3 討論
MYB類轉(zhuǎn)錄因子家族特有的MYB結(jié)構(gòu)域是一段含有約51-53個氨基酸殘基的肽段,包含一系列高度保守的氨基酸殘基和間隔序列[22]。其廣泛分布于真核生物中,也是植物中最大的轉(zhuǎn)錄因子家族之一[23]。生物信息學分析發(fā)現(xiàn),小桐子MYB基因家族包含213個成員,其中R2R3-MYB成員122個。目前Katiyar 等[24]報道水稻和擬南芥中分別包含155和197個MYB成員,其中發(fā)現(xiàn)水稻中含R2R3-MYB基因109個;Du等[25-27]在玉米和大豆中分別鑒定到157和244個R2R3-MYB成員;Willcins等[28]在楊樹中鑒定到192個R2R3-MYB成員;Li等[22]在黃瓜基因組中鑒定到55個R2R3-MYB基因。而本研究構(gòu)建進化樹分析顯示,胡楊、黃瓜與小桐子同屬雙子葉分支,表明它們的親緣關(guān)系、生物學功能比較近,但數(shù)據(jù)剖析結(jié)果與之相反,數(shù)量相差較大,反而與單子葉植物相接近。但一些研究者也發(fā)現(xiàn),有些雙子葉植物與小桐子R2R3-MYB成員數(shù)量相差甚少,如Matus等[29]在葡萄中鑒定到108個R2R3-MYB成員;Katiyar[24]等于擬南芥中發(fā)現(xiàn)其有126個R2R3-MYB成員。因此,可以推知,MYB基因家族成員雖然分布在不同物種中,隨著親緣關(guān)系的近與遠其數(shù)量有一定的變化,但對于大多數(shù)單、雙子葉植物來說,它們的親緣關(guān)系出現(xiàn)交叉性。
MYB基因家族不僅參與植物的大部分生長發(fā)育過程[31],還在植物體內(nèi)參與調(diào)控其生物的合成及信號傳導的過程,如調(diào)節(jié)花青素、類黃酮、木質(zhì)素的合成、毛狀體的分化、抗逆信號的傳導等[16-17]。在研究中我們發(fā)現(xiàn),小桐子JcMYB308基因?qū)儆赗2R3-MYB亞家族中的一員。諸多文獻報道,R2R3-MYB亞家族所含轉(zhuǎn)錄因子與植物的多種非生物脅迫應答如低溫密切相關(guān)。如Cominelli等和Seo等[32-33]研究發(fā)現(xiàn)在擬南芥中AtMYB60和AtMYB96在ABA信號傳遞過程中起調(diào)控作用,具有調(diào)控氣孔導度、提
高植物抗旱和抗病能力的功能;其中AtMYB30、AtMYB60、AtMYB96及VvMYB60的共同特點是都參與抗逆信號的傳遞與調(diào)控過程;Ding等[34]報道了擬南芥基因AtMYB15的過量表達可以提高擬南芥對ABA的敏感性,并提高了植物的抗旱性。本研究通過基因結(jié)構(gòu)和進化分析顯示及qRT-PCR分析結(jié)果初步斷定JcMYB308基因可能參與小桐子抗冷性的形成過程,它在不同組織中均有表達,其中于葉中,在低溫脅迫0.5 h-12 h持續(xù)誘導高表達;還發(fā)現(xiàn)它的表達于某一時段或時刻高表達后又開始趨于下降,若作圖呈現(xiàn)鐘罩性。對此現(xiàn)象我們認為,小桐子生活于干熱河谷地區(qū),由于缺乏對低溫的適應能力,對其植株于12℃進行低溫處理,達到最大抗性的低溫訓化時間后其體內(nèi)的生理生化活動依次減弱,所以表達量下降。此外,Agarwal等[21]在擬南芥中也發(fā)現(xiàn)AtMYB15參與抗寒基因的表達調(diào)控;Van等[35]研究表明與之聚類于同一亞家族的AtMYB72與植物誘導系統(tǒng)性抗性獲得有關(guān)。雖然本實驗研究的小桐子R2R3-MYB亞家族成員之一MYB308基因在其它物種中報道甚少,但基于以上研究充分證實,MYB
類轉(zhuǎn)錄因子家族與植物的耐脅迫性密切相關(guān)。就當前的研究而言,MYB基因家族雖作為一個大家族,數(shù)量甚多,但與植物抗冷性相關(guān)的基因僅有一小部分。本研究結(jié)果有助于更好地了解MYB轉(zhuǎn)錄因子在小桐子中對低溫響應的作用。

圖4 植物MYB308氨基酸多序列比對
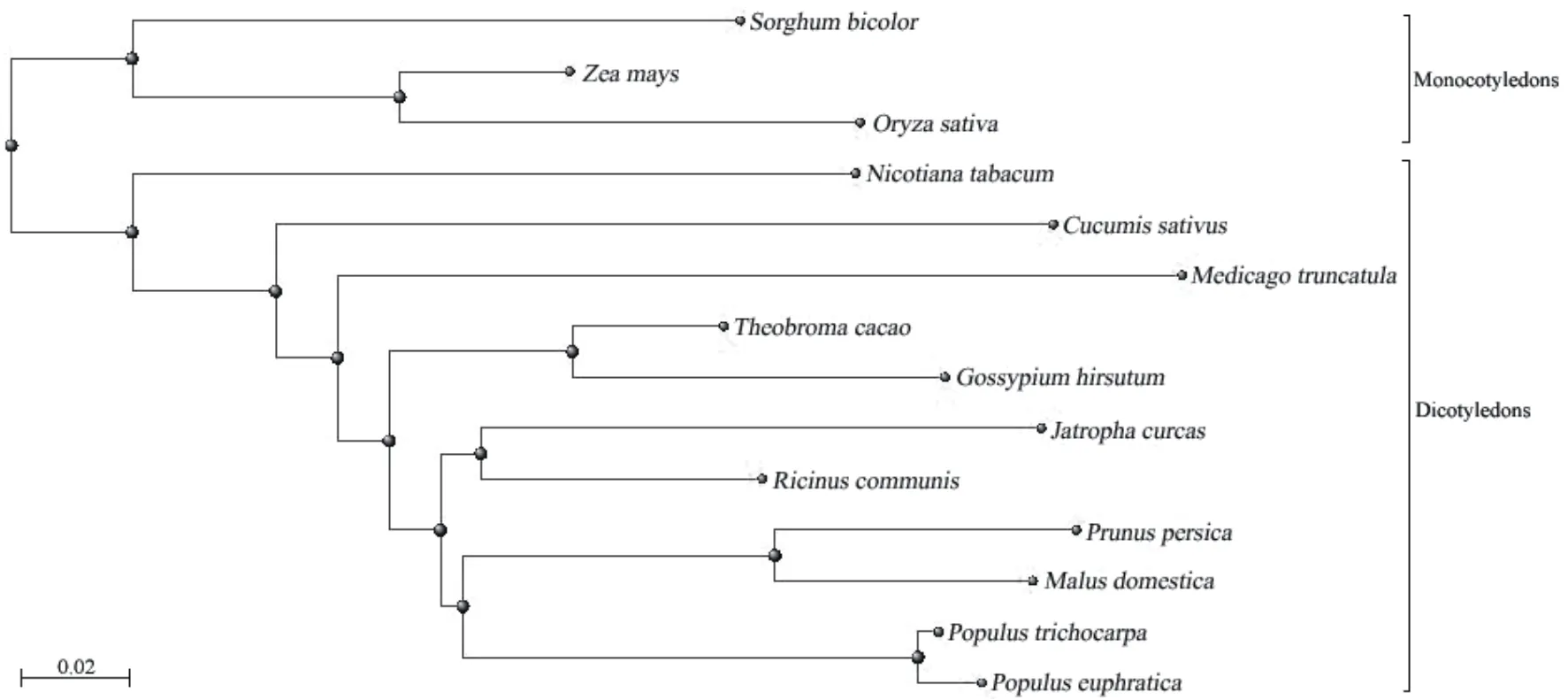
圖5 小桐子JcMYB308與其他物種MYB308轉(zhuǎn)錄因子的進化分析
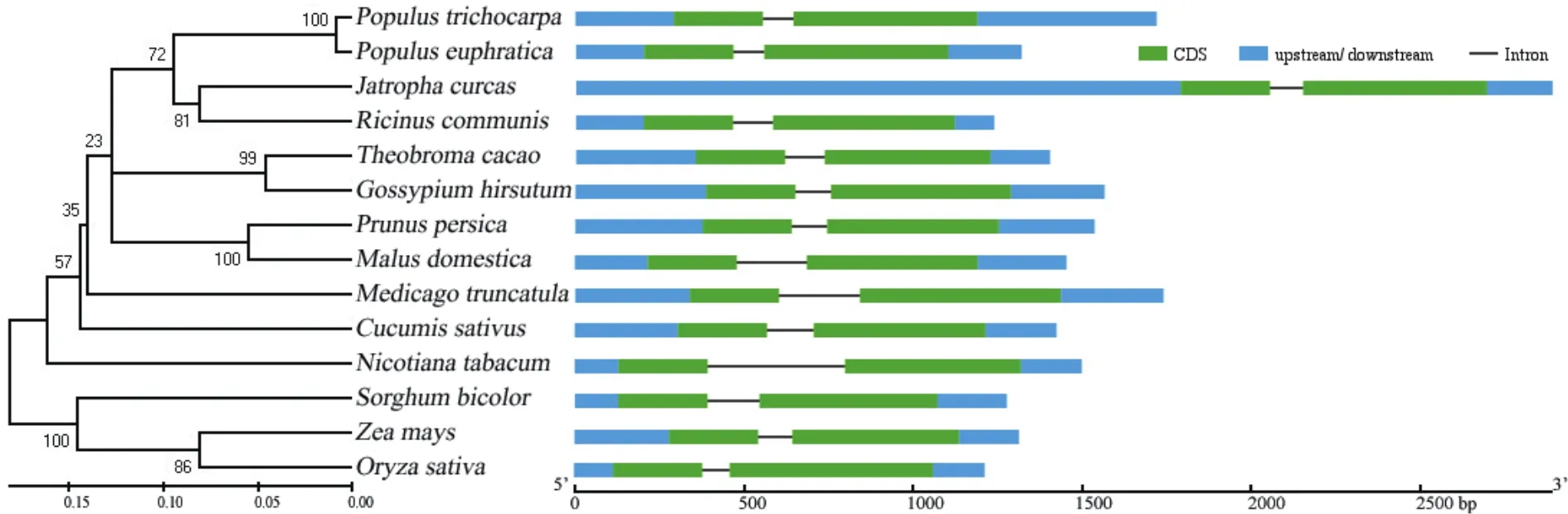
圖6 植物MYB308的基因結(jié)構(gòu)分析
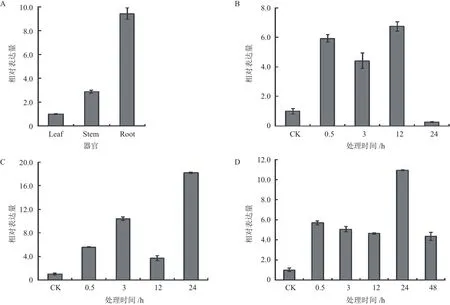
圖7 小桐子JcMYB308基因的表達模式分析
4 結(jié)論
本研究基于小桐子最新注釋全基因組數(shù)據(jù)庫,通過生物信息學鑒定得知小桐子全基因組共含213個MYB基因家族成員,聚類為6個亞家族,其中MYB308基因?qū)儆赗2R3-MYB亞家族成員。利用RTPCR成功克隆了小桐子JcMYB308基因,其克隆片段長度為713 bp,基因結(jié)構(gòu)具有較高的保守性,均含有兩個外顯子,進化樹顯示其與同屬大戟科的蓖麻親緣關(guān)系最近,序列一致性為62.7%。qRT-PCR表達分析表明,小桐子JcMYB308基因的表達存在組織表達特異性,在根中表達量較高,而在葉片中表達量相對較低,在根與莖中低溫脅迫24 h時達到最大表達量。
[1]張無敵, 宋洪川, 偉小巋, 等. 元謀縣小桐子種植的適應性研究[J]. 農(nóng)業(yè)與技術(shù) , 2001, 21(2):22-25.
[2]曾覺民. 可大力發(fā)展的生物質(zhì)能源植物-膏桐[J]. 云南林業(yè),2006, 27(2):21-22.
[3]陳麗, 吳軍, 曾妮, 等. 用GC-MS分析不同采收和貯存時期的麻瘋種子油的脂肪酸[J]. 熱帶亞熱帶植物學報, 2007, 15(5):443-446.
[4] 王濤. 中國主要生物質(zhì)燃料油木本能源植物資源概況與展望[J]. 科技導報 , 2005, 23(5):12-14.
[5]Nakashima K, Ito Y, Yamaguchi-Shinozaki K. Transcriptional regulatory network in response to abiotic stress in Arabidopsis and grasses[J]. Plant Physiology, 2009, 149(1):88-95.
[6]李靜, 吳芬宏, 陳延燕, 等. 麻風樹種子提取物對幾種害蟲的殺蟲活性[J]. 農(nóng)藥 , 2006, 45(1):57-58.
[7]范菊娣, 楊松, 宋寶安, 等. 麻風樹農(nóng)藥和醫(yī)藥生物活性研究進展[J]. 農(nóng)藥 , 2006, 45(5):298-301.
[8]王兆玉, 杜延琪, 岑岳柱, 等. 小油桐葉甲醇提取物對福壽螺的藥效試驗[J]. 南方醫(yī)科大學學報, 2009, 29(6):1235-1237.
[9]劉杰, 李黔柱, 尹航, 等. 麻風樹植物資源的研究與開發(fā)利用進展[J]. 貴州大學學報:自然科學版, 2006, 23(2):105-110.
[10] 劉玉君, 沈世華. 小桐子種子油體蛋白的提取及其電泳分析[J]. 林業(yè)科學 , 2008, 44(8):37-40.
[11]劉輝, 李德軍, 鄧治. 植物應答低溫脅迫的轉(zhuǎn)錄調(diào)控網(wǎng)絡研究進展[J]. 中國農(nóng)業(yè)科學, 2014, 47(18):3523-3533.
[12]Wang HB, Zou ZR, Wang SS, et al. Global analysis of transcriptome responses and gene expression profiles to cold stress of Jatropha curcas L[J]. PLoS One, 2013, 12(12):e82817.
[13]Wang HB, Zou ZR, et al. Deep sequencing- based transcriptome analysis of the oil-bearing plant Physic Nut(Jatropha curcas L.)under cold stress[J]. Plant Omics, 2014, 7(3):178-187.
[14]Zhang L, Ma H. Complex evolutionary history and diverse domain organization of SET proteins suggest divergent regulatory interactions[J]. New Phytologist, 2011, 195(1):248-263.
[15]Lipsick JS. One billion years of Myb[J]. Oncogene, 1996, 13(2):223-235.
[16]Jiang C, Gu J, Chopra S, et al. Ordered origin of the typical twoand three-repeat Myb genes[J]. Gene, 2004, 326(2):13-22.
[17]Wilkins O, Nahal H, Foong J, et al. Expansion and diversification of the populus R2R3-MYB family of transcription factors[J]. Plant Physiology, 2009, 149(2):981-993.
[18]Takahashi T, Nakagoshi H, Sarai A, et al. Human A-myb gene encodes a transcriptional activator containing the negative regulatory domains[J]. FEBS Letters, 1995, 358(1):89-96.
[19]Foos G, Grimm S, Klempnauer KH. Functional antagonism between members of the myb family:B-myb inhibits v-myb-induced gene activation[J]. EMBO J, 1992, 11(12):4619-4629.
[20]Kranz H, Scholz K, Weisshaar B. c-MYB oncogene-like genes encoding three MYB repeats occur in all major plant lineages[J].Plant Journal, 2000, 21(2):231-235.
[21]Agarwal M, Hao YJ, Kapoor A, et al. A R2R3 Type MYB transcription factor Is involved in the cold regulation of CBF genes and in acquired freezing tolerance[J]. J Biol Chem, 2006, 281(49):37636-37645.
[22]Li Q, Zhang C, Li J, et al. Genome-wide identification and characterization of R2R3-MYB family in Cucumis sativus[J].PLoS One, 2012, 7(10):e47576.
[23]Riechmann JL, Heard J, Martin G, et al. Genome-wide comparative analysis among eukaryotes[J]. Science, 2000, 290(5499):2105-2110.
[24]Katiyar A, Smita S, Lenka SK, et al. Genome-wide classification and expression analysis of MYB transcription factor families in rice and Arabidopsis[J]. BMC Genomics, 2012, 13 :544.
[25]Du H, Feng BR, Yang SS, et al. The R2R3-MYB transcription factor gene family in maize[J]. PLoS One, 2012, 7(6):e37463.
[26]Du H, Yang SS, Liang Z, et al. Genome-wide analysis of the MYB transcription factor superfamily in soybean[J]. BMC Plant Biology, 2012, 12(7):106.
[27] Du H, Feng BR, et al. The R2R3-MYB transcription factor gene family in maize[J]. PLoS One, 2012, 7(6):e37463.
[28]Willcins O, Nahal H, Foong J, et al. Expansion and diversification of the Populus R2R3-MYB family of transcription factors[J].Plant Physiology, 2009, 149(2):981-993.
[29]Matus JT, Aquea R, Arce-Johnson P. Analysis of the grape MYB R2R3 subfamily reveals expanded wine quality-related glades and conserved gene structure organization across Vitis and Arabidopsis genomes[J]. BMC Plant Biology, 2008, 8(7):83.
[30]Braun EL, Grotewold E. Newly discovered plant c-myb-like genes rewrite the rvolution of the plant MYB gene family[J]. Plant Physiology, 1999, 121(1):21-24.
[31]段俊枝, 李瑩, 周雷, 等. 利用基因工程技術(shù)提高水稻耐冷性的研究進展[J]. 浙江農(nóng)業(yè)學報, 2015, 27(4):705-712.
[32]Cominelli E, Galbiati M, Vavasseur A, et al. A guard-cell-specific MYB transcription factor regulates stomatal movements and plant drought tolerance[J]. Curr Biol, 2005, 15(13):1196-1200.
[33]Seo PJ, Park CM. MYB96-mediated abscisic acid signals induce pathogen resistance response by promoting salicylic acid biosynthesis in Arabidopsis[J]. New Phy, 2010, 186(2):471-483.
[34]Ding ZH, Li SM, An XL, et al. Transgenic expression of MYB15 confers enhanced sensitivity to abscisic acid and improved drought tolerance in Arabidopsis thaliana[J]. Genetics and Genomics,2009, 36(1):17-29.
[35]Van dES, Verhagen BW, Van DR, et al. MYB72 Is required in early signaling steps of rhizobacteria-induced systemic resistance in Arabidopsis[J]. Plant Physiology, 2008, 146(3):1293-1304.
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領(lǐng)巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
