白背飛虱幾丁質合成酶1基因的結構及特性研究
2018-01-31 03:45:35陳靜張道偉錢正敏
生物技術通報 2018年1期
關鍵詞:檢測
陳靜 張道偉 錢正敏
(1. 遵義醫學院基礎醫學院,遵義 563000;2. 遵義師范學院生命科學學院,遵義 563002)
幾丁質是一種在真菌、甲殼動物及節肢動物中廣泛存在的多糖類生物高分子,是昆蟲氣管、表皮、圍食膜等組織的重要組成部分,在對昆蟲內部組織器官的保護和抵抗外源生物的侵染中發揮著非常重要的作用[1-3]。幾丁質的合成和降解是昆蟲生長發育的重要過程,所以也成為害蟲控制的理想靶標。幾丁質對昆蟲具有保護作用,但同時也限制了昆蟲的生長,所以當昆蟲成長到一定階段時,就需要蛻去舊表皮,合成更大的新表皮,以適應昆蟲的生長。而新表皮的合成就是由幾丁質合成酶(Chitin synthase,CHS)等酶系催化合成新的幾丁質到新的表皮中。
CHS是幾丁質合成的關鍵酶之一。理論分子量為160-180 kD,是糖基轉移酶家族成員[4-5]。通過對不同種昆蟲的CHS蛋白序列進行比對,結果發現CHS有3個相對保守的結構域:結構域A、B、C。結構域A位于CHS的N端,是一段跨膜螺旋結構;結構域B位于CHS的活性中心,含有2個保守的氨基酸組成EDR和QRRRW;結構域C位于CHS的C端,包含一段相對保守的氨基酸序列和7個跨膜螺旋,這個區域被認為參與對幾丁質合成的調控。在大多數昆蟲中存在兩個CHS:CHS1和CHS2,這兩個CHS被認為參與不同組織器官的幾丁質合成。其中,CHS1主要參與表皮和氣管中幾丁質的合成,CHS2主要參與圍食膜中幾丁質的合成。另外,有研究發現在岡比亞按蚊Anopheles gambiae的復眼中可以檢測到CHS1和CHS2基因的表達。赤擬谷盜中可以檢測到CHS1基因主要在卵和蛹期中表達[6-7]。對不同種昆蟲中CHS基因的結構研究發現,大多數昆蟲的CHS1基因存在一個可變剪接的外顯子,而CHS2基因中沒有發現可變剪接的現象。CHS基因的序列在多種昆蟲中已被陸續獲得,如銅綠蠅(Lucilia cuprina)、埃及伊蚊(Aedes aegypti)、果蠅(Drosophila melanogaster)、赤擬谷盜(Tribolium castaneum)、甜菜夜蛾(Spodoptera exigua)、岡比亞按蚊(A.gambiae)等,但是還有待于更深層次的研究[8]。RNA干擾(RNAi,RNA interference)技術已被廣泛應用于基因功能研究中。利用RNAi技術研究CHS1基因對東亞飛蝗(Locusta migratoria manilensis)生長發育的影響,結果發現沉默CHS1的可變外顯子a會導致東亞飛蝗發育遲緩,蛻皮障礙,而沉默CHS1的可變外顯子b會引起腿部畸形[9]。
白背飛虱隸屬同翅目飛虱科,是危害我國水稻正常生長的主要害蟲之一[10]。白背飛虱除了自身刺吸水稻汁液外,還會傳播水稻病毒,給水稻的生產帶來毀滅性的災難[11-12]。本研究基于白背飛虱的轉錄組數據獲得了白背飛虱CHS1基因的全長序列,對該基因序列的兩個可變外顯子進行了分析并探究了白背飛虱中CHS1基因在各齡期發育階段及各組織器官中的表達水平。通過顯微注射的方法對白背飛虱的CHS1基因進行了RNAi實驗。通過上述研究,旨為闡明CHS1基因在白背飛虱中的生理作用,為利用CHS1基因治理白背飛虱奠定基礎。
1 材料與方法
1.1 材料
1.1.1 實驗材料 白背飛虱由本實驗室飼養于人工氣候室無任何處理的水稻上。飼養條件:(27±1)℃,相對濕度70%,光周期16L∶8D。
1.1.2 試劑 PCR相關試劑,反轉錄RT-PCR相關試劑,熒光定量PCR相關試劑,克隆載體pMD-18T,DNA分子量標準Marker均購自TAKARA公司;動物RNA提取試劑盒,膠回收試劑盒、質粒回收試劑盒購自北京天根公司;大腸桿菌感受態細胞DH5α購自北京全式金公司;dsRNA合成試劑盒T7 RiboMax Express RNAi System購自Promega公司;引物合成及序列測定由上海生工公司完成,其它常規試劑均為國產分析純。
1.2 方法
1.2.1 白背飛虱CHS1基因的獲得 通過對本課題組前期獲得的白背飛虱轉錄組數據的分析,得到白背飛虱CHS1基因。根據這段序列設計全長擴增的驗證引物SfCHS1-F和SfCHS1-R(引物序列見表1)。PCR擴增程序:94℃預變性5 min,94℃變性30 s,56℃退火30 s,72℃延伸5 min,35個循環,72℃延伸10 min。擴增產物經凝膠回收后與pMD-18T載體連接,轉化感受態細胞DH5α,菌落PCR鑒定正確后進行測序。
1.2.2 白背飛虱CHS1基因可變剪接的驗證 根據已有研究報道,昆蟲CHS1基因存在可變剪接現象,所以我們對比了不同昆蟲CHS1基因可變剪接出現的位置,在可變剪接位點附近設計了引物SfCHS1-YZF和SfCHS1-YZR(表1)進行驗證。PCR擴增程序:94℃預變性5 min;94℃變性30 s,56℃退火30 s,72℃延伸 1 min,35個循環;72℃延伸 10 min。擴增產物經凝膠回收后與pMD-18T載體連接,轉化感受態細胞DH5α,菌落PCR鑒定正確后進行測序。

表1 本實驗所用的引物
1.2.3 白背飛虱CHS1基因的時空表達特異性檢測 取4齡1-3 d,5齡1-3 d的白背飛虱若蟲,羽化1 d和2 d的白背飛虱成蟲,分別用RNA提取試劑盒進行RNA的提取及反轉錄。采用實時熒光定量PCR技術檢測SfCHS1基因在白背飛虱4齡到成蟲各蟲期的表達情況。
取5齡1 d的白背飛虱若蟲,CO2麻醉后于冰上解剖出表皮、氣管和腸。分別用RNA提取試劑盒進行RNA的提取及反轉錄。采用實時熒光定量PCR技術檢測SfCHS1基因在白背飛虱表皮,氣管,腸中表達情況。用于檢測SfCHS1基因時空表達特異性的引物為:SfCHS1-YGF和SfCHS1-YGR(表1)。以白背飛虱β-actin基因為內參進行熒光定量PCR,檢測引物為β-actin-YGF和β-actin-YGR。熒光定量PCR反應體系:SYBR(2×)5 μL,cDNA 模板 1 μL,上、下游引物各 0.2 μL,ddH2O 3.6 μL。反應條件為 :95℃預變性10 s,95℃變性5 s,60℃延伸30 s,35
個循環。每個樣品重復3次測定。
1.2.4 顯微注射RNAi實驗 設計用于合成SfCHS1基因dsRNA的引物SfCHS1-dsRNA-F和SfCHS1-dsRNA-R及陰性對照綠色熒光蛋白GFP基因的引物 GFP-dsRNA-F 和 GFP-dsRNA-R( 表 1), 按 照dsRNA合成試劑盒的說明進行SfCHS1基因及GFP基因dsRNA的合成,用微量注射儀(Eppendorf公司)將dsRNA導入白背飛虱5齡1 d的若蟲體內(10 ng/頭,注射方法參照文獻[12]),以無關dsRNA-GFP作為對照,采用實時熒光定量PCR方法檢測SfCHS1基因的相對表達量。
1.2.5 序列及數據分析 引物設計采用Primer 5軟件,測序結果用DNAMAN軟件進行拼接,SfCHS1基因開放閱讀框分析采用DNAstar軟件。氨基酸序列同源比對采用BLASTx程序,蛋白系統進化樹構建采用MEGA 7.0 軟件,以Neighbor-joining算法進行分析,蛋白分子量及等電點預測采用在線軟件(http://www.expasy.org),糖基化位點預測采用在線軟件(http://www.cbs.dtu.dk/services/NetNGlyc/), 數 據的統計分析采用SPSS 19.0軟件。
2 結果
2.1 白背飛虱CHS1基因的序列驗證及分析
通過對轉錄組測序數據的分析及全長擴增驗證,獲得了白背飛虱SfCHS1基因全長序列(GenBank登陸號:KY987034.1)。SfCHS1基因開放閱讀框ORF長4 719 bp,編碼1 572個氨基酸。預測蛋白的分子量為180.6 kD,等電點為6.72。預測有6個N-糖基 化 位 點(18-21:NFSD;425-428:NLTI;917-920:NISL;956-959:NWTS;1 289-1 292:NSSV;1 327-1 330:NISY),有16個跨膜螺旋結構。根據已有研究報道,昆蟲CHS1基因存在可變剪接現象,所以對比了不同昆蟲CHS1基因可變剪接出現的位置,在可變剪接位點附近設計了引物,然后通過PCR擴增和測序,獲得了白背飛虱幾丁質合成酶1的兩個可變外顯子SfCHS1a和SfCHS1b,兩個外顯子區共有54個核苷酸和13個氨基酸的差異(圖1)。可變外顯子包含一個跨膜區段。
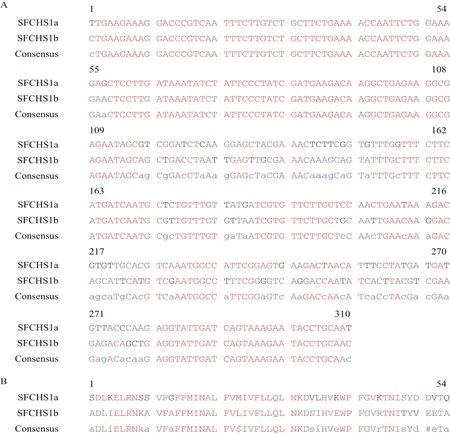
圖1 白背飛虱SfCHS1基因兩個可變外顯子的核苷酸和蛋白序列比對結果
2.2 白背飛虱CHS1基因的進化樹分析
利用Mega 7軟件將白背飛虱CHS1基因的蛋白序列與其它昆蟲的CHS1基因的蛋白序列進行進化樹分析,結果(圖2)發現白背飛虱的CHS1基因與同為同翅目飛虱科的灰飛虱(L striatellus)及褐飛虱(N. lugens)的親緣關系最近,達到97%。這個結果與Vector NTI軟件分析出的白背飛虱CHS1蛋白序列與其他昆蟲的CHS1蛋白序列比對的結果一致。
2.3 白背飛虱CHS1基因的時間表達特性
選取4齡1-3 d,5齡1-3 d,成蟲1-2 d共8個時間點對SfCHS1基因的表達進行了分析,結果(圖3)發現SfCHS1基因在4齡和5齡期的第1天,也即蛻皮完成后,合成新的表皮時表達量最高,在表皮合成后的幾天快速下降,在成蟲期的第1天也會上升,而后逐漸下降。在4齡和5齡期的最后1 d,SfCHS1基因的表達量最低。
2.4 白背飛虱CHS1基因的組織表達特異性
選取5齡1 d的白背飛虱若蟲解剖后進行組織特異性表達檢測。定量PCR檢測結果(圖4)表明,白背飛虱的CHS1基因主要在表皮中大量表達,相對表達量為25,其次為氣管,相對表達量為7.8,在腸中也有少量表達,相對表達量為2.5。

圖2 CHS1蛋白的進化樹分析
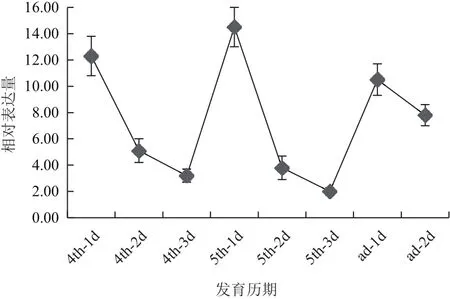
圖3 定量PCR檢測SfCHS1 mRNA發育表達模式
2.5 顯微注射RNAi后CHS1基因的表達分析
選取5齡1 d的白背飛虱若蟲,用顯微注射儀將SfCHS1基因的dsRNA(10 ng/頭)導入白背飛虱體內,在注射后的24 h和 48 h分別檢測SfCHS1基因的表達量。結果如圖5所示,在注射SfCHS1基因dsRNA后的24 h,即可檢測到SfCHS1基因的表達量出現顯著下降,為對照組表達量的40%,注射48 h后,SfCHS1基因的表達量僅為對照的9%,均存在極顯著差異(P<0.01,t-test)。
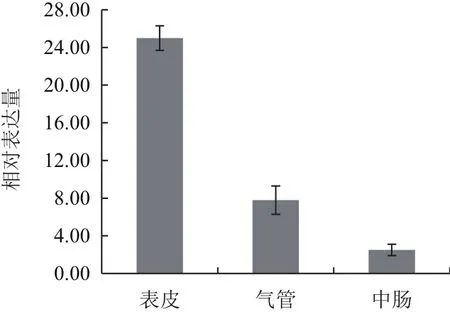
圖4 SfCHS1基因在不同組織中的相對表達量
2.6 顯微注射RNAi后白背飛虱的存活率
選取5齡1 d的白背飛虱若蟲,用顯微注射儀將SfCHS1基因及GFP基因的dsRNA(10 ng/頭)導入白背飛虱體內(每組20頭蟲,3組重復),在注射后的24 h、48 h、72 h、96 h分別統計白背飛虱的存活率。結果如圖6所示,CHS1基因干擾后對白背飛虱的存活率有極大的影響,注射48 h后,存活率與對照相比,有極顯著性差異(P<0.01,t-test),為對照的0.6倍,注射72 h后,存活率為對照的0.3倍,注射96 h后,存活率為對照的0.13倍。
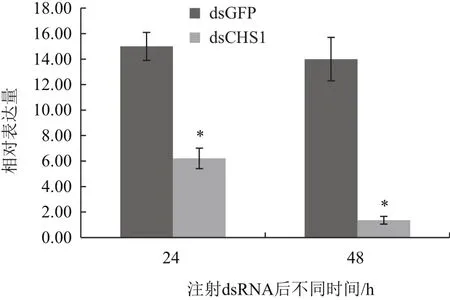
圖5 RNAi后SfCHS1基因的相對表達量
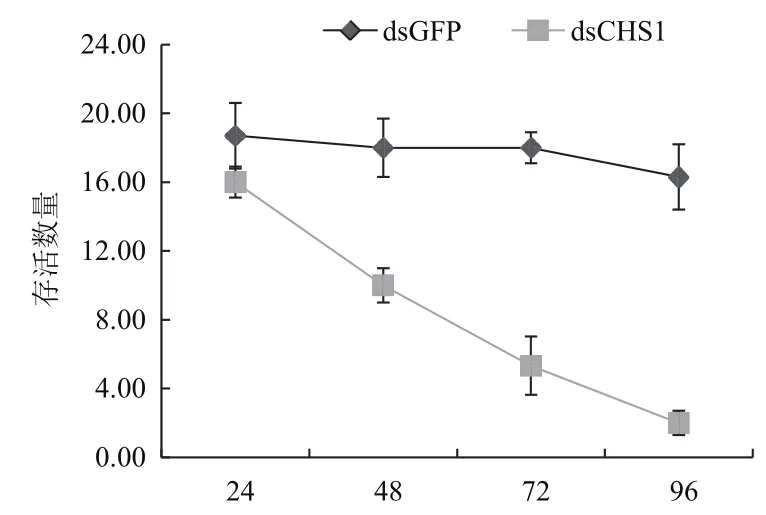
圖6 RNAi后白背飛虱的存活率
3 討論
昆蟲在生長發育的過程中要經歷舊表皮的蛻去和新表皮的生長,而幾丁質作為昆蟲表皮的主要成分,其合成和分解也是被嚴格調控的。在幾丁質合成的過程中,CHS扮演著非常重要的作用,是幾丁質合成的關鍵酶。通過本課題組前期的白背飛虱轉錄組數據分析,獲得了白背飛虱CHS1基因的全長序列。目前,研究發現在很多昆蟲中CHS1基因存在可變剪接的現象,如岡比亞按蚊、赤擬谷盜、桔小實蠅、褐飛虱等[13-17]。為了探究白背飛虱中是否也存在這樣的可變剪接,對比了白背飛虱和褐飛虱的CHS1基因序列,在可能出現可變剪接的位置設計了引物進行序列的擴增和測序,結果發現在白背飛虱中也是存在兩個可變外顯子。利用進化樹分析軟件Mega 7分析白背飛虱與其它昆蟲CHS1蛋白的進化關系,結果顯示白背飛虱CHS1蛋白與同為同翅目的昆蟲親緣關系較近,聚為一小支,其中,又與飛虱科的褐飛虱及灰飛虱的關系最近。進化樹分析的結果在一定程度上反映了CHS1蛋白的進化歷程,為后續CHS1基因功能的研究奠定基礎。然后采用熒光定量PCR的方法對白背飛虱CHS1基因在8個不同的時間點的mRNA表達量進行了檢測。結果發現CHS1基因存在明顯的時間表達特異性,在白背飛虱若蟲4齡和5齡期的第1天表達量最高,隨即快速下降,在成蟲期的第1天也會上升,而后逐漸下降。這一結果表明在每個齡期初始,即需要大量幾丁質參與表皮形成時,參與幾丁質合成的CHS1基因的mRNA表達量會快速升高,而在每個齡期的最后1 d,即蛻皮開始時,表達量是最低的。同樣,采用熒光定量PCR的方法,我們檢測了CHS1基因在白背飛虱3個組織器官中mRNA的表達水平。結果發現CHS1基因在檢測的3個組織即表皮、氣管和中腸中都有表達,但是表達量存在極顯著差異。其中,在表皮中的表達量最高。應該是在表皮合成的過程中,需要大量幾丁質的原因。另外,幾丁質也是氣管的重要組成部分,所以在氣管中也檢測到了CHS1基因的表達,只是表達量顯著低于表皮中。在中腸中也檢測到了CHS1基因的少量表達,推測可能是在中腸中有含有幾丁質的組織,也有可能是解剖中腸的時候,上面粘黏了氣管組織。
目前,褐飛虱已成為我國水稻上的首要害蟲,而殺蟲劑大規模使用帶來的害蟲抗藥性和對環境的污染使無害防治理念逐漸形成。目前,利用RNAi技術防治褐飛虱的研究報道已有很多,像通過飼喂法RNAi技術研究海藻糖合成酶的功能[18],構建RNAi載體轉化水稻研究特定基因的功能等[19]。而RNAi轉基因水稻將有可能成為防治褐飛虱的潛在方法。在本實驗中,通過注射法RNAi干擾檢測SfCHS1基因在mRNA表達水平上的沉默效果。結果顯示,注射較低劑量(10 ng/頭)的雙鏈RNA即可產生較好的干擾效果,顯著降低SfCHS1基因的表達及產生明顯的死亡表型。通過本研究對SfCHS1基因的干擾研究,為探究其功能和作用機理奠定了基礎。
4 結論
本研究通過前期對白背飛虱進行了轉錄組的測序和數據分析,獲得了白背飛虱幾丁質合成酶1基因 SfCHS1的cDNA全長序列。SfCHS1基因開放閱讀框為4 719 bp,編碼1 572個氨基酸。PCR擴增及測序結果表明SfCHS1基因存在兩個可變剪切,產生兩個轉錄本(CHS1a和CHS1b)。時間表達特異性分析結果表明,SfCHS1在每個齡期的第1天即幾丁質合成時的表達量最高。組織特異性檢測結果表明,SfCHS1主要在白背飛虱的表皮中表達,其次為氣管,在中腸中也有少量表達。顯微注射RNAi的結果表明該方法能顯著降低SfCHS1基因的轉錄水平,達到良好的RNAi效果并產生顯著的死亡表型。
[1]Cohen E. Chitin synthesis and inhibition, a revisit[J]. Pest Mangement Science, 2001, 57(10):946-950.
[2]Merzendorfer H. The cellular basis of chitin synthesis in fungi and insects:common principles and differences[J]. European Journal of Cell Biology, 2011, 90(9):759.
[3]Merzendorfer H. Insect chitin synthases:a review[J]. Journal of Comparative Physiology B, 2006, 176(1):1-15.
[4]Au-Young J, Robbins PW. Isolation of a chitin synthase gene(CHS1)from Candida albicans by expression in Saccharomyces cerevisiae[J]. Molecular Microbiology, 1990, 4(2):197-207.
[5]Yarden O, Yanofsky C. Chitin synthase plays a major role in cell wall biogenesis in Neurospora crassa[J]. Genes Dev, 1991, 5:2420-2430.
[6]Arakane Y, Zhu YC, Kramer KJ, et al. Characterization of two chitin synthase genes of the red flour beetle, Tribolium castaneum, and alternate exon usage in one of the genes during development[J].Insect Biochemistry and Molecular Biology, 2004, 34:291-304.
[7]Arakane Y, Muthukrishnan S, Kramer KJ, et al. The Tribolium chitin synthase genes TcCHS1 and TcCHS2 are specialized for synthesis of epidermal cuticle and midgut peritrophic matrix[J]. Insect Molecular Biology, 2005, 14:453-463.
[8]Tellam R, Vuocolo T, Johnson SE, et al. Insect chitin synthase cDNA sequence, gene organization and expression[J]. European Journal of Biochemistry, 2000, 267:6025-6043.
[9] Zhang J, Liu X, Zhang J, et al. Silencing of two alternative splicingderived mRNA variants of chitin synthase 1 gene by RNAi is lethal to the oriental migratory locust, Locusta migratoria manilensis(Meyen)[J]. Insect Biochem Mol Biol, 2010, 40(11):824-833.
[10]沈君輝, 尚金梅, 劉光杰. 中國的白背飛虱研究概況[J]. 中國水稻科學, 2003, 17:7-22.
[11]周國輝, 溫錦君, 蔡德江等. 呼腸孤病毒科斐濟病毒屬一新種:南方水稻黑條矮縮病毒[J]. 科學通報, 2008, 53(20):2500-2508.
[12]翟保平, 周國輝, 陶小榮等. 稻飛虱暴發與南方水稻黑條矮縮病流行的宏觀規律和微觀機制[J]. 應用昆蟲學報, 2011, 48(3):480-487.
[13]Wang Y, Fan HW, Huang HJ, et al. Chitin synthase 1 gene and its two alternative splicing variants from two sap-sucking insects,Nilaparvata lugens and Laodelphax striatellus(Hemiptera:Delphacidae)[J]. Insect Biochem Mol Biol, 2012, 42(9):637-646.
[14]Li T, Chen J, Fan X, et al. MicroRNA and dsRNA targeting chitin synthase A reveal a great potential for pest management of the hemipteran insect Nilaparvata lugens[J]. Pest Management Science. 2017, 73(7):1529-1537.
[15]Yang WJ, Xu KK, Cong L, et al. Identification, mRNA expression,and functional analysis of chitin synthase 1 gene and its two alternative splicing variants in oriental fruit fly, Bactrocera dorsalis[J]. Int J Biol Sci, 2013, 9(4):331-342.
[16]Zhang X, Zhu KY. Biochemical characterization of chitin synthase activity and inhibition in the African malaria mosquito, Anopheles gambiae[J]. Insect Science, 2013, 20(2):158-166.
[17]Zhuo W, Fang Y, Kong L, et al. Chitin synthase A:a novel epidermal development regulation gene in the larvae of Bombyx mori[J]. Mol Biol Rep, 2014, 41(7):4177-4186.
[18]Chen J, Zhang DW, Yao Q, et al. Feeding-based RNA interference of a trehalose phosphate synthase gene in the brown planthopper,Nilaparvata lugens[J]. Insect Mol Biol, 2010, 19(6):777-786.
[19]Zha W, Peng X, Chen R, et al Knockdown of midgut genes by dsRNA-transgenic plant-mediated RNA interference in the hemipteran insect Nilaparvata lugens[J]. PLoS One, 2011, 6(5):e20504.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
