梨全基因組生長素反應因子(ARF)基因家族鑒定及表達分析
2018-02-04 08:56:15歐春青姜淑苓王斐趙亞楠
中國農業科學 2018年2期
關鍵詞:分析
歐春青,姜淑苓,王斐,趙亞楠
?
梨全基因組生長素反應因子(ARF)基因家族鑒定及表達分析
歐春青,姜淑苓,王斐,趙亞楠
(中國農業科學院果樹研究所/農業部園藝作物種質資源利用重點實驗室, 遼寧興城 125199)
【目的】明確梨轉錄因子在梨基因組中的數量、結構和組織表達差異,為揭示轉錄因子在梨樹生長素信號途徑及在矮生梨生長發育中的作用奠定理論基礎。【方法】根據文獻報道的蘋果、擬南芥等植物中的轉錄因子基因,利用BLAST軟件鑒定梨基因組中的。采用SMART、PROSITE、WebLogo 3、DNAMAN 5.0、MEME、GSDS 2.0和MEGA 5.1等軟件對其蛋白和基因序列進行生物信息學分析。采用qPCR方法檢測梨在矮生梨‘中矮1號’不同組織器官及3個不同生長勢梨品種木質部和韌皮部中的表達情況。【結果】鑒定得到31個梨,所有PbARF蛋白均含有1個Auxin_resp結構域和1個B3結構域,除PbARF11、12、24、25和26外,其他序列的C末端還存在一個PB1(AUX_IAA)結構域。保守元件分析表明,PbARF基因家族包含15個保守元件,并非每個蛋白均含有所有保守元件。分組鑒定和進化樹分析結果顯示,PbARF蛋白可以被分為4組。基因結構分析表明,基因家族成員由2—15個外顯子組成,基因結構進化高度保守。qPCR結果顯示,所有在‘中矮1號’根、韌皮部、木質部、葉、花和果實中均有表達,表達模式各有不同,在3個梨品種韌皮部中的表達表現出植株越矮表達量越高的趨勢,等4個基因在3個梨品種木質部中的表達表現為植株越矮表達量越低的趨勢。【結論】梨基因組中含有31個基因家族成員;所有PbARF蛋白均含有Auxin_resp和B3兩個高度保守的結構域;結構進化高度保守;所有在‘中矮1號’不同組織器官、基砧杜梨根系及3個梨品種的木質部和韌皮部中均有表達,其中,、等5個基因的表達可能與梨樹植株的高矮有關。
梨;ARF;轉錄因子;生物信息學;基因表達
0 引言
【研究意義】‘中矮1號’是中國農業科學院果樹研究所選育的中國第一個優良梨樹矮化砧木,其株型矮化緊湊,且具有使嫁接樹矮化、早果的作用[1],在梨樹矮化密植栽培中發揮著重要作用,但其矮生和矮化機制尚不清楚,限制了新的矮生型品種和矮化砧木的選育及改良。植株高矮與莖的伸長緊密相關,生長素是調節植物莖伸長的重要激素之一,而植物對生長素的反應受許多基因的調控,生長素響應因子(auxin response factor,ARF)則是其中一類較為重要的調節基因,它能夠通過調節植物對生長素的反應而調控植株的生長發育[2-3]。因此,鑒定梨基因組中轉錄因子成員并分析其結構及其在矮生梨樹不同組織器官中的表達規律,將有助于解析該家族基因在矮生梨樹中的作用機制,同時,也為進一步克隆梨樹和研究其在生長素信號途徑中的作用奠定基礎。【前人研究進展】轉錄因子調節生長素反應是通過調節生長素反應基因的表達來實現的,它可以與植物生長素反應基因啟動子上的生長素反應元件(auxin response elements,AuxRE)TGTCTC特異結合,進而調節生長素反應基因的表達[4]。在植物中,轉錄因子是以基因家族的形式存在的,是第一個被鑒定出來的[5],繼而在擬南芥中共鑒定出23個[6],隨著越來越多植物基因組測序的完成,很多大田作物、蔬菜和果樹中的也已經被鑒定出來,如玉米中鑒定出31個[7],水稻25個[8],大豆51個[9],番茄17個[10],黃瓜18個[11],葡萄19個[12],蘋果29個[13],甜橙19個[14],番木瓜11個[15],香蕉47個[16]。【本研究切入點】梨基因組測序數據的發布已使梨基因組中的分離與鑒定成為可能[17],但還未見相關研究報道,尤其缺少不同的表達與植株高矮關系的研究。【擬解決的關鍵問題】本研究利用生物信息學方法鑒定梨基因組中所有,以明確該家族基因在梨樹中的結構特點與進化特征,通過熒光定量PCR技術分析該基因家族成員在梨樹不同組織器官和不同品種中的表達差異,為進一步研究其在梨樹矮生性狀形成過程中的作用奠定基礎。
1 材料與方法
1.1 植物材料
供試材料‘中矮1號’(矮生)、‘錦香’(半矮生,‘中矮1號’母本)及‘早酥’(非矮生),樹齡20年以上,基砧均為杜梨(Bge.),每個品種3株。分別于2016年4月27日采集‘中矮1號’的花(每花序只采集1朵邊花)和根(基砧杜梨),6月21日采集上述3個品種新梢頂部幼葉、新梢中部的韌皮部和木質部(每個單株采集新梢3個,摘除葉片,剪去枝條基部和頂部幼嫩處,將韌皮部和木質部分離,分別迅速剪碎混勻),6月30日采集‘中矮1號’的果實(每個單株采集果實3個,切成小塊混勻),錫箔紙包裹,液氮速凍后保存于-80℃冰箱中待用。
1.2 梨基因組中ARF家族基因鑒定
由于梨與蘋果均屬于薔薇科仁果類果樹,二者親緣關系較近,因此,本研究以蘋果和模式植物擬南芥中已鑒定出的ARF序列為誘餌,利用NCBI網站的BLAST在線軟件(http://blast.ncbi.nlm.nih.gov/Blast. cgi)對梨基因組(taxid:225117)進行定向tblastn檢索,數據庫選擇RefSeq RNA,從而獲得梨候選的mRNA、蛋白和對應的基因序列。利用SMART(http://smart.embl-heidelberg.de/)和PROSITE(http://prosite.expasy.org/)在線軟件對候選基因的蛋白序列進行分析,以確定其含有所特有的Auxin_resp保守結構域。
1.3 梨ARF蛋白序列的保守結構域及保守元件分析
利用本地軟件DNAMAN v6、在線軟件WebLogo 3(http://weblogo.threeplusone.com/)和MEME Suite 4.11.1(http://meme-suite.org/tools/meme)對梨ARF蛋白序列的保守結構域和保守元件進行進一步分析。
1.4 梨ARF蛋白的進化分析
以獲取的梨ARF蛋白序列和文獻報道的蘋果、葡萄和擬南芥等植物的ARF蛋白序列進行進化樹分析,利用MEGA5軟件構建系統進化樹,采用NJ 方法(執行參數:bootstrap:1 000次重復、Poission model和pairwise deletion)。
1.5 梨ARF的內含子、外顯子結構分析
根據BLAST鑒定獲得梨的CDS序列和對應的基因序列,采用在線軟件GSDS2.0(http://gsds.cbi.pku.edu.cn/)對其進行內含子、外顯子結構分析。
1.6 梨ARF表達分析
將1.1中采自不同單株的相同樣品等量均勻混合研磨,以Plantol高效總RNA提取試劑盒(深圳萊伯克生物科技有限公司)提取各樣品的總RNA,使用DNaseⅠ(大連寶生物科技有限公司)去除總RNA中的DNA,用Prime ScriptTMRT reagent Kit(大連寶生物科技有限公司)對RNA進行反轉錄,采用SYBR?Premix ExTMⅡ(大連寶生物科技有限公司)進行RT-qPCR反應,試驗過程均按各試劑盒的使用說明書進行。RT-qPCR反應采用20 μL體系:SYBR Premix Ex Taq II 10 μL,正、反向引物各1 μL(10 mmol·L-1),cDNA 1 μL,ddH2O 7 μL。反應程序為:95℃,5 min;95℃,30 s;55—60℃(表1),30 s;72℃,30 s,32個循環。以(GU830958.1)為內參,所用引物見表1。采用2-ΔΔCT方法計算各基因的相對表達量。RT-qPCR反應設置3次技術重復,采用DPS 7.05軟件對結果進行差異顯著性分析。
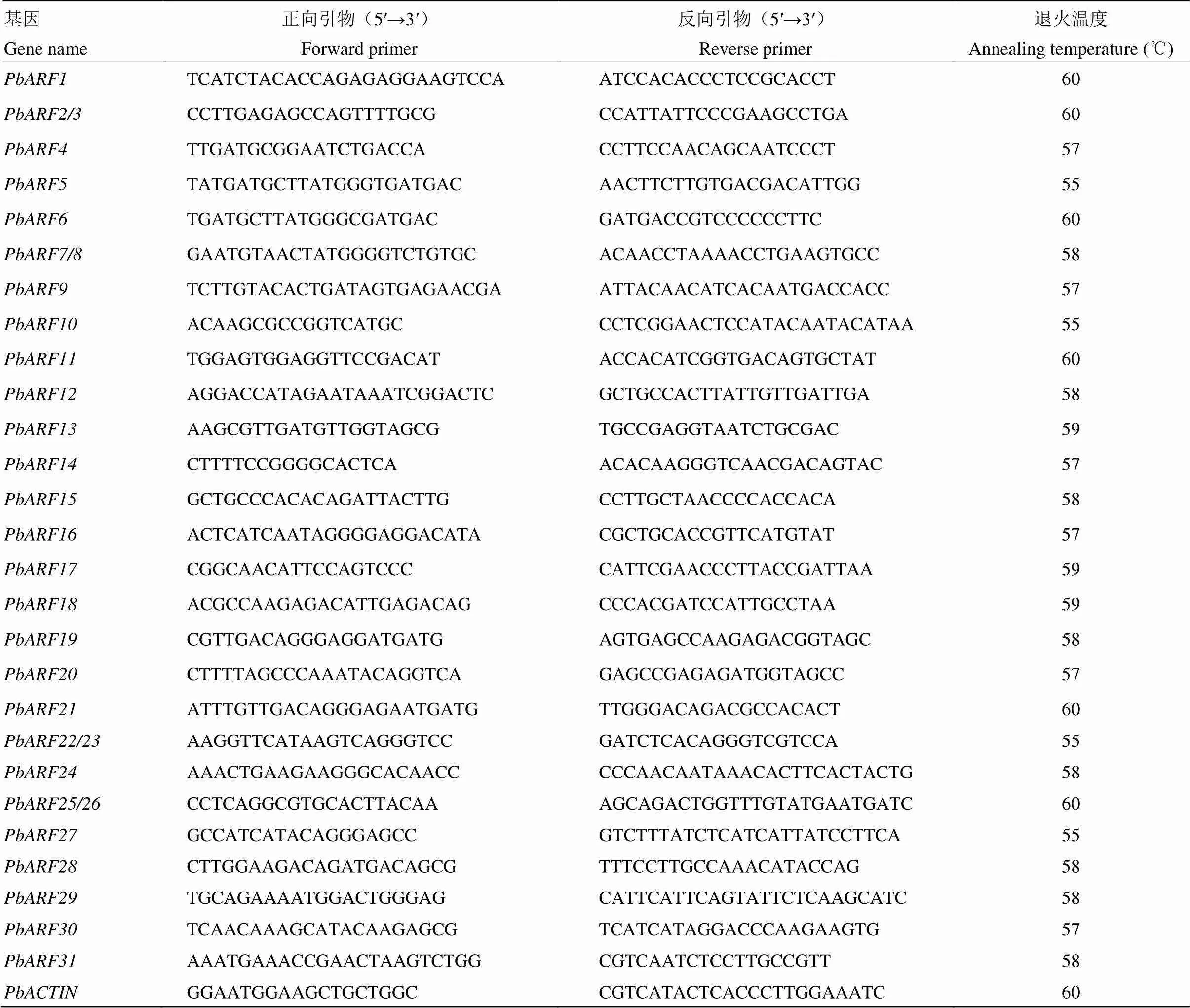
表1 qPCR所用引物序列及退火溫度
2 結果
2.1 梨基因組中ARF家族基因鑒定
根據蘋果和擬南芥ARF蛋白的序列信息,在梨基因組中檢索,并通過結構域分析去除不含Auxin_resp結構域的基因,最終共獲得31個梨,為了便于后續分析,將其以—的方式進行命名,其中,將同一基因位點出現的多個轉錄變異體視為同一基因,后續分析時僅以轉錄變異體(transcript variant)X1對應的序列作為該基因的代表。分析表明,的長度在587—1 154 aa,分子量在64.78—128.88 kD,等電點在5.07—7.82(表2),除和、和等2對序列完全相同外,其他序列均各不相同。

表2 梨ARF
aNCBI網站GenBank數據庫中預測的mRNA序列編號;b類生長素反應因子2轉錄變異體
aThe accession number of predicted mRNA sequence from NCBI`s GenBank database;bAuxin response factor 2-like transcript variant
2.2 梨ARF結構域分析
通過SMART和PROSITE在線軟件對所獲得的蛋白序列進行檢索分析,所有候選蛋白的N端除含有1個AFR蛋白所特有的Auxin_resp結構域外,還含有1個B3結構域,除PbARF11、12、24、25和26外,其他序列的C末端還存在一個PB1(AUX_IAA)結構域。通過DNAMAN比對和Weblogo在線分析,發現上述3個結構域在梨中高度保守。其中,B3結構域中有40個氨基酸在所有序列中是完全保守的,約占結構域總長度(103 aa)的38.8%;Auxin_resp結構域中有9個氨基酸在所有序列中是完全保守的,約占結構域平均長度(83 aa)的10.8%;PB1結構域中有10個氨基酸在含有該結構域的序列中是完全保守的,約占結構域平均長度(83 aa)的12.0%(圖1)。
利用在線軟件MEME對梨ARF蛋白序列進行分析,結果顯示,梨ARF蛋白含有15個保守元件。其中,元件1、2和10屬于B3結構域,它們與元件3和元件5共同構成DNA結合結構域;元件6、7、8和14屬于Auxin_resp結構域;元件4、9和15屬于PB1結構域;元件11、12和13為未知結構域。除PbARF18缺失元件3和5外,所有序列均含有DNA結合結構域的5個元件和Auxin_resp結構域4個元件中的至少3個元件(PbARF5—8缺失元件8,PbARF24—31缺失元件14)。PbARF1—10和13—23含有PB1結構域中的所有3個元件,而PbARF11和12則缺失了其中的元件4和9,PbARF27—31則缺失了其中的元件15,PbARF27—28除缺失元件15外,還缺失元件4。此外,除PbARF11和12外,所有序列均含有未知元件11,而未知元件12僅在PbARF24和25—31中出現,未知元件13只在PbARF13和15—24中出現(圖2)。
2.3 梨ARF蛋白的進化分析
利用MEGA5.05軟件對本研究獲得的31個梨及文獻報道的29個蘋果[13]、19個葡萄[12]和23個擬南芥[6]的共102個AFR氨基酸序列進行進化分析。結果表明,所有102個ARF成員可以被清晰分成4組:ClassⅠ、ClassⅡ、ClassⅢ和ClassⅣ,每組中都含有上述4個物種中的成員。其中,PbARF1—8等8個成員在ClassⅠ中,PbARF9—12等4個成員在ClassⅡ中,PbARF13—23等11個成員在ClassⅢ中,PbARF24—31等8個成員在ClassⅣ中。在所有31個PbARF中,除PBARF19、20、27和28等4個成員外,其他成員均能在蘋果中找到與其配對的同源序列,蘋果中也僅有MdARF1、4、5和14等4個成員在梨中沒有與其配對的同源序列。在ClassⅠ中,AtARF12—15、20—23等8個擬南芥成員單獨聚在一個小分支上,在梨、蘋果和葡萄等其他3種果樹中均未發現與其高度同源的序列(圖3)。
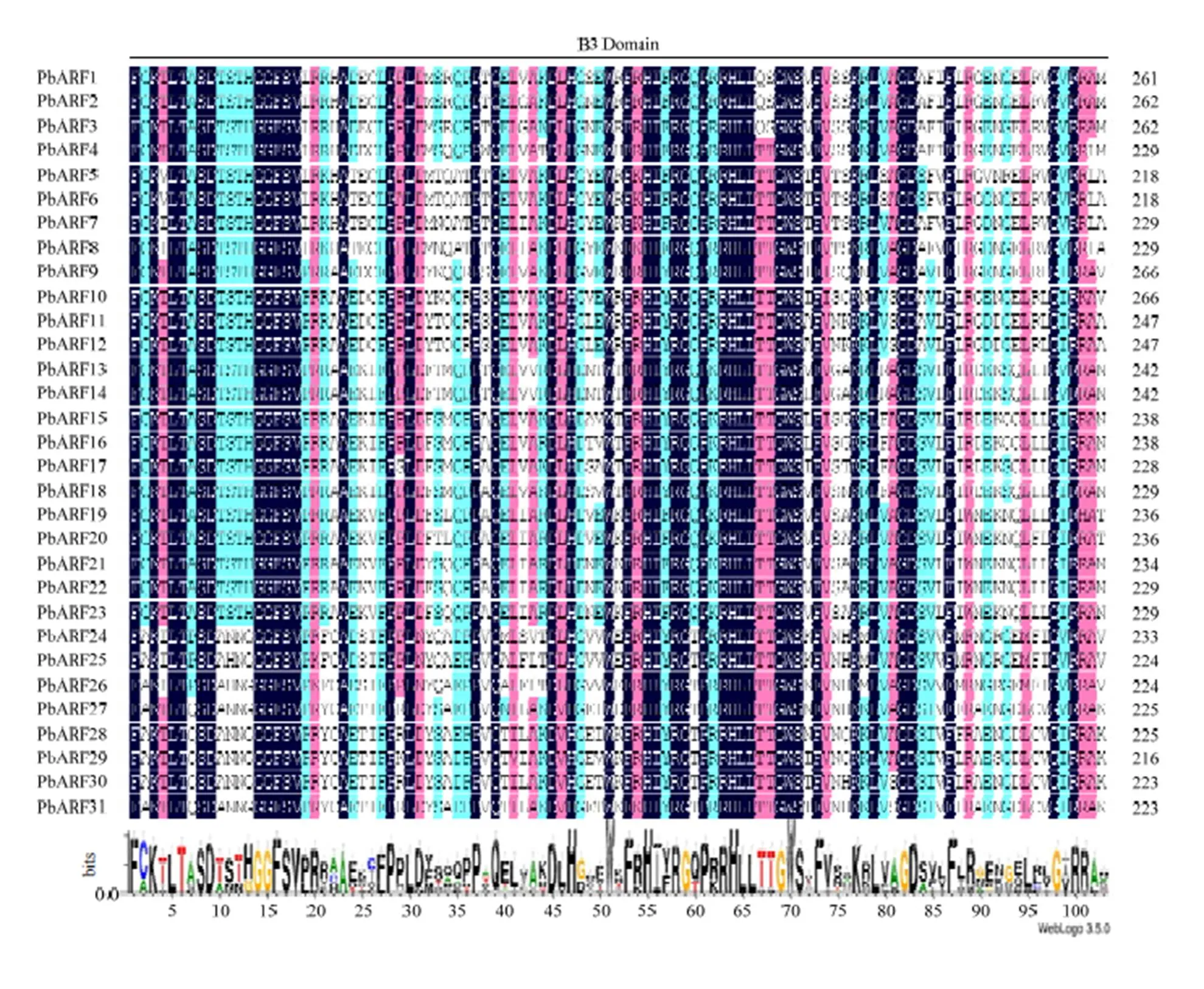
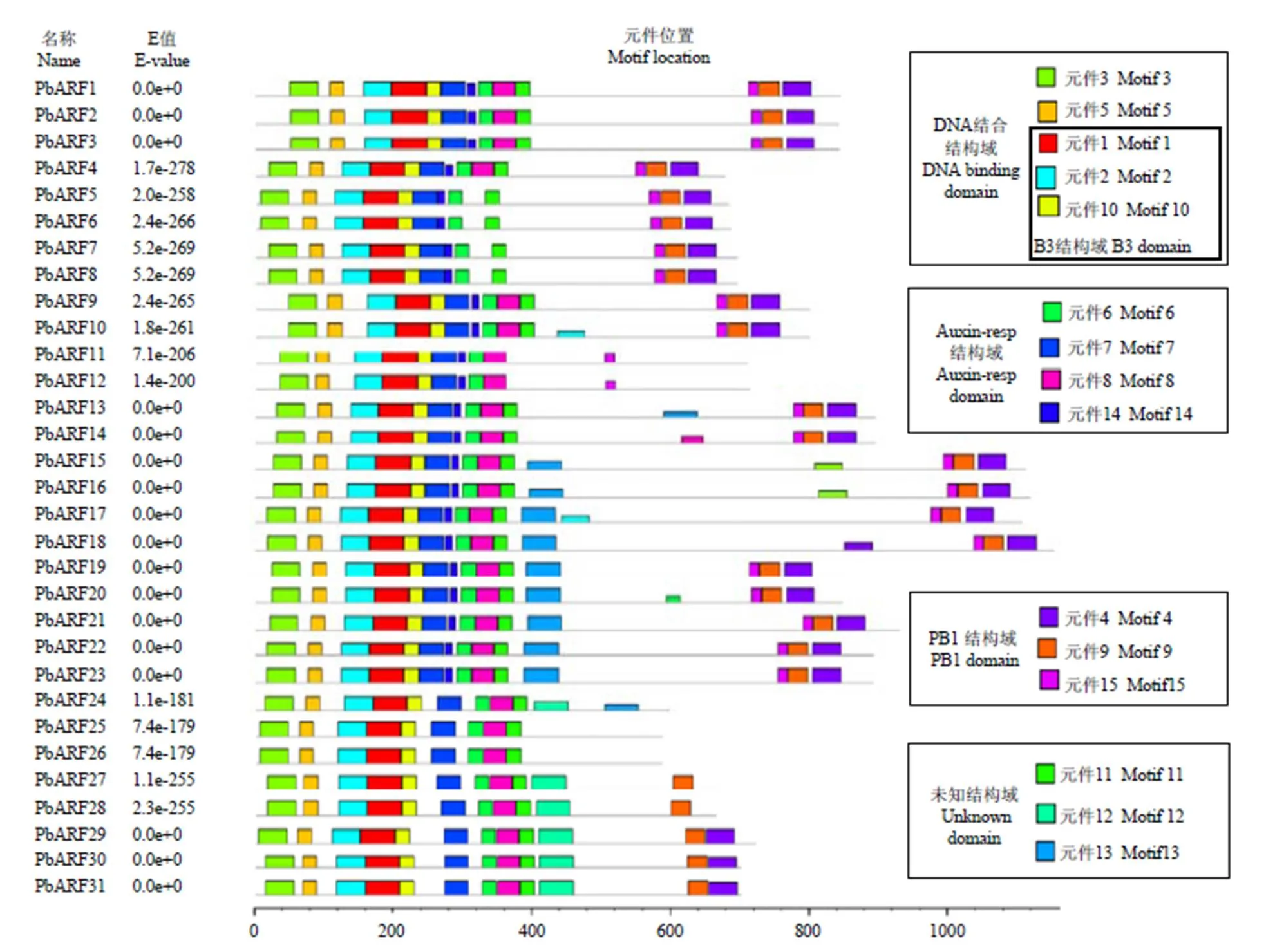
所有保守元件(Motif 1—15)都是利用在線軟件MEME檢索梨ARF蛋白序列獲得,不同顏色矩形塊的長度和排列順序代表各保守元件在蛋白序列上的實際大小和分布位置
2.4 梨ARF的內含子、外顯子結構分析
對獲得的31個PbARF蛋白序列進行了進一步的進化分析,發現這些蛋白也可以被清晰的聚為4組,這與圖3的分組結果基本一致,只是不同組間的進化關系略有不同。對獲得的的CDS序列和對應的基因序列采用在線軟件GSDS2.0(http:// gsds.cbi.pku.edu.cn/)進行內含子和外顯子結構分析,發現家族基因外顯子的數量在2—15,其中ClassⅣ成員的外顯子數量最少,僅為2—3個,ClassⅡ成員的外顯子數量為11—12,ClassⅠ和ClassⅢ成員的外顯子數量多為14個,僅個別成員為13或15個(圖4)。
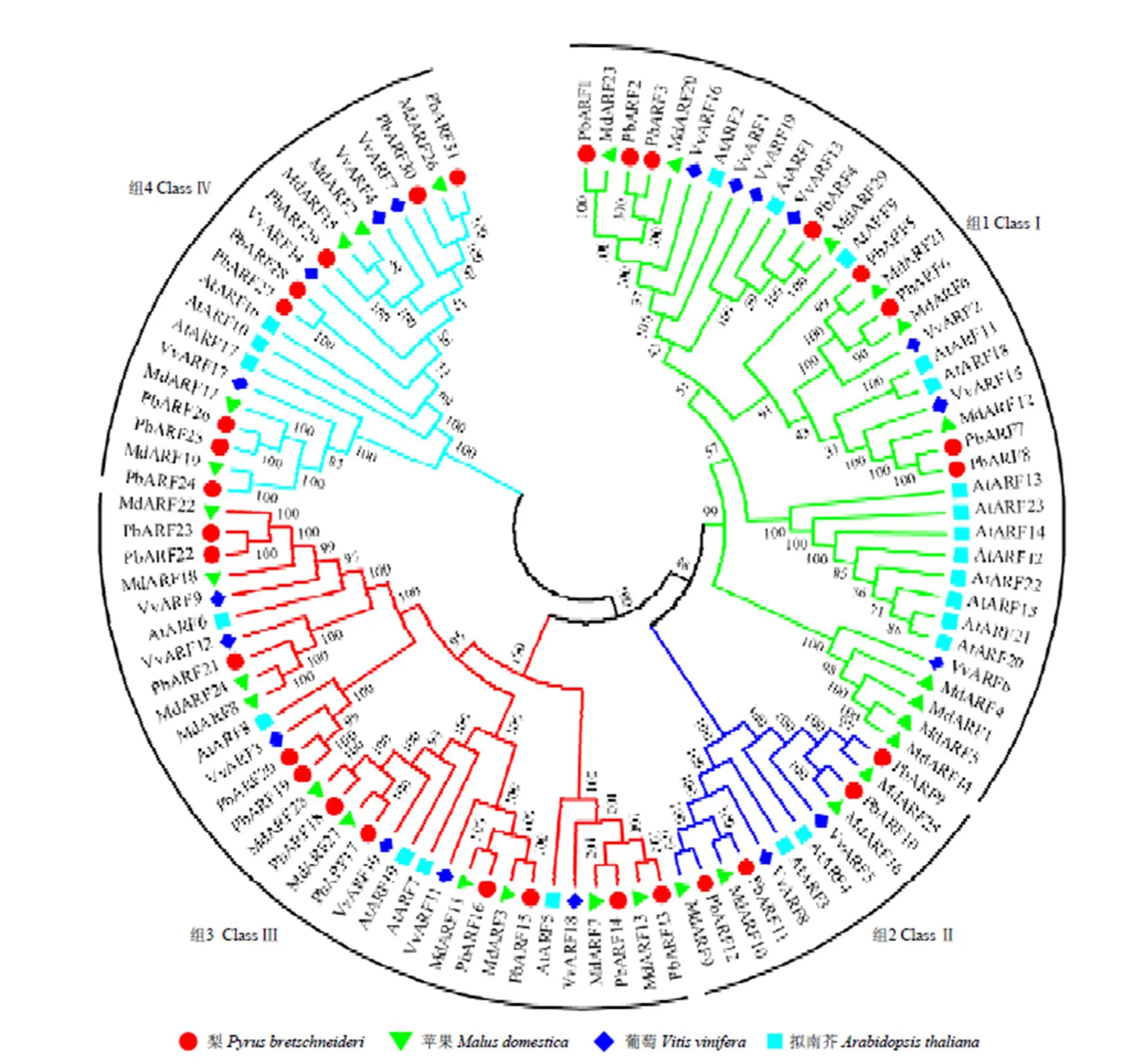
該進化樹是利用MEGA5軟件采用NJ方法構建的,bootstrap測驗采用1 000次重復,節點上的數字代表分枝的可信度,不同分組ARF蛋白的分枝顏色不同
2.5 梨ARF表達分析
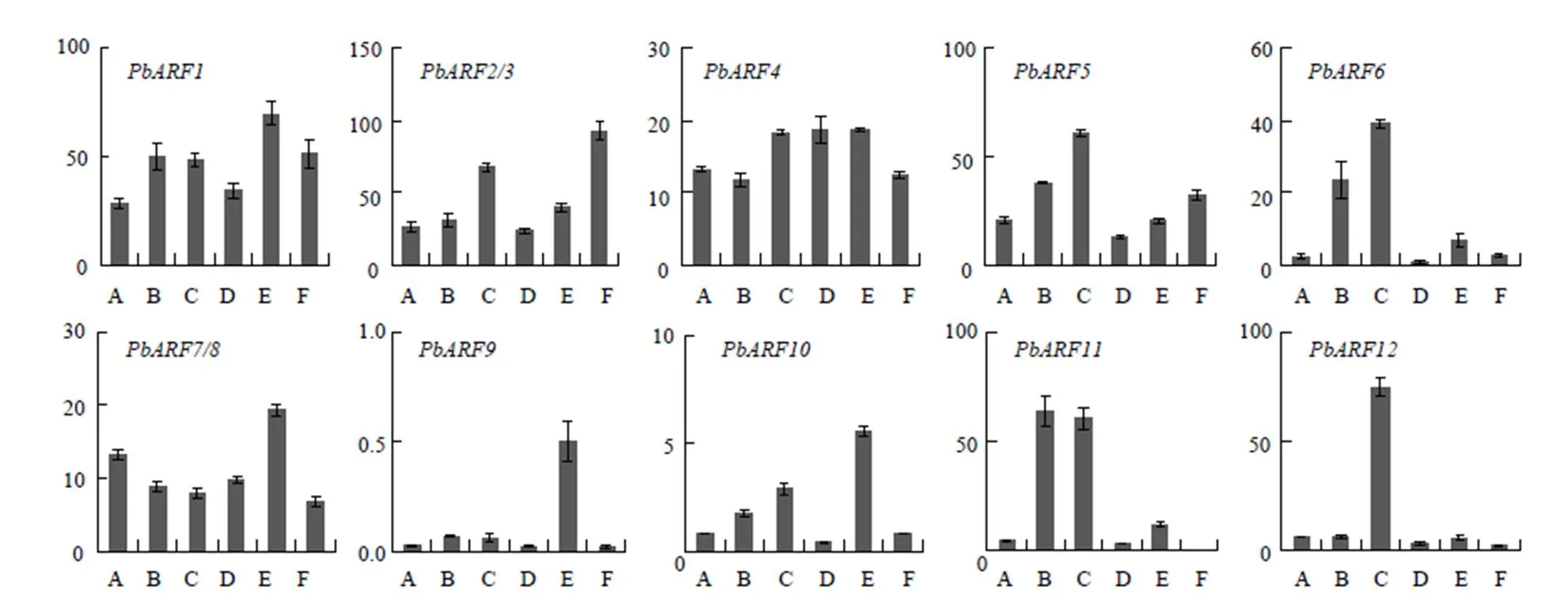
運用熒光定量PCR的方法對基因家族成員在矮生梨品種‘中矮1號’不同組織器官中的表達情況進行分析。由于和和和和等4對基因的序列相同或或高度相似,難以設計特異表達引物加以區分,因此,表達量的檢測結果的是1對基因的共同表達量。組織表達結果表明,所檢測的基因家族成員幾乎在‘中矮1號’所有組織中均有表達。其中,相對于其他成員基因表達量較低;和在果實中的表達量相對高于其他組織;在花中的表達量相對高于其他組織;等7個成員則在木質部中的表達量相對較高,除在木質部中表達相對較高外,在韌皮部中的表達量也相對較高(圖5)。
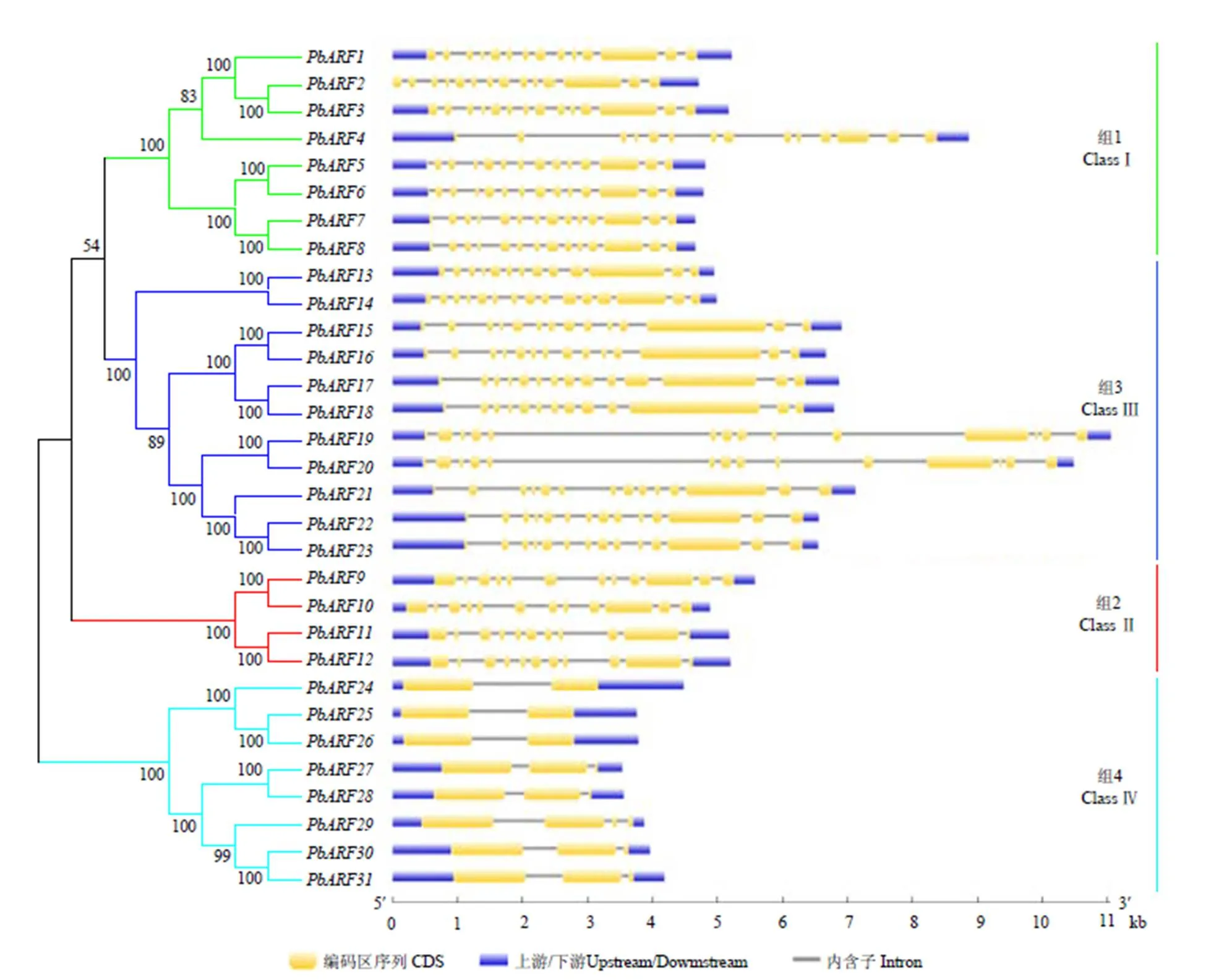
進化樹的構建方法同圖3;內含子和外顯子分布結構圖由在線軟件GSDS2.0生成,矩形塊的長度代表其實際的序列長度
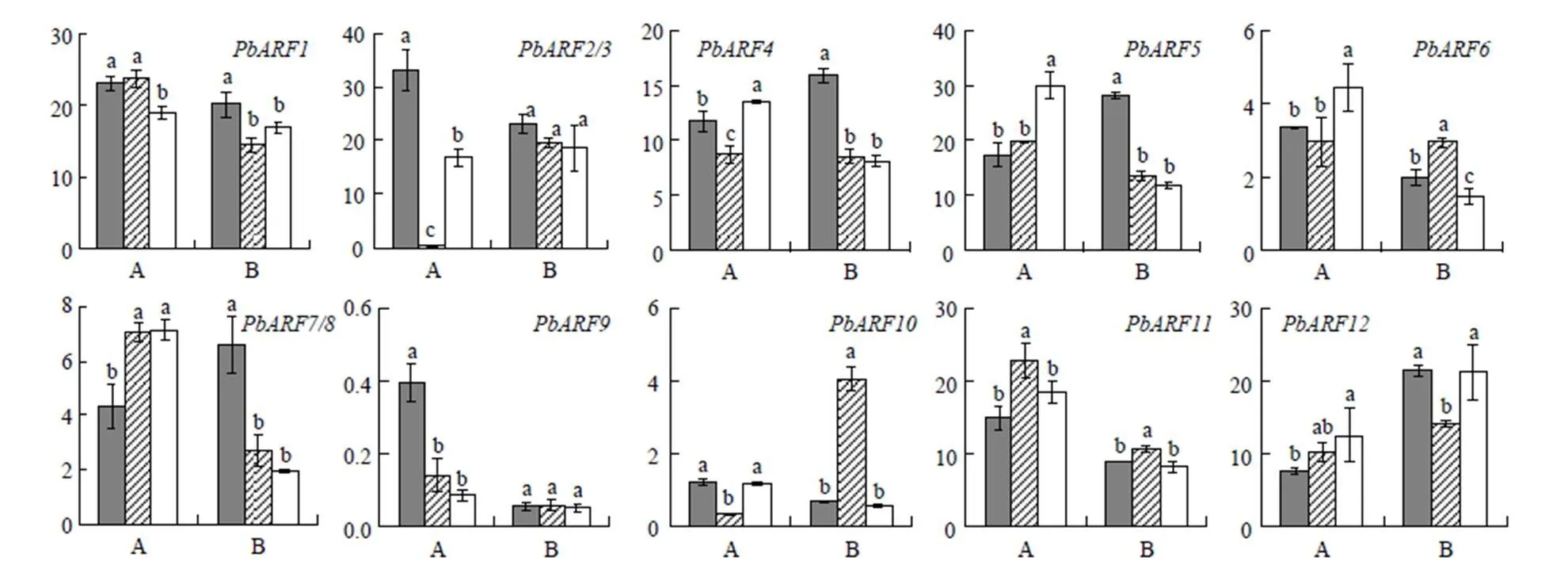
進一步比較基因家族成員在‘早酥’(喬化)、‘錦香’(‘中矮1號’母本,半矮化)和‘中矮1號’(矮化)等3個梨品種木質部和韌皮部中的表達情況。結果表明,不同在3個品種韌皮部和木質部中的表達各有高低,多無明顯規律,僅在3個品種韌皮部中的表達表現為植株越矮表達量越高的趨勢,等4個基因在3個品種木質部中的表達表現為植株越矮表達量越低的趨勢(圖6)。
3 討論
隨著高通量測序和生物信息技術的發展,越來越多物種的基因組序列已得到解析,使得從基因組層面上對某一家族基因進行全面的鑒定和分析成為可能。ARF是生長素信號途徑中一類重要的轉錄因子,其在基因組中即以家族的形式存在,分別調控植物生長發育的許多重要環節。ARF基因家族成員已在多個物種中被分離鑒定出來,但不同物種中ARF基因家族成員數量不同,本研究自梨基因組中鑒定出PbARF基因31個,與玉米[7]中的成員數量一致,比擬南芥、蘋果[6,8,10-15]等多個物種中的成員數量均多,但少于大豆(51個)[9]和香蕉(47個)[16]。雖然梨的數量與玉米中的數量一樣多,但其基因組大小為527 Mb[17],遠小于玉米的2 300 Mb[18];雖然基因組大小與香蕉的523 Mb[19]接近,但數量卻比香蕉中的少的多。由此可見,在基因組中的數量多少不完全取決于基因組大小。有研究表明基因復制對基因家族的形成起著重要作用[20],在蘋果中的研究即發現串聯復制、片段復制和全基因組復制對蘋果基因組的擴展具有重要作用[13]。梨與蘋果同屬薔薇科果樹,ARF蛋白的進化分析結果表明,二者的親緣關系較近,雖然梨基因組小于蘋果(742 Mb[21]),但二者ARF基因數量接近,僅各有4個ARF蛋白沒有形成互相配對的同源序列,其他成員均能在另一個基因組中找到與之配對的同源序列。因此,推測梨基因組中的進化也經歷了與蘋果類似的過程,但由于基因組大小和組成不同,個別基因的進化過程和功能可能又略有不同。
通過系統進化和基因內含子、外顯子組成結構等分析,本研究中的PbARF蛋白可被劃分為4組,該分組結果與水稻、葡萄和甜橙等[8,12,19]的研究結果一致,與蘋果、玉米等[7,19]的研究結果略有不同,其不同的主要原因是他們對成員較多的組又進行了細致劃分。但與梨一致的是,在其他物種中也有一個外顯子數量較少的分組,其他分組成員的外顯子數量均較多,可見,在外顯子和內含子進化上具有較大的一致性。
本研究鑒定的梨AFR蛋白多含有3個保守結構域,1個AFR蛋白所特有的Auxin_resp結構域、1個B3結構域和1個PB1結構域,但PbARF11、12、24、25和26等5個成員僅有2個保守結構域,PB1結構域缺失,與前人研究結果類似[22]。研究表明,B3結構域具有DNA結合活性[23],其可以與生長素反應基因啟動子上的作用元件結合,進而通過與B3結構域相連的Auxin_resp結構域激活或抑制相關基因的表達[3]。PB1結構域是一個介導蛋白質-蛋白質互作的結構域,另一個在生長素信號途徑中起抑制作用的AUX/IAA蛋白中也具有類似結構域,2個蛋白可以通過該結構域的互作而形成ARF-ARF、ARF-Aux/IAA和 Aux/IAA-Aux/IAA同源或異源寡聚體。當生長素濃度低時,ARF與Aux/IAA形成異源寡聚體,抑制生長素反應基因的表達;當生長素濃度升高時,Aux/IAA蛋白被泛素化途徑降解,抑制作用解除[22]。但PbARF11、12、24、25和26等PB1結構域缺失的ARF是否有類似的功能還有待進一步研究。
熒光定量PCR表達分析結果表明,本研究中各在梨樹根、韌皮部、木質部、葉、花和果實等不同組織器官中均可表達,在蘋果[13]和番茄[24]等作物上也獲得了類似結果。通過對擬南芥突變體的研究表明,可以對開花時間和花器官的質量產生影響[25],本研究中PbARF19—23與其處于同一進化分枝,其中,即在花器官中具有相對較高的表達量,可能與具有類似的功能。對根冠的形成具有重要作用[26],本研究中PbARF27—31與其處于較近的分枝上,在根中均具有相對較高的表達量,可能與根的發育相關。在側根的形成、根系和下胚軸的向重力性中起重要作用[27],番茄中的同源基因則在花中具有較高的表達量,調控授粉受精過程[28],本研究中PbARF17、18與AtARF7、19處于較近的分枝上,在花中均有相對較高的表達量,與番茄中的結果一致,其功能或許與擬南芥不同。
‘中矮1號’不僅自身矮化,其作中間砧對嫁接品種也具有矮化作用,而中間砧僅由木質部和韌皮部兩部分組成,可見控制其矮化性狀的基因只能在木質部和韌皮中起作用;前期研究結果表明,新梢停長早是‘中矮1號’矮生的原因之一,6月中下旬是‘中矮1號’即將停長的關鍵時期[29]。因此,本研究還于6月21日采集3個不同生長勢梨品種新梢的木質部和韌皮部,比較了在其中的表達情況,發現在3個品種韌皮部中的表達表現為植株越矮表達量越高的趨勢,等4個基因在3個品種木質部中的表達表現為植株越矮表達量越低的趨勢。PbARF16、17、18與AtARF7和AtARF19處于同一分枝,PbARF27、29與AtARF10和AtARF16處于較近分枝上,研究表明,上述基因在擬南芥中均可對根系的生長發育產生影響[26-27],其中,AtARF7和AtARF19功能有重疊,擬南芥和雙突變體具有側根發育嚴重缺陷和根及下胚軸向地性異常等表型,但各自的單突變體均無此表型[27];番茄中的同源基因具有調控果實授粉受精的功能[28];和也具有一定程度的功能冗余,他們均為microRNA160的靶基因,受其負調控,主要影響根冠的發育,同時,還在種子的萌發過程中起重要作用[30],但均未見對植株高度影響的報道。盡管如此,由于梨樹是木本植物,與擬南芥和番茄等物種的差異極大,這些基因是否能夠影響梨樹植株的生長還需進一步研究。
4 結論
梨基因組中含有31個ARF基因家族成員;所有PbARF蛋白均含有Auxin_resp和B3等2個高度保守的結構域;PbARF蛋白可以被劃分為4組,基因結構進化高度保守;所有在‘中矮1號’不同組織器官及‘早酥’(喬化)、‘錦香’(‘中矮1號’母本,半矮化)和‘中矮1號’(矮化)3個梨品種的木質部和韌皮部中均有表達,其中,等5個基因的表達可能與梨樹植株的高矮有關。
[1] 姜淑苓, 賈敬賢, 紀寶生, 馬力. 梨矮化砧木—中矮1 號. 中國果樹, 2000(3): 1-3.
Jiang S L, Jia J X, Ji B S, Ma L. Pear dwarfing stocks-Zhongai 1., 2000(3): 1-3. (in Chinese)
[2] Vanneste S, Friml J. Auxin: a trigger for change in plant development., 2009, 136(6): 1005-1016.
[3] GUILFOYLE T J, HAGEN G. Auxin response factors., 2007, 10(5): 453-460.
[4] HAGEN G, GUILFOYLE T. Auxin-responsive gene expression: genes, promoters and regulatory factors., 2002, 49(3/4): 373-385.
[5] ULMASOV T, HAGEN G, GUILFOYLE T J. ARF1, a transcription factor that binds to auxin response elements., 1997, 276(5320): 1865-1868.
[6] REMINGTON D L, VISION T J, GUILFOYLE T J, REED J W. Contrasting modes of diversification in the Aux/IAA and ARF gene families., 2004, 135(3): 1738-1752.
[7] XING H Y, PUDAKE R N, GUO G G, XING G F, HU Z R, ZHANG Y R, SUN Q X, NI Z F. Genome-wide identification and expression profiling of auxin response factor (ARF) gene family in maize. BMC Genomics, 2011, 12(1): 178.
[8] WAN D K, PEI K M, FU Y P, SUN Z X, LI S J, LIU H Q, TANG K, HAN B, TAO Y Z. Genome-wide analysis of the auxin response factors (ARF) gene family in rice (Oryza sativa). Gene, 2007, 394(1): 13-24.
[9] HA C V, LE D T, NISHIYAMA R, WATANABE Y, SULIEMAN S, TRAN U T, MOCHIDA K, DONG N V, YAMAGUCHI-SHINOZAKI K, SHINOZAKI K, TRAN L P. The auxin response factor transcription factor family in soybean: genome-wide identification and expression analyses during development and water stress. DNA Research, 2013, 20(5): 511-524.
[10] KUMAR R, TYAGI A K, SHARMA A K. Genome-wide analysis of auxin response factor (ARF) gene family from tomato and analysis of their role in flower and fruit development. Molecular Genetics and Genomics, 2011, 285(3): 245-260.
[11] 盛慧, 秦智偉, 李文濱, 周秀艷, 武濤, 辛明. 黃瓜生長素反應因子(ARF)家族鑒定及表達特異性分析. 中國農業科學, 2014, 47(10): 1985-1994.
SHENG H, QIN Z W, LI W B, ZHOU X Y, WU T, XIN M. Genome-wide identification and expression analysis of auxin response factor (ARF) family in cucumber. Scientia Agricultura Sinica, 2014, 47(10): 1985-1994. (in Chinese)
[12] WAN S B, LI W L, ZHU Y Y, LIU Z M, HUANG W D, ZHAN J C. Genome-wide identification, characterization and expression analysis of the auxin response factor gene family in Vitis vinifera. Plant Cell Reports, 2014, 33(8): 1365-1375.
[13] 李慧峰, 冉昆, 何平, 王海波, 常源升, 孫清榮, 程來亮, 李林光. 蘋果生長素響應因子(ARF)基因家族全基因組鑒定及表達分析. 植物生理學報, 2015, 51(7): 1045-1054.
LI H F, RAN K, HE P, WANG H B, CHANG Y S, SUN Q R, CHENG L L, LI L G. Genome-wide identification and expression analysis of auxin response factor (ARF) gene family in apple. Plant Physiology Journal, 2015, 51(7): 1045-1054. (in Chinese)
[14] LI S B, OUYANG W Z, HOU X J, XIE L, HU C G, ZHANG J Z. Genome-wide identification, isolation and expression analysis of auxin response factor (ARF) gene family in sweet orange (Citrus sinensis). Frontiers in Plant Science, 2015, 6: 119.
[15] LIU K D, YUAN C C, LI H L, LIN W H, YANG Y J, SHEN C J, ZHENG X L. Genome-wide identification and characterization of auxin response factor (ARF) family genes related to flower and fruit development in papaya (Carica papaya L.). BMC Genomics, 2015, 16(1): 901.
[16] HU W, ZUO J, HOU X W, YAN Y, WEI Y X, LIU J H, LI M Y, XU B Y, JIN Z Q. The auxin response factor gene family in banana: genome-wide identification and expression analyses during development, ripening, and abiotic stress. Frontiers in Plant Science, 2015, 6: 742.
[17] WU J, WANG Z W, SHI Z B, ZHANG S, MING R, ZHU S L,KHAN M A, TAO S T, KORBAN S S, WANG H, CHEN N J, NISHIO T, XU X, CONG L, QI K J, et al. The genome of the pear (Pyrus bretschneideri Rehd.). Genome Research, 2013, 23(2): 396-408.
[18] SCHNABLE P S, WARE D, FULTON R S, STEIN J C, WEI F S, PASTERNAK S, LIANG C Z, ZHANG J W, FULTON L, GRAVES T A, WILSON R K. The B73 maize genome: complexity, diversity, and dynamics. Science, 2009, 326(5956): 1112-1115.
[19] D’HONT A, DENOEUD F, AURY J M, BAURENS F C, CARREEL F, GARSMEUR O, NOEL B, BOCS S, DROC G, ROUARD M, WINCKER P. The banana (Musa acuminata) genome and the evolution of monocotyledonous plants. Nature, 2012, 488(7410): 213-217.
[20] FLAGEL L E, WENDEL J F. Gene duplication and evolutionary novelty in plants., 2009, 183(3): 557-564.
[21] VELASCO R, ZHARKIKH A, AFFOURTIT J, DHINGRA A, CESTARO A, KAYANARAMAN A, FONTANA P, BHATNAGAR S K, TROGGIO M, PRUSS D, VIOLA R. The genome of the domesticated appleBorkh.)., 2010, 42(10): 833-839.
[22] GUILFOYLE T J. The PB1 domain in auxin response factor and Aux/IAA proteins: a versatile protein interaction module in the auxin response., 2015, 27(1): 33-43.
[23] SUZUKI M, KAO C Y, MCCARTY D R. The conserved B3 domain of VIVIPAROUS1 has a cooperative DNA binding activity., 1997, 9(5): 799-807.
[24] KUMAR R, TYAGI A K, SHARMA A K. Genome-wide analysis of auxin response factor (ARF) gene family from tomato and analysis of their role in flower and fruit development., 2011, 285(3): 245-260.
[25] NAGAL P, ELLIS C M, WEBER H, PLOENSE S E, BARKAWI L S , GUILFOYLE T J, HAGEN G, ALONSO J M, COHEN J D, FARMER E E, ECKER J R, REED J W. Auxin response factors ARF6 and ARF8 promote jasmonic acid production and flower maturation., 2005, 132(18): 4107-4118.
[26] WANG J W, WANG L J, MAO Y B, CAI W J, XUE H W, CHEN X Y. Control of root cap formation by microRNA-targeted auxin response factors in., 2005, 17(8): 2204-2216.
[27] OKUSHIMA Y, OVERVOORDE P J, ARIMA K, ALONSO J M, CHAN A, CHANG C, ECKER J R, HUGHES B, LUI A, NGUYEN D, ONODERA C, QUACH H, SMITH A, YU G X, Theologis A. Functional genomic analysis of the AUXIN RESPONSE FACTOR gene family members in: unique and overlapping functions of ARF7 and ARF19., 2005, 17(2): 444-463.
[28] DE JONG M, WOLTERS-ARTS M, FERON R, MARIANI C, VRIEZEN W H. The Solanum lycopersicum auxin response factor 7 () regulates auxin signaling during tomato fruit set and development., 2009, 57: 160-170.
[29] OU C Q, JIANG S L, WANG F, TANG C Y, HAO N N. An RNA-Seq analysis of the pearL.) transcriptome, with a focus on genes associated with dwarf., 2015, 4: 69-77.
[30] LIU P P, MONTGOMERY T A, FAHLGREN N, KASSCHAU K D, NONOGAKI H, CARRINGTON J C. Repression of AUXIN RESPONSE FACTOR10 by microRNA160 is critical for seed germination and post-germination stages., 2007, 52(1): 133-146.
(責任編輯 趙伶俐)
Genome-wide Identification and Expression Analysis of() Gene Family in Pear
OU ChunQing, JIANG ShuLing, WANG Fei, ZHAO YaNan
(Institute of Pomology, Chinese Academy of Agricultural Sciences/Key Laboratory of Horticultural Crops Germplasm Resources Utilization, Ministry of Agriculture, Xingcheng 125100, Liaoning)
【Objective】The objectives of this research are to identify the auxin response factor (ARF) family genes from pear ()genome, to know the profile offamily such as gene number, gene structure and tissue expression in pear, and to provide theoretical basis for revealing what roles theplay in auxin signaling pathway and in growth and development of dwarf pear.【Method】genes in pear genome were identified by BLAST software based ongenes from apple and. SMART, PROSITE, WebLogo 3, DNAMAN 5, MEME, GSDS 2 and MEGA 5.1 software were used for bioinformatics analysis of ARF protein and gene sequences. The qPCR method was used to detect the relative expression ofgenes in different tissues of dwarf pear ‘Zhongai 1’ and in xylem and phloem of 3 pear cultivars with different growth vigor.【Result】Total of 31genes were identified from pear genome. All the PbARFs contain two domains of Auxin_resp and B3, and except for PbARF11, 12, 24, 25 and 26, the rest also contain a PB1 domain. Conservative motif analysis result showed that there are 15 motifs in PbARFs, but not every PbARF protein contains all the motifs. The PbARFs were divided into four classes based on phylogenetic analysis. Gene structure analysis result showed that there are 2-15 exons ins, The gene structure ofis high conservative. The qPCR result showed that all thegenes were expressed in the root, phloem, xylem, leaf, flower and fruit of ‘Zhongai 1’ pear and the expression pattern was various. The relative expression of,,,in the xylem of 3 pear cultivars showed that the more dwarf the plant, the lower the expression.【Conclusion】Auxin response factor family in pear contains 31 genes. All the 31 PbARF proteins contain both of Auxin_resp and B3 conservative domains, and were divided into four classes. The gene structure ofis high conservative. All the 31s were expressed in different tissues of ‘Zhongai 1’, root of rootstockand in phloem and xylem of 3 cultivars. Thereinto, the expression of,,,,may be relevant to pear plant height.
pear; ARF; transcription factor; bioinformatics; gene expression
2017-06-26;
2017-09-12
國家科技支撐計劃(2013D02B01,2013BAD01B04-23)、中國農業科學院科技創新工程(CAAS-ASTIP-2016-RIP)、中央級公益性科研院所基本科研業務費專項(Y2016CG19 1610032012006)、遼寧省果樹產業技術體系專項(LNGSCYTX-15-X)
歐春青,E-mail:ochunqing@163.com。
姜淑苓,E-mail:jshling@163.com
猜你喜歡
現代畜牧科技(2021年9期)2021-10-13 06:39:14
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
當代經濟研究(2016年5期)2016-12-01 03:12:05
現代農業(2016年5期)2016-02-28 18:42:46
出版與印刷(2016年3期)2016-02-02 01:20:11
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
華北水利水電大學學報(社會科學版)(2014年3期)2014-04-16 04:38:31
終身教育研究(2014年5期)2014-02-28 01:23:06
