紫花苜蓿MsSOS1基因的克隆與表達分析
2018-02-13 01:42:52方志紅崔苗苗任彬琳石永紅劉建寧王運琦董寬虎高洪文吳欣明
草地學報 2018年6期
方志紅, 崔苗苗, 張 燕, 任彬琳, 石永紅, 劉建寧,王運琦,董寬虎, 高洪文, 吳欣明*
(1. 山西省農業科學院畜牧獸醫研究所, 山西 太原 030032; 2. 中國農業科學院北京畜牧獸醫研究所, 北京 100193;3. 山西農業大學動物科技學院, 山西 太谷 030801)
鹽脅迫是影響植物生長發育和產量的重要因素之一[1],全世界約有8億公頃的土地被鹽化,其中約有五分之一的可耕地受到鹽脅迫的危害[2-3],土壤中的Na+是影響植物生長的主要因子,高濃度的Na+會對植物體造成生理性干旱和離子毒害[4],從而抑制植物的生長甚至會引起植物死亡[5-6],為了生存,植物會利用各種各樣的機制來抵御環境中的有害因子[7-8]。SOS(salt overly sensitive)信號通路是植物抵御鹽脅迫的一個至關重要的途徑,它主要揭示了在鹽脅迫下植物體內關于離子穩態調節和耐鈉性的機理[9],SOS1、SOS2和SOS3在一條共同的SOS信號轉導途徑中起作用,通過對該通路中SOS1、SOS2和SOS3突變體的研究發現,SOS1(Salt Overly Sensitive 1)是植物在抵御鹽脅迫的過程中必需的一個重要的、與植物耐鹽性關系最直接的一個基因[4],它編碼的SOS1蛋白可以將植物體內的多余的Na+進行區隔化或外排來降低細胞中的Na+濃度來適應高濃度的鹽環境[3,10-12],從而降低鹽脅迫的傷害[13-15]。
已經有很多研究表明,SOS1基因在植物的耐鹽方面起著重要作用,它主要在介導Na+外排、參與Na+的長距離運輸、介導K+的吸收或轉運和調節細胞質pH幾個方面起著重要的作用[16-18]。近些年來,質膜Na+/H+逆向轉運蛋白(SOS1)在植物耐鹽性研究中的作用受到廣泛重視,SOS1基因最初是在擬南芥中被發現的,擬南芥SOS1突變體對鹽超敏感,對其進行過表達發現可以提高轉基因擬南芥的耐鹽性,表明它在植物的耐鹽機制中起著非常重要的作用[19-20]。其后許多植物的SOS1基因已經被克隆鑒定和進行了功能分析,如十字花科植物擬南芥(Arabidopsisthaliana)[21]、鹽芥(Thellungiellasalsuginea)[22]、芥菜(Brassicajuncea)[23],楊柳科植物胡楊(Populuseuphratica)[24],藜科植物鹽地堿蓬(Suaedasalsa)[25],番杏科植物海馬齒(Sesuviumportulacastrum)[26],蒺藜科植物霸王(Zygophyllumxanthoxylum)[27],禾本科植物小麥(Triticumaestivum)[28]、水稻(Oryzasativa)[29]、大麥(Hordeumvulgare)[30]、獐茅(Aeluropuslittoralis)[31],茄科植物番茄(Lycopersiconesculentum)[32],豆科植物大豆(GlycinemaxandGlycinesoja)[33]和草木樨(Melilotusofficinalis)[34]。這些研究都可以表明,在鹽脅迫下SOS1蛋白可以通過減少根部Na+的累積來提高植物的耐鹽性。很多實驗研究也都可以表明SOS1基因可以恢復并使其轉化體的耐鹽能力增強[26,28,33,35-41]。
雖然SOS1基因在很多植物中被克隆鑒定,但是在豆科植物中研究的比較少,只是在大豆和草木樨中被報道,紫花苜蓿SOS1基因的研究還未見報道。紫花苜蓿易于栽培、產量高,是可以為家畜提供優良蛋白飼料的重要豆科牧草[42-43],但是在鹽堿地上不能大面積的種植[44],為了解決這一問題,利用生物技術手段培育抗逆性強的品種則成為一條切實可行的有效途徑[45],本研究利用同源克隆的方法獲得了紫花苜蓿MsSOS1基因的全長cDNA序列并對其進行了分析和不同逆境脅迫下的表達分析,以期為探討SOS1基因所在的SOS途徑在紫花苜蓿中的作用機制和合理開發利用紫花苜蓿培育抗逆新品種提供理論依據。
1 材料與方法
1.1 試驗材料
以紫花苜蓿(Medicagosativa)種子(中苜1號,北京畜牧獸醫研究所提供)為實驗材料。實驗所需的試劑為ExTaq(Takara)、DNA Maker(Takara)、pMD?18-Tvector(Takara),Total RNA(Invitrogen),RevertAidTMFirstStrand cDNA Synthesis Kit(Fermentas公司),大腸桿菌菌株E.coliDH5α(北京天根生化科技有限公司),凝膠DNA回收試劑盒(Axygen),SMARTTM RACE cDNA Amplification Kit(Clontech),In-Fusion?HD Cloning Kit(Clontech),其他常用試劑為國產分析純。
1.2 試驗方法
1.2.1材料培養與處理 挑選籽粒飽滿且整齊度一致的紫花苜蓿種子,消毒后用滅菌蒸餾水沖洗2~3遍,播種于鋪有濾紙的培養皿中,置于光照培養箱中培養,待長出兩片子葉后,移至1/2Hoagland營養液中于培養箱中進行培養;培養箱溫度為(28±2)℃/(23±2)℃,光照時間16 h·d-1,光照強度約600 μmol·m-2·s-1,相對濕度60%~80%。培養4周后選取長勢一致的紫花苜蓿幼苗進行脅迫處理,每個處理3次重復。干旱和ABA處理分別用25% PEG6000和100 μmmol·L-1ABA處理0,12,24,48,72 h,冷脅迫(4℃)處理0,4,8,12,24 h后,取其葉片;組織表達特異性分析是分別采取同一株紫花苜蓿上的根、莖、葉和花等組織,-80℃保存備用。
1.2.2總RNA的提取及cDNA第一鏈的合成 參照Trizol總RNA抽提試劑盒說明書(Invitrogen公司)提取1.2.1中樣品的總RNA,經1%瓊脂糖凝膠電泳檢測RNA質量后進行cDNA的合成(cDNA Synthesis Kit,Fermentas公司),產物于-20℃保存備用。
1.2.3引物的設計及合成 根據NCBI上查找到的其它物種的SOS1基因序列進行同源性比對,排除相似性不高的序列,找到高度保守的區域,利用Primer 5.0軟件設計特異性引物MsSOS1-F和MsSOS1-R(表1),送英俊公司合成。
1.2.4紫花苜蓿MsSOS1基因保守區片段的克隆 PCR擴增體系為(10 μl體系):10×Ex Taq Buffer 1 μl,dNTP Mixture 1 μl,cDNA(<500 ng) 0.5 μl,TaKaRa Ex Taq(5 U/μl)0.1 μl,引物MsSOS1-F(10 μm)1 μl,MsSOS1-R(10 μm)1μl,ddH2O 5.4 μl。擴增條件為:94℃ 5 min,94℃ 30 s,56℃ 60 s,30 cycles;72℃ 10 min。擴增產物回收后連接到載體pMD?18-T上,轉化DH5α后挑取陽性克隆測序。
1.2.5紫花苜蓿MsSOS1基因全長克隆 根據克隆得到的紫花苜蓿MsSOS1基因中間片段設計3′-RACE引物MsSOS1-F1和5′-RACE引物MsSOS1-F2(表1),進行全長PCR的擴增,獲得的PCR產物連接載體進行轉化,并挑取陽性克隆送公司測序。

表1 實驗中的引物Table 1 Primers used in the experiment
1.2.6紫花苜蓿MsSOS1基因的生物信息學分析 利用軟件DNAStar進行紫花苜蓿MsSOS1基因ORF的查找和氨基酸翻譯;利用ExPASy工具對MsSOS1蛋白進行理化性質、高級結構和生物學功能的預測分析、信號肽預測、亞細胞定位情況進行分析;利用DNAMAN6.0進行序列的同源性比對分析,MEGA 5.10軟件構建系統進化樹(NJ法)。
1.2.8Real-time PCR檢測紫花苜蓿MsSOS1基因的組織表達特異性 根據紫花苜蓿MsSOS1基因的全長序列設計Real-time PCR引物qRT-MsSOS1-F、qRT-MsSOS1-R(表1),以Actin基因為內參設計特異性引物Actin-F、Actin-R(表1)。使用Bio-Rad CFX96進行Real-time PCR擴增,擴增程序為:94℃ 2min,94℃ 5sec,58℃ 15sec,72℃ 10sec,45個循環,每個樣品3次重復。
利用2-△△Ct方法[46]中的△△Ct=(CtTarget-CtActin)處理-(CtTarget-CtActin)對照公式計算紫花苜蓿根、莖、葉、花中MsSOS1基因的表達量并繪出柱形圖。
2 實驗結果與分析
2.1 紫花苜蓿MsSOS1基因中間片段的獲得
以反轉錄得到的cDNA為模版,PCR擴增得到約784 bp的目的片段(如圖1所示),將PCR結果進行回收、連接、轉化并測序,測序結果進行Blast比對分析,結果顯示獲得的目的片段序列與鷹嘴豆(Cicerarietinum)、大豆(Glycinemax)、花生(Arachisipaensis)、綠豆(Vignaradiata)等基因的同源性達到80%以上,據此推斷該序列是紫花苜蓿MsSOS1基因的一部分,可以進行后續實驗。
2.2 紫花苜蓿MsSOS1基因5′端序列和3′端序列的獲得
根據獲得的目的片段設計RACE引物進行PCR擴增,獲得了5′端片段序列(約1 785 bp)和3′端片段序列(約2 712 bp)(如圖1所示),利用軟件將獲得的5′端序列、3′端序列和中間序列進行拼接,最終獲得紫花苜蓿MsSOS1基因的全長cDNA序列(約3 757 bp)。設計ORF(開放閱讀框)引物進行PCR擴增,獲得約3 000 bp的特異性目的條帶,測序后確定該序列為紫花苜蓿MsSOS1基因ORF序列(約2 580 bp)。
2.3 紫花苜蓿MsSOS1蛋白的生物信息學分析
2.3.1紫花苜蓿MsSOS1基因全長序列及蛋白的理化性質分析 紫花苜蓿MsSOS1基因全長cDNA序列長3 757 bp,包含922 bp的5′非編碼區,2 580 bp的開放閱讀框(ORF)和255 bp的3′非編碼區(圖2),編碼一個分子式為C4337H6816N1156O1258S31、相對分子量為96.3 kDa、理論等電點為6.51的氨基酸序列(859 aa)(表3)。含有堿性氨基酸(K,R)88個;酸性氨基酸(D,E)94個;疏水氨基酸(A,I,L,F,W,V)320個;極性氨基酸(N,C,Q,S,T,Y)218個。該蛋白中Leu(97個,11.3%),Ser(86個,10.0%),Val(59個,6.9%),Glu(58個,6.8%)和Ala(58個,6.8%)相對含量比較多;Asp+Glu為94,Arg+Lys為88;親水性平均數為-0.075,脂肪指數(AI)為95.23,蛋白質三級結構不穩定系數(II)為47.13,表現為不穩定狀態,由此我們可以推測其可能為不穩定的酸性蛋白質。

圖1 紫花苜蓿MsSOS1基因cDNA克隆過程的PCR產物電泳圖譜Fig.1 Electrophoresis in the process of MsSOS1 cDNA cloning注:M為DL5000 DNA Marker;1為RT-PCR產物;2為5′-RACE PCR產物;3為3′-RACE PCR產物;4為MsSOS1基因全長PCR產物Note:M:DL5000 DNA Marker;1:RT-PCR product of MsSOS1 gene;2:5′-RACE PCR product of MsSOS1 gene;3:3′-RACE PCR product of MsSOS1 gene;4:The full-length PCR product of MsSOS1 gene

圖2 紫花苜蓿MsSOS1基因cDNA及其推導氨基酸序列Fig.2 Nucleotide sequence and predicted amino acid sequence of MsSOS1注:左側和右側的數字分別代表核苷酸和氨基酸位點,綠色區域為起始密碼子(ATG)和終止密碼子(TGA),黃色區域代表3′及5′非編碼區,下劃線為預測的跨膜區域Note:The nucleotide sequences and amino acid residues are indicated by numbers of the left and right. The start codon(ATG) and the stop codon(TGA) were green shaded. The red shaded was 3′-UTR and 5′-UTR;The underline was predictive transmembrane region
2.3.2紫花苜蓿MsSOS1蛋白的同源性分析 與其他植物SOS1蛋白的同源性比較結果顯示,紫花苜蓿MsSOS1蛋白與鷹嘴豆(Cicerarietinum)、大豆(Glycinemax)、羽扇豆(Lupinusangustifolius)、花生(Arachisipaensis)、葡萄(Vitisvinifera)等的同源性相對較高(表2),由此可以確定該序列確實是紫花苜蓿SOS1基因,命名為MsSOS1。

表2 紫花苜蓿SOS1蛋白與其它植物SOS1蛋白的同源性分析Table 2 Homology analysis of the MsSOS1 withSOS1 from some higher plants
2.3.3紫花苜蓿MsSOS1蛋白二級結構預測 對紫花苜蓿MsSOS1蛋白二級結構進行預測發現其主要由α-螺旋、β-轉角、無規則卷曲和β-折疊組成(圖3),其中以α-螺旋為主,占46.22%;其次是無規則卷曲,占37.25%;β-轉角含量最少,僅占5.01%。
2.3.4紫花苜蓿MsSOS1蛋白的保守結構域分析 運用生物信息學軟件對MsSOS1蛋白的保守結構域進行分析,結果發現紫花苜蓿MsSOS1蛋白的N-端含有NhaP、Na+/H+Exchanger結合位點和b-cpal、Na+/H+Exchanger superfamily等保守結構域,C-端含有cNMP_binding結合位點和Crp、CAP_ED、CAP_ED superfamily、Crp superfamily等保守結構域,表明該基因應該是Na+/H+轉運蛋白家族中的一員(圖4)。
2.3.5紫花苜蓿MsSOS1蛋白的跨膜結構分析 對紫花苜蓿MsSOS1蛋白的跨膜結構進行分析發現,MsSOS1蛋白的N端和C端均位于細胞外,含有4個跨膜區域,其跨膜區氨基酸序列位置分別為29-51,63-85,100-118,138-160(圖5);對信號肽位點進行分析發現,紫花苜蓿MsSOS1蛋白無信號肽位點存在(圖6)。

圖3 MsSOS1蛋白的二級結構Fig.3 Predicted secondary structure for MsSOS1 protein

圖4 MsSOS1蛋白的功能結構域Fig.4 The conserved domains of the MsSOS1 protein

圖5 MsSOS1蛋白的跨膜區分析Fig.5 Analysis of transmembrane helicesof MsSOS1 protein

圖6 MsSOS1蛋白信號肽預測分析NN-法Fig.6 Analysis and prediction of signal peptideof MsSOS1 based on NN-method
2.3.6紫花苜蓿MsSOS1蛋白的疏水性分析 運用ProtScale程序對紫花苜蓿MsSOS1蛋白的疏水性進行,發現該蛋白中含有疏水區域,N-末端具有4個跨膜區,C-末端有一個很長的親水性尾部,面向細胞膜內腔(圖7),這與先前報道過的擬南芥(Arabidopsisthaliana)、水稻(Oryzasativa)、獐茅(Aeluropuslittoralis)的SOS1蛋白的研究結果相似[26-27,30]。

圖7 MsSOS1蛋白疏水性分析Fig.7 Analysis of hydrophobility of MsSOS1 protein
2.3.7紫花苜蓿MsSOS1蛋白亞細胞定位預測 對紫花苜蓿MsSOS1蛋白進行亞細胞定位結果顯示:denoplasmic reticulum(47.8%) > plasma membrane(26.1%) > nuclear(4.3%) > vesicles of secretorysystem(4.3%) > vacuolar(4.3%) > mitochondrial(4.3%) > cytoplasmic(4.3%) > Golgi(4.3%),由此可以預測紫花苜蓿MsSOS1蛋白可能主要定位于內質網中。
2.3.8紫花苜蓿MsSOS1蛋白的氨基酸序列多重比對及系統進化樹分析 將紫花苜蓿MsSOS1蛋白序列同Genbank上查找的其他已知物種的SOS1蛋白序列進行多重比對發現,MsSOS1蛋白與豆科植物鷹嘴豆(Cicerarietinum)CaSOS1(XM_004504555.2)、大豆(Glycinemax)GmSOS1(NM_001258010.1)和綠豆(Vignaradiata)VrSOS1(KC855193.1)的同源性非常近,在85%以上,分別為91%、87%、85%(圖8)。
利用MEGA5.0軟件對紫花苜蓿MsSOS1蛋白同其他植物的Na+/H+逆向轉運蛋白進行系統進化樹分析發現,紫花苜蓿MsSOS1蛋白與已經報道的具有Na+/H+逆向轉運蛋白運輸功能的幾種植物的同源性很近,而與定位在內膜系統上的NHX類的Na+/H+逆向轉運蛋白的同源性較低(圖9),這些都可以說明MsSOS1可能是質膜型Na+/H+逆向轉運蛋白(SOS1),可能與紫花苜蓿的耐鹽性有著密切的關系。

圖8 紫花苜蓿MsSOS1與其它植物SOS1蛋白的多重比對分析Fig.8 Multiple alignment analysis of MsSOS1 with other plant SOS1 proteins注:多重比對采用DNAMAN軟件進行。Ms:紫花苜蓿Medicago sativa;Ca:鷹嘴豆Cicer arietinum;Ai:花生Arachis ipaensis;Gm:大豆Glycine max;Vr:綠豆Vigna radiata。橫線表示跨膜區Note:The sequence were aligned with DNAMAN software;The transverse line was transmembrane helices
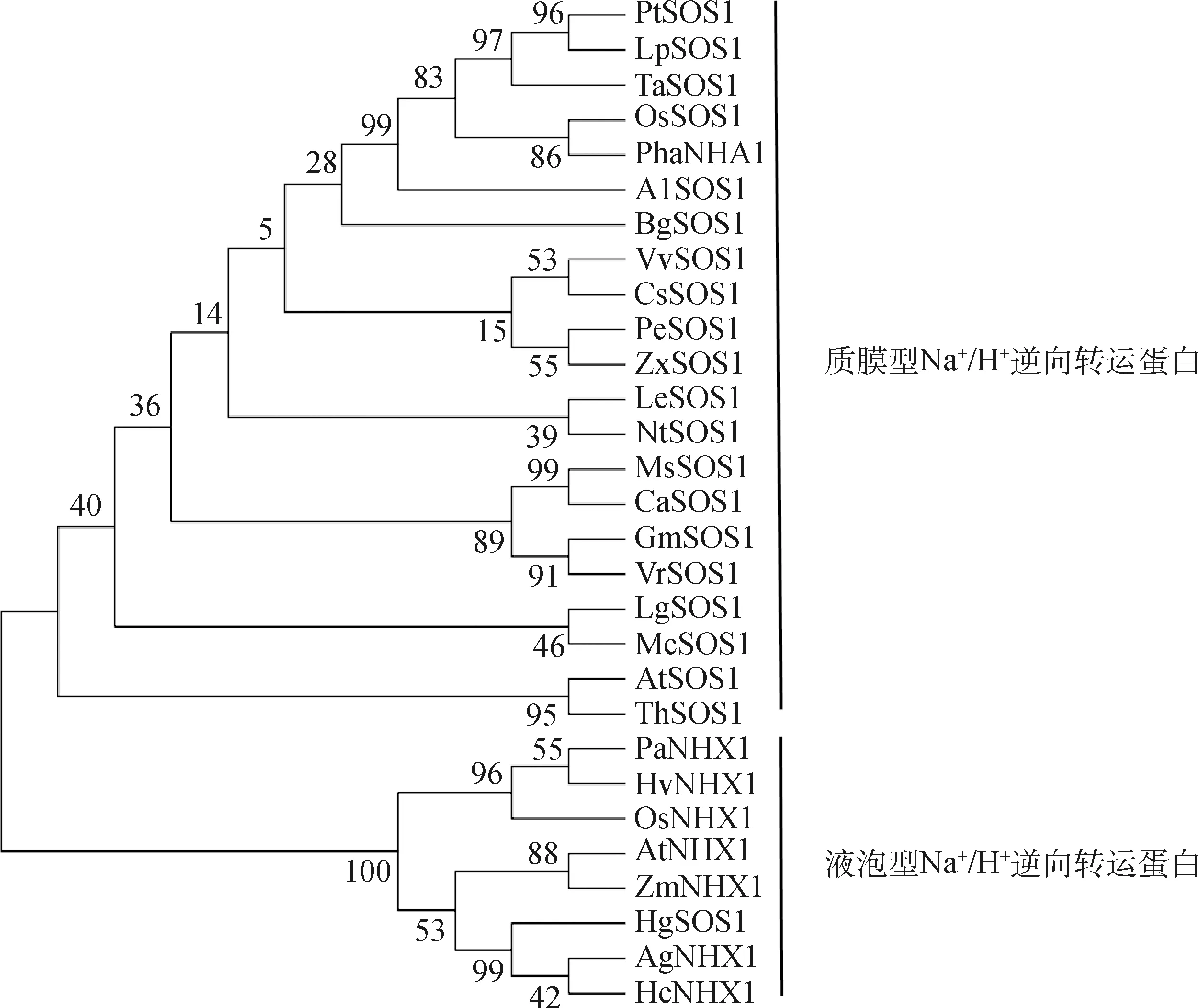
圖9 紫花苜蓿MsSOS1與其他植物Na+/H+逆向轉運蛋白的系統進化樹分析Fig.9 Phylogenetic analysis of MsSOS1 in Medicago sativa with closely related Na+/H+antiporters from different plant species
2.4 紫花苜蓿MsSOS1基因的組織特異性表達分析
對MsSOS1基因在同一株紫花苜蓿不同組織中的相對表達量進行分析并作圖(圖10),以花中MsSOS1基因的表達量為對照發現,在正常生長條件下,MsSOS1基因在紫花苜蓿各組織中均有表達,在根中的表達量最高,且明顯高于其它組織,是對照的11.9倍,葉中的表達量次之,是對照的6.7倍,莖中的表達量不是很明顯,但也達到了對照的3.4倍(圖10)。由此結果可以看出,MsSOS1基因在紫花苜蓿的各個器官中均有表達,只是表達峰度不同。

圖10 MsSOS1基因的組織表達特異性Fig.10 Organ-specific expression pattern of MsSOS1
2.5 不同非生物脅迫下紫花苜蓿MsSOS1基因的表達分析
為了研究紫花苜蓿MsSOS1基因是否受非生物脅迫的誘導,分別對紫花苜蓿幼苗進行了不同的非生物脅迫處理,分析不同組織中MsSOS1基因的表達情況。
2.5.14℃脅迫下MsSOS1基因在紫花苜蓿中的表達量分析 4℃脅迫條件下,以0 h為對照分析MsSOS1基因在不同時間段的表達差異。結果發現,該基因受4℃脅迫誘導比較明顯。葉片中MsSOS1基因的表達量隨著脅迫時間的增加呈現出先升高后降低的趨勢,且于8 h達到最大值;莖和根中的表達量隨著處理時間的增加均呈現先降低后增高的趨勢,但均低于對照(圖11)。
2.5.2PEG6000脅迫下MsSOS1基因在紫花苜蓿中的表達量分析 25% PEG6000模擬干旱條件下,以0 h為對照分析MsSOS1基因在不同時間段的表達差異。結果發現,該基因受PEG6000誘導表達比較明顯,在根、莖和葉中均有不同程度的表達(圖12)。在葉中,隨著PEG誘導時間的變化,MsSOS1基因的表達呈逐漸上升的趨勢,于72 h表達量顯著增加且達到最大表達量,是對照的7.5倍;隨著PEG脅迫時間的延長,MsSOS1基因在根和莖中的表達量基本上都是出現先升高后降低的趨勢,根中的表達量于12 h達到最大值,是對照的3.67倍;莖中的表達量于24 h達到最大值,是對照的8.23倍。

圖11 MsSOS1基因在4℃脅迫下的表達模式分析Fig.11 Expression pattern of MsSOS1 in response to 4℃ stress

圖12 MsSOS1基因在PEG脅迫下的表達模式Fig.12 Expression pattern of MsSOS1 in response to PEG stress
2.5.3脫落酸(ABA)脅迫下MsSOS1基因在紫花苜蓿中的表達量分析 以0 h為對照分析MsSOS1基因在ABA脅迫下的不同時間段的表達差異。結果發現,在ABA誘導下,隨著時間的延長,MsSOS1基因主要在莖和葉中表達(圖13),在葉中,MsSOS1基因的表達量在12 h達到最低值,24 h達到最大值,是對照的3.14倍;在莖中,隨著處理時間的延長,MsSOS1基因的表達量呈現先升高后降低的趨勢,且于24 h達到最大值,是對照的9倍;ABA處理對MsSOS1基因在根部的表達影響不顯著,隨著脅迫時間的延長,表達量幾乎不變。
3 討論
目前,很多研究已經確定了從藍細菌到真菌、從藻類到高等植物中耐鹽性的決定因素。在正常條件下,植物體細胞質中維持比較高的K+/Na+,而當它受到鹽脅迫時,植物就會啟動體內多種適應機制如傳感、信號轉導、基因表達和代謝途徑等來適應這些不利環境。為了了解這些機制中有哪些部分可以提高植物對鹽脅迫的適應性,我們重點研究了紫花苜蓿中與鹽度耐受性相關的主要基因的分離、鑒定和表達模式分析。在本研究中,我們克隆得到了紫花苜蓿MsSOS1基因的全長序列并對其進行了序列和表達分析。紫花苜蓿MsSOS1蛋白N-端具有4個高度同源的、具有水溶性特征的、主要負責Na+轉運的功能結構域,這與前人所報道的一般具有10~12個跨膜結構域有所不同,這可能是由于物種之間的差異所造成的;C-端是可以確定其確實是屬于參與細胞內Na+調控轉運的Na+/H+逆向轉運蛋白家族成員的高度親水的調控結構域,末端還有一個很長的、面向細胞質內腔的親水性尾部,推測它可能是通過與逆境相關的調控因子發生互作來提高植物的抗性;另外,C-末端還存在1個SOS1蛋白的活性調節區(即磷酸化調節位點),這與擬南芥(Arabidopsisthaliana)、水稻(Oryzasativa)和獐茅(Aeluropuslittoralis)等的SOS1蛋白結構相似[26-27,31],只是不同的物種之間存在一些差異。

圖13 MsSOS1基因在ABA處理下的表達模式Fig.13 Expression pattern of MsSOS1 in response to ABA treatment
對編碼后的MsSOS1蛋白序列進行分析發現,該蛋白與已經報道的高等植物的SOS1基因蛋白序列相似度比較高;通過比對分析發現該蛋白與豆科植物鷹嘴豆(Cicerarietinum)、大豆(Glycinemax)等豆科植物的SOS1蛋白的同源性比較高,與其它科屬植物的同源性比較低,說明SOS1蛋白在不同的物種之間雖然具有較高的保守性,但也存在一定的差異;但是,將其與模式植物擬南芥相比,同源性相對較低,說明這兩種蛋白之間的差異較大,由此可以推測它們在結構和功能上也可能會存在不同的差異[32],這也就需要我們通過進一步的功能表達分析研究該基因在耐鹽性狀方面的特點來進行鑒別。另外,已有研究報道SOS1基因在植物的耐鹽方面起著至關重要的作用,同時研究發現SOS1基因超表達可以使植物的耐鹽性增強[47],這也預示著紫花苜蓿的MsSOS1基因在植物的耐鹽方面可能也起著重要的作用。
采用實時熒光定量的方法對紫花苜蓿MsSOS1基因進行組織特異性表達分析,結果發現該基因在紫花苜蓿生育期的根、莖、葉、花中均有表達,這與擬南芥AtSOS1[21]和菊苣CiSOS1[48]在根和地上部分均有表達的結果相一致。從圖11中也可以看到,紫花苜蓿MsSOS1基因在生育期的表達峰值為:根>葉>莖>花,具有根部表達的特性,它主要表現為根部組織通過對紫花苜蓿MsSOS1基因的表達進行上調的方法,將根系中多余的Na+排除體外,降低根中Na+濃度來抵御Na+對植物體本身的毒害作用。對紫花苜蓿MsSOS1基因在不同非生物脅迫下的表達模式研究發現,不同的非生物脅迫表達模式有所區別,Munns等[49]研究認為植物激素ABA是植物適應鹽脅迫過程最初的信號物質,在植物應答鹽脅迫的過程中具有重要的作用。已有文獻報道,ABA處理植株能增強其在鹽脅迫條件下的適應能力[50],Shi等研究發現擬南芥AtSOS1基因在ABA處理下表達量沒有影響,而在本研究中,MsSOS1基因在根中的表達影響不明顯,主要在莖和葉中表達,莖中的表達相對比較明顯,推測該基因可能具有在莖維管束薄壁細胞表達的特異性[51];本研究中,MsSOS1基因還受到PEG脅迫的強烈誘導,無論是在根還是在莖和葉中均有不同程度的表達,與Song等對大島野路菊(Chrysanthemumcrassum)CcSOS1基因的研究相一致[52],說明該基因也可能參與了紫花苜蓿的抗旱過程。
4 結論
本研究從豆科植物中苜1號紫花苜蓿中克隆得到一個MsSOS1基因,對其進行序列分析發現,該基因包含一個2 580 bp的開放閱讀框,編碼859個氨基酸;蛋白序列與豆科植物鷹嘴豆的氨基酸序列的同源性最高,為91%,進化樹分析發現該基因屬于質膜型Na+/H+逆向轉運蛋白。另外,利用qRT-PCR技術對其進行表達分析發現,該基因的表達具有組織表達特異性,且在根中的表達量最大,其次是莖和葉;4℃、PEG和ABA等非生物脅迫都會誘導MsSOS1基因的表達,只是表達峰度不同,推測該基因可能在紫花苜蓿的抗逆分子機制中發揮一定的作用。這對于了解豆科植物Na+/H+逆向轉運蛋白基因參與植物抗逆性的分子機制有了一定的了解,并為深入解析紫花苜蓿MsSOS1基因的功能和培育抗逆品種提供了良好的理論基礎。
猜你喜歡
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
電子制作(2018年18期)2018-11-14 01:48:24
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
山東工業技術(2016年15期)2016-12-01 05:31:22
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
