miR-141及MMP-16對骨性關節炎疼痛及軟骨損傷的影響
2018-03-03 02:54:31余文雄
現代中西醫結合雜志 2018年4期
關鍵詞:手術
胡 玲,馬 垚,胡 冰,余文雄,左 毅
(武漢科技大學附屬天佑醫院,湖北 武漢 430064)
骨性關節炎(OA)是一種以關節軟骨變性、破壞及骨質增生為特征的慢性疼痛性疾病。我國OA患病率約為15%,同時伴隨年齡的增長,其患病率不斷升高,最終導致殘疾[1-2]。近年來的研究表明,微小RNA(miRNA)具有調節軟骨發育及維持軟骨代謝平衡的作用,通過對其下游多靶點基因的干預,可調節軟骨細胞分化、生長和修復,從而影響OA的病變過程[3]。miR-141是OA患者血液中具有高度特異性的miRNAs,其降低與關節軟骨細胞凋亡和炎癥相關,可影響OA的進展;同時其可通過對炎性因子及神經細胞中致痛因子的調控,從而對OA產生的疼痛具有調節作用[4]。miR-141家族在OA患者軟骨組織中的表達明顯高于正常組織[5],生物信息學分析發現其能被特定的基質金屬蛋白酶-16(MMP-16)所調控而影響骨關節炎的進展。因此如何在早期阻斷關節軟骨的損傷,減輕或消除疼痛,改善關節功能,降低致殘率是OA研究領域中的重要課題。本實驗擬通過觀察OA模型大鼠背角神經元中miR-141及軟骨組織中MMP-16的表達情況來探究它們的相關性及功能意義,以期為OA嚴重程度的判斷及靶向治療提供新思路和理論依據。
1 實驗資料
1.1 動物 健康雄性SD大鼠30只,體質量250~300 g,武漢大學實驗動物中心提供。分籠進行飼養,自由飲食。環境溫度20~26 ℃,濕度50%~70%,晝夜節律以1∶1進行調節。
1.2 主要試劑和儀器 兔抗大鼠MMP-16抗體(博士德BA1280),兔二步法檢測試劑盒(中杉金橋PV-6001);miScript Ⅱ反轉錄試劑盒(QIAGEN-218161)、miScript SYBR Green PCR 檢測試劑盒(QIAGEN-218073)、1% 瓊脂糖、漩渦振蕩機(江蘇海門其林貝爾儀器制造有限公司);Heraguard EC 超凈工作臺( 美國 Thermo Scientific公司) ;單道可調移液器(德國 Eppendorf 公司);CFX96 熒光定量PCR儀(美國 Bio-Rad 公司);Power Pac Basic 電泳儀(美國 Bio-Rad 公司);水平電泳槽(美國 Bio-Rad 公司)。
1.3 分組及造模 將SD大鼠編號后,隨機分為實驗組20只和對照組10只。采用苯巴比妥麻醉大鼠,仰臥位固定,以左側膝關節作為手術側,常規備皮消毒。髕旁內側切口顯露膝關節,暴露髕骨后向外側脫位,盡量屈曲膝關節顯露前交叉韌帶,實驗組直視下切斷前交叉韌帶,抽屜試驗確認完全切斷后徹底止血,將髕骨復位,1號絲線逐層閉合關節腔。對照組不切斷前交叉韌帶,其余處理同實驗組。術后大鼠手術側給予固定,分籠飼養,每只大鼠肌注青霉素20萬IU/d預防感染,連續術后3 d。
1.4 冷痛閾測定 分別在手術前連續3 d以及手術2,8周后,于每天上午的固定時間點進行測量。測量前將大鼠靜置入多孔鋼絲籠中,適應環境15 min后,在右側皮區(右側為優勢區,更加敏感)懸滴0.5 mL冷丙酮,記錄滴下冷丙酮后1 min內后爪抓撓皮區或理毛的次數,重復測量3次,每次間隔5 min,取3次記錄值的平均值作為冷痛閾值。
1.5 軟骨組織病理切片觀察 2組分別于手術2,8周后各斷頸處死一半大鼠,取2 cm×0.5 cm軟骨組織,經生理鹽水沖洗后,即行多聚甲醛固定、脫鈣、沖洗、脫水、透明、石蠟包埋、切片,通過蘇木精-伊紅染色觀察大鼠骨關節的形態變化。
1.6 背根神經節組織中miR-141表達檢測 采用RT-PCR法檢測。斷頸處死大鼠后取L4—6脊髓組織,分離出雙側脊髓背角組織,提取總RNA,在熒光定量PCR儀上按照標準步驟進行反應。按照反應體系配置說明配備總反應液25 μL:其中2×Quantiect SYBR Green PCR 12.5 μL,10×miscript Universal Primer 2.5 μL,10×miscript Primer Assay 141 2.5 μL,RNase-free water 5 μL,Template CDNA 2.5 μL,每個樣本設3個復孔,反應條件為:95 ℃預變5 min,1個循環。15 s 94 ℃,30 s 55 ℃,30 s 70 ℃,40個循環。計算2-ΔΔCT值進行定量分析,2-ΔΔCT值表示目的基因相對表達量。
1.7 MMP-16蛋白表達檢測 采用免疫組化法檢測。取4 μm厚的軟骨組織切片,常規脫蠟至水,0.3% H2O2孵育10 min阻斷內源性過氧化物酶。0.01 mol/L PBS沖洗5 min×3次,將切片浸入0.01 mol/L檸檬酸鹽抗原修復液(pH=6.0),高壓2 min抗原修復,冷卻至室溫。0.01 mol/L PBS沖洗5 min×3次,滴加一抗(MMP-16濃度為1∶200),4 ℃冰箱孵育過夜。0.01 mol/L PBS沖洗5 min×3次,滴加生物素標記的二抗,室溫孵育30 min。0.01 mol/L PBS沖洗5 min×3次,DAB顯色,蘇木素復染,樹膠封片。PBS代替一抗作為陰性對照。鏡下陽性細胞核呈深淺不一的棕色,隨機10個視野下計數陽性著色細胞。
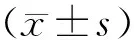
2 結 果
2.1 手術前后大鼠冷痛閾值比較 手術2,8周后,對照組大鼠對冷丙酮刺激的陽性反應次數未見明顯變化(P均>0.05);實驗組大鼠對丙酮刺激的陽性反應次數明顯高于對照組(P<0.05),并且隨著時間的延長,陽性反應次數明顯增加(P<0.05)。見表1。
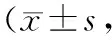
表1 2組大鼠手術前后冷痛閾值比較次)
注:①與手術前1 d比較,P<0.05;②與手術后第2周比較,P<0.05;③與對照組比較,P<0.05。
2.2 大鼠滑膜軟骨組織HE染色表現 對照組大鼠假手術2周和8周后HE染色表現無明顯變化,軟骨細胞形態扁平,排列整齊,結構清晰,無炎性細胞浸潤,無充血增生,間質內無纖維化。實驗組大鼠手術2周后軟骨細胞發生收縮,數量增多、排列紊亂,基質染色減退,此時發生以巨噬細胞、單核細胞、嗜酸性粒細胞為主的急性炎性細胞浸潤,可見明顯的增生血管和輕度纖維化;手術8周后,大鼠軟骨細胞數量明顯減少,形態消失,可見成團積聚的炎癥細胞出現,同時組織裂隙較深,基質不可染色,出現關節結構的改變。見圖1~3。

圖1 假手術2周后對照組大鼠滑膜軟骨組織HE染色表現 (×100)
2.3 大鼠背根神經節組織中miR-141相對表達量 手術2,8周后,實驗組大鼠背根神經節組織中miR-141相對表達量均明顯高于對照組(P均<0.05),且手術8周后明顯高于手術2周后(P<0.05)。見表2。

圖2 手術2周后實驗組大鼠滑膜軟骨組織HE染色表現(×100)

圖3 手術8周后實驗組大鼠滑膜軟骨組織HE染色表現(×100)

組別n手術2周后手術8周后對照組513.43±2.3414.54±3.82實驗組1028.54±62.72①34.56±7.98①②
注:①與對照組比較,P<0.05;②與手術2周后比較,P<0.05。
2.4 大鼠軟骨組織中MMP-16蛋白表達情況 對照組大鼠假手術2,8周后骨組織中MMP-16蛋白表達量比較差異無統計學意義(P均>0.05);實驗組大鼠手術2,8周后骨組織中MMP-16蛋白表達量均明顯高于對照組(P均<0.05),且手術8周后明顯高于手術2周后(P<0.05)。見圖4~6。

圖4 假手術2周后對照組大鼠軟骨組織中MMP-16蛋白表達情況(×100)
3 討 論
OA作為一種累及關節軟骨的慢性退行性病變,其病因尚不清楚,患者主要表現為關節疼痛及腫脹,這與軟骨及其微環境的合成代謝障礙所致的胞外基質(ECM)退化和關節軟骨的損傷有關,隨著軟骨的破壞、外界機械因素刺激以及游離物質的析出,導致疼痛加劇及關節活動受限,最終出現關節畸形[6-7]。miRNAs是一類長度為22個小核苷酸,來源于內源染色體上的非編碼單鏈RNA,它們在進化上具有高度的保守性,能夠通過與靶mRNA特異性的堿基互補配對,引起靶mRNA 降解或抑制其翻譯,從而調控基因進行轉錄后的表達。參與細胞增殖凋亡、器官形成、炎癥免疫應答、炎性疼痛、腫瘤發生等進程[8],近年來研究表明miRNAs的表達差異與OA發生、發展過程中關節軟骨破壞、軟骨細胞凋亡、滑膜炎癥以及疼痛等病理變化關系密切[9]。miR-141是一種在軟骨組織中表達的miRNAs。張文明[5]發現在成骨過程中,miR-141是成骨負性調節因子,它通過轉錄后抑制降低Dlx5/Msx2/Runx2信號軸的級聯作用,減少成骨表達和成骨作用。申發娟等[10]研究發現miR-141高表達的胃腺癌SGC-901細胞E-cadherin表達增加,N-cadherin、MMP-9表達減少,細胞遷移減少。這些都提示miR-141與骨細胞代謝生長、腫瘤、炎癥有密切關系。本實驗發現,實驗組手術2,8周后背根神經節組織中miR-141表達量均明顯高于對照組,且手術8周后高于手術2周后,表明隨著OA的發展,miR-141呈現持續上升的趨勢。另外手術2周和8周后實驗組大鼠對丙酮刺激的陽性反應次數明顯高于對照組,且隨著病情進展其陽性反應次數明顯增加。推測脊髓背根神經元中miR-141升高可以導致OA疼痛的發生,并可反映嚴重程度。

圖5 手術2周后實驗組大鼠軟骨組織中MMP-16蛋白表達情況(×100)

圖6 手術8周后實驗組大鼠軟骨組織中MMP-16蛋白表達情況(×100)
隨著分子生物學的快速發展,MMP-16在OA發病中的作用備受關注。Nobuaki等[11]發現IL-1β通過Wnt16信號通路誘導MMP-16產生并促進成骨細胞的增殖,但在OA患者中發現MMP-16不僅可以降解ECM,而且也可以激活MMP-2在酶原水平上調節細胞外基質降解,這都促進了OA的發展[12]。Zhang等[13]證實下調miR-155可以導致MMP-16的基因表達增加,降解ECM,從而引起椎間盤疾患,而上調MMP-16的表達可達到治療腰椎間盤突出的作用。本實驗發現,實驗組大鼠手術2,8周后骨組織中MMP-16蛋白表達量均明顯高于對照組,且手術8周后明顯高于手術2周。這表明MMP-16在OA的病理過程中起到降解軟骨細胞,促進病情發展的作用。
綜上所述,在OA的發展過程中,脊髓背角細胞中的miR-141可導致關節疼痛,而MMP-16可以競爭性地降解軟骨細胞ECM,促進OA的發展。兩者相互作用促進病情的發展,最終可能導致OA晚期出現關節疼痛和畸形,但兩者之間如何互相調節還有待于進一步研究。
[1] Min Z,Zhang R,Yao J,et al. MicroRNAs associated with osteoarthritis differently expressed in bone matrix gelatin (BMG) rat model[J]. Int J Clin Exp Med,2015,8(1):1009-1017
[2] Murata K,Furu M,Yoshitomi H,et al. Comprehensive microRNA analysis identifies miR-24 and miR-125a as plasma biomarkers for rheumatoid arthritis[J]. Plos One,2013,8(7): 69118-69120
[3] Alcaraz MJ,Megías J,García-Arnandis I, et al. New molecular targets for the treatment of osteoarthritis[J]. Biochem Pharmacol,2010,80(1):13-21
[4] Shen WS,Xu XQ,et al. Potential mechanisms of microRNA-141-3p to alleviate chronic inflammatory pain by downregulation of downstream target gene HMGB1: in vitro and in vivo studies[J]. Gene Therapy,2017,24(6):353-360
[5] 張文明. miR-141調控OP大鼠BMSCs成骨分化機制及健骨顆粒干預作用研究[D]. 福州:福建中醫藥大學, 2016
[6] Benito MJ,Veale DJ,FitzGerald O,et al. Synovial tissue inflammation in early and late osteoarthritis[J]. Ann Rheum Dis,2005,64(9):1263-1267
[7]ZeaerstromC,JiangW,WahamaaH,etal.Pivotaladvance:inhibitionofHMGB1nucleartranslocationasamechanismfortheanti-rheumaticeffectsofgoldsodiumthiomalate[J].LeukocBiol,2008,83(1):31-38
[8]StadlerBM,Ruohola-BakerH.SmallRNAs:keepingstemcellsinline[J].Cell,2008,132(4):563-566
[9]ChenB,HuangI.MicroRNAs’involvementinosteoarthritisandtheprospectsfortreatments[J].EvidBasedComplementAlternatMed,2015,10(26):236179
[10] 申發娟,蘇娟,張慶瑜,等. 上調miR-200a、miR-141表達對胃腺癌細胞生長的體外研究[J]. 天津醫藥,2013,41(6):517-519
[11]NobuakiOzeki,MakioMogi,NaokoHase,etal.Wnt16SignalingisrequiredforIL-1β-inducedmatrixmetalloproteinase-16-regulatedproliferationofhumanstemcell-derivedosteoblasticcells[J].IntJMolSci,2016,17(2):221
[12]JungJC,WangPX,ZhangG,etal.Collagenfibrilgrowthduringchickentendondevelopment:matrixmetalloproteinase-2anditsactivation[J].CellTissueRes,2009,336(1):79-89
[13]ZhangWL,ChenYF.RoleofmiR-155intheregulationofMMP-16expressioninintervertebraldiscdegeneration[J].JOrthopRes,2017,35(6):1323-1334
猜你喜歡
環球時報(2022-12-23)2022-12-23 09:28:37
昆明醫科大學學報(2022年1期)2022-02-28 07:45:04
中老年保健(2021年11期)2021-08-22 03:13:36
昆明醫科大學學報(2021年2期)2021-03-29 07:42:46
河北畫報(2020年10期)2020-11-26 07:20:50
小學閱讀指南·低年級版(2017年1期)2017-03-13 20:07:35
中國衛生標準管理(2015年3期)2016-01-14 03:41:47
中國醫療美容(2015年1期)2015-07-12 10:06:38
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:54
西南軍醫(2014年5期)2014-04-25 07:42:48
