母豬繁殖性能相關5個候選基因的PCR-RFLP多態性研究
2018-03-07 10:25:02吳華莉涂尾龍曹建國都啟晶羊兆洪談永松
上海農業學報 2018年1期
吳華莉,涂尾龍,曹建國,3,都啟晶,常 華,羊兆洪 ,談永松*
(1上海市農業科學院畜牧獸醫研究所,上海201106;2上海市調控生物學重點實驗室,上海200241;3上海種豬工程技術研究中心,上海201302;4青島農業大學食品科學與工程學院,青島266109;5云南農業大學動物科學技術學,昆明650201;6淮安洪泰良種豬繁育有限公司,淮安223472)
豬總產仔數、產活仔數和出生窩重是母豬重要的繁殖性狀,與眾多調控基因有關。目前學者們針對豬繁殖性狀相關基因開展研究,如雌激素受體(Oestrogen receptor,ESR)、含凝血酶敏感蛋白模體的去解聯金屬蛋白酶1基因(A disintegrin and metalloproteinase with thrombospondin motifs-1,ADAMTS1)、巖藻糖轉移酶1基因(α1,2-fucosyl transferase,FUT1)、核受體輔激活蛋白 1(Nuclear receptor co-activator 1,NCOA1)和視黃醇結合蛋白4基因(Retinol-binding proteins 4,RBP4)等[1]。ESR通過與雌激素的作用促進細胞合成,參與陰道上皮細胞和子宮內膜細胞分化[2]。ADAMTS1在豬卵丘-卵母細胞復合體中表達,體外培養的作用是促進卵母細胞成熟[3]。NCOA1是類固醇受體的輔助激活劑家族成員,NCOA1和NCOA3共同作用調控胎盤的形態變化和胚胎存活[4]。FUT1基因M307位存在A/G突變,導致丙氨酸突變蘇氨酸,從而顯著減少FUT1酶的活性,引起GG和AG型個體對產腸毒素大腸桿菌(ETEC)F18敏感,而AA基因型則表現為抗性[5]。有研究報道證實FUT1基因AA型斷奶仔豬成活率高于BB型的2倍,可作為提高斷奶仔豬成活率的分子標記[6-7]。RBP4基因在囊胚擴張時期的表達量增加,并且在懷孕10—12 d的母豬子宮內膜也呈現高表達[8-9]。此外,RBP4基因也被發現是孕體植入前的主要分泌產物[10]。
本研究采用PCR-RFLP方法檢測ESR、NCOA1、ADAMTS1、FUT1和RBP4基因在杜洛克、長白和大白群體中的基因型頻率和等位基因頻率,并分析它們與豬總產仔數(Total number born,TNB)、產活仔數(Number born alive,NBA)和出生窩重(Birth litter weight,BLW)的相關性。
1 材料與方法
1.1 試驗用豬及樣品
來自上海祥欣畜禽有限公司有杜洛克(100頭)、大白(150頭)和長白(80頭),10月齡母豬,用于本試驗5個基因的多態性分析。采集的豬耳組織塊置于75%酒精中帶回實驗室。采用AXYGEN試劑盒(上海鼎盛生物科技有限公司)提取DNA并測定濃度,檢測合格的樣品放入-20℃冰箱保存備用。
收集生產記錄包括耳號、TNB、NBA和BLW。針對每個樣品對應5個基因的不同基因型和生產性能記錄進行數據整理、比對和估計最小二乘均值等處理。
1.2 PCR-RFLP分析
使用特異性引物擴增5個基因的PCR產物分別進行瓊脂糖檢測,要求目的條帶清晰明亮且無雜帶,方可進行相應酶切試驗。5個基因PCR擴增引物序列,染色體位置,退火溫度和對應內切酶見表1。酶切體系(20μL):6μL PCR產物、5 U限制性內切酶0.5μL、Buffer 1μL和ddH2O 12.5μL。限制性內切酶購于Fermentas和寶生物工程有限公司。
1.3 統計方法
計算出每個SNP基因頻率和等位基因頻率,雜合度的計算公式:H=1-∑pi。
1.4 PCR產物酶切與分型
使用1.5%瓊脂糖電泳檢測5個基因PCR擴增產物。使用2.5%瓊脂糖電泳檢測酶切后產物。不同基因型分別選取3個樣品進行測序。根據電泳圖譜和測序結果進行基因型分型。5個基因不同基因型判定標準見表2。

表1 5個基因PCR擴增引物序列、染色體位置、退火溫度和對應內切酶Table 1 The primer sequences,chromosomal locations,temperatures,and endonucleases of five genes

表2 5個基因PCR產物酶切與帶型Table2 Restriction digestion patterns of five gene PCR products bp
1.5 統計分析
計算5個基因在3個豬群體內的基因型頻率和基因頻率。采用SPSS軟件進行5個基因不同基因型TNB、NBA和BLW差異比較分析。Yijk=μ+gk+eijk,yijk表示繁殖性狀(TNB、NBA和BLW),μ為群體均值,gk為基因效應,eijk為隨機殘差。
2 結果與分析
2.1 5個基因在3個豬群體內的基因型頻率與基因頻率
5個基因在3個豬群體內的基因型頻率和等位基因頻率見表3。

表3 5個基因在3個豬群體中的基因型頻率與等位基因頻率Table 3 Genotype and allele frequencies of five genes in three pig groups
ESR基因在杜洛克群體內只檢測出AA型,在長白群體內,檢測出AA型(0.94)和AB型(0.06);在大白群體內檢測出AA型(0.54)、BB型(0.03)和AB型(0.43);在長白和大白群體內A等位基因頻率均高于B等位基因頻率。NCOA1基因在杜洛克群體內檢測出AA型(0.59)、BB型(0.05)和AB型(0.36);在大白群體內檢測出AA型(0.78)、BB型(0.05)和AB型(0.17);在長白群體內只檢測出AA型;在3個豬群體內A等位基因頻率均高于B等位基因頻率。ADAMTS1基因在杜洛克群體內檢測出AA型(0.37)、BB型(0.13)和 AB型(0.50);在長白群體內檢測出 AA型(0.14)、BB型(0.40)和 AB型(0.46);在大白群體內檢測出AA型(0.10)、BB型(0.46)和AB型(0.44);除了在杜洛克群體外,B等位基因頻率均高于A等位基因頻率。FUT1基因在杜洛克群體內檢測出AA型(0.16)、BB型(0.44)和AB型(0.40);在長白群體內檢測出BB型(0.79)和AB型(0.21);在大白群體內檢測出AA型(0.03)、BB型(0.57)和AB型(0.40);在3個豬群體內B等位基因頻率均高于A等位基因頻率。RBP4基因在杜洛克群體內檢測出AA型(0.68)、BB型(0.14)和AB型(0.18);在長白群體內檢測出 AA型(0.41)、BB型(0.21)和 AB型(0.38);在大白群體內檢測出AA型(0.25)、BB型(0.47)和AB型(0.28);除了大白豬群體外,A等位基因頻率均高于B等位基因頻率。表3中各基因的基因型頻率經χ2檢驗均不顯著,符合哈代-溫伯格平衡定律。
2.2 5個基因不同基因型TNB、NBA和BLW在3個豬群體內差異比較
5個基因不同基因型在3個豬群體的TNB、NBA和BLW差異比較分析見表4、表5和表6。
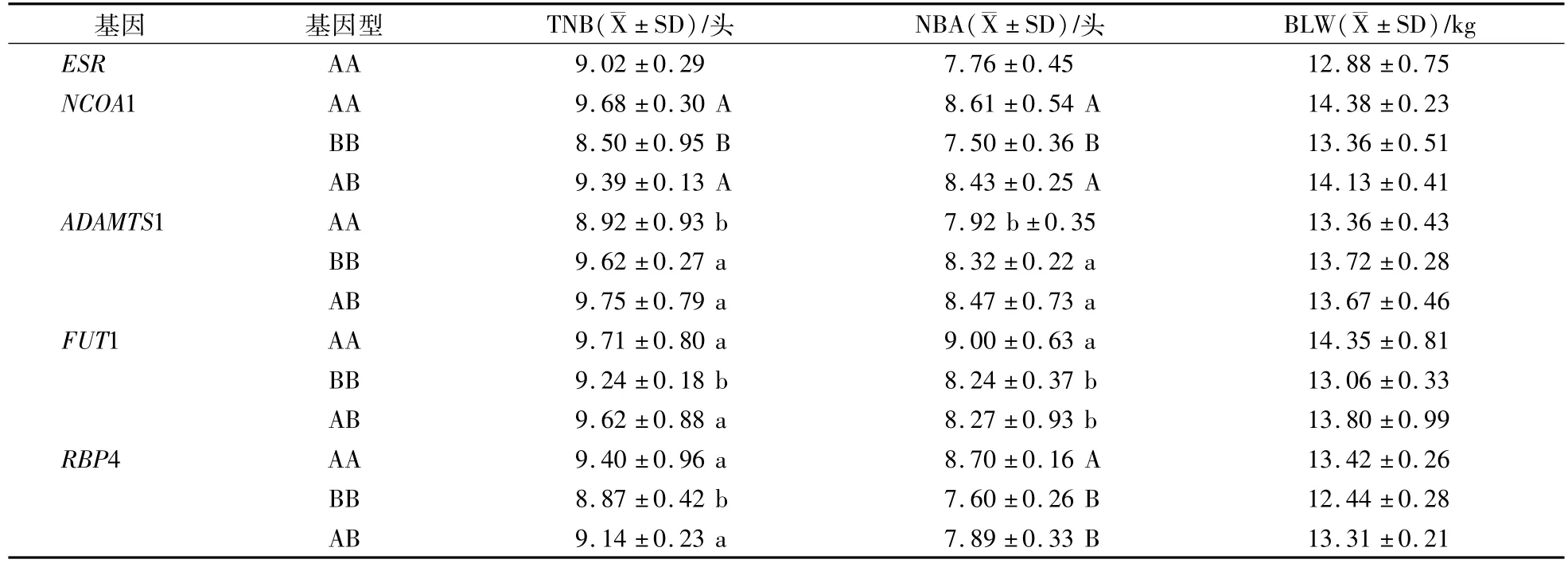
表4 5個基因對杜洛克TNB、NBA和BLW影響Table 4 The influence of five genes to Duroc TNB,NBA and BLW
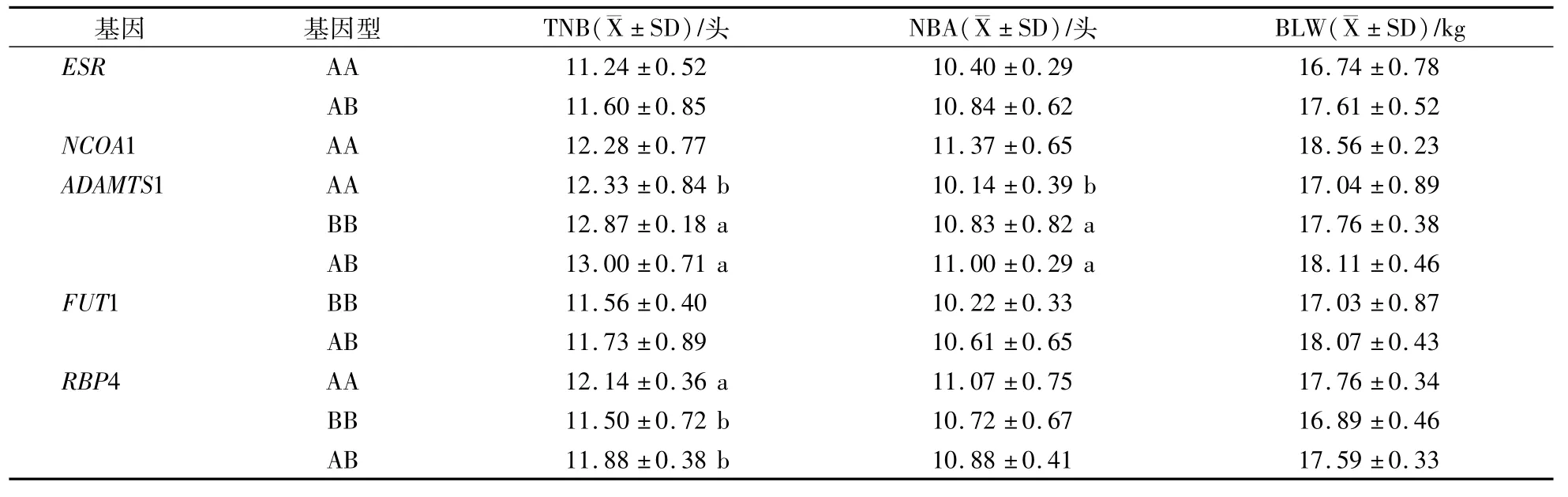
表5 5個基因對長白TNB、NBA和BLW影響Table 5 The influence of five genes to landrace TNB,NBA and BLW
在長白群體內ESR基因AB型的TNB、NBA、BLW比AA型的分別高0.36頭、0.44頭和87 g(P>0.05);在大白群體內BB型的TNB、NBA、BLW比AA型的分別高0.36頭、0.41頭和98 g(P>0.05)。在大白群體內BB型的TNB、NBA和BLW最高;在杜洛克群體內只檢測出AA型。
NCOA1基因在杜洛克和大白群體內TNB、NBA和BLW都表現AA型>AB型>BB型的趨勢。在杜洛克群體內AA型的3個指標比BB型的分別高1.18(P<0.01)、1.11頭(P<0.01)和102 g(P>0.05);在大白群體內AA型的TNB、NBA、BLW比BB型的分別高0.53頭(P<0.05)、1.92頭(P<0.01)和102 g(P>0.05);在長白群體內只檢測出AA型。
ADAMTS1基因在杜洛克、長白和大白群體內TNB、NBA和BLW都表現AB型>BB型>AA型的趨勢。在杜洛克群體內AB型的TNB、NBA、BLW比AA型的分別高0.83頭(P<0.05)、0.55頭(P<0.05)和31 g(P>0.05);在長白群體內AB型的3個指標比AA型的分別高0.67頭(P<0.05)、0.86頭(P<0.05)和107 g(P>0.05);在大白群體內AB型的3個指標比AA型的分別高0.88頭(P<0.05)、0.54頭(P<0.05)和56 g(P>0.05)。
FUT1基因在杜洛克和大白群體內TNB、NBA和BLW都表現AA型>AB型>BB型趨勢。在杜洛克群體內AA型的TNB、NBA、BLW比BB型的分別高0.47頭(P>0.05)、0.76頭(P<0.05)和89 g(P>0.05);在大白群體內AA型的3個指標比BB型的分別高0.56(P<0.05)、1.06頭(P<0.01)和108 g(P>0.05);在長白群體內AB型的3個指標比BB型的分別高0.17頭、0.39頭和104 g(P>0.05)。
RBP4基因在杜洛克、長白和大白群體內TNB、NBA和BLW都表現AA型>AB型>型BB趨勢。在杜洛克群體內AA型TNB、NBA、BLW比BB型的分別高0.53(P<0.05)、1.1頭(P<0.01),和98 g(P>0.05);在長白群體內AA型的3個指標比BB型的分別高0.64頭(P<0.05)、0.35頭和87 g(P>0.05);在大白群體內AA型的3個指標比BB型的分別高0.86(P<0.05)、1.29頭(P<0.01)和107 g(P>0.05)。
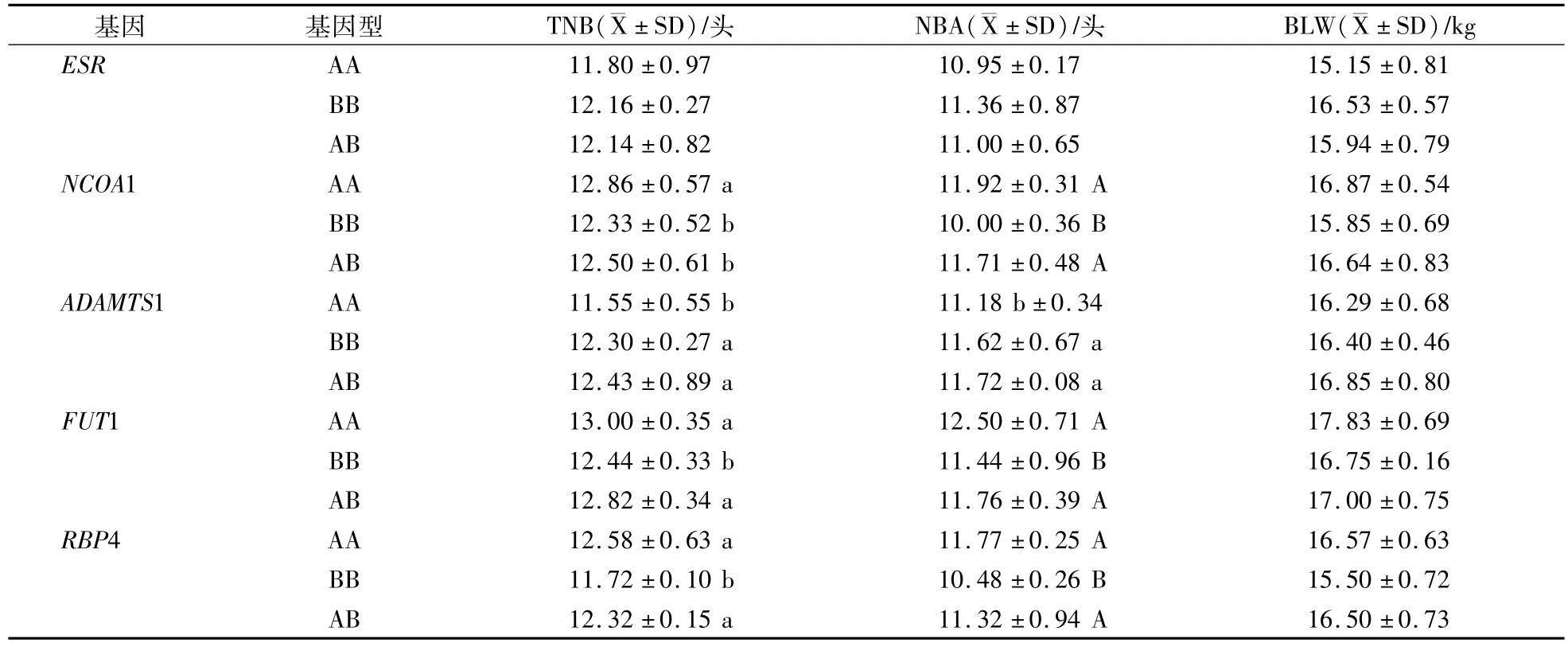
表6 5個基因對大白TNB、NBA和BLW影響Table 6 The influence of five genes to large white TNB,NBA and BLW
3 討論
3.1 ESR基因對豬繁殖性能的影響
目前國內外學者有關ESR基因對豬產仔數影響的報道中存在不同觀點。普遍認為,ESR基因不同基因型的繁殖性能表現為BB>AB>AA[11-13]。ESR基因Pvu II位點B等位基因能提高豬產仔數[1]。Short等[14]提出ESR基因B等位基因來源于中國豬種,后來經過與西方豬種雜交選育而存在。Rothschild等[1]證實ESR基因等位基因B與TNB相關聯,BB型初產TNB和NBA比AA型的多2.3頭(P<0.01),所有胎次多1.5頭(P<0.01),他也證實ESR基因對梅山豬TNB和NBA影響達到顯著水平,然而該基因對杜洛克、長白和大白TNB和NBA影響不顯著(P>0.05)。而Hunyadi-Bagi等[15]最新報道指出ESR基因在匈牙利大白、杜洛克和皮特蘭豬群體內只檢測出AA型和AB型,AA型的TNB和NBA比AB型的高。本研究結果顯示,ESR基因在大白群體內BB型的TNB和NBA比AA型和AB型多,但差異不顯著(P>0.05)。這與蘇一等[16]、Rothschild等[17]研究證實B等位基因為優勢等位基因的結果一致。本研究結果顯示ESR基因在3個豬群體內AA型頻率較高,且在杜洛克群體內未檢測出BB型。
3.2 NCOA1基因對豬繁殖性能的影響
NCOA1基因是類固醇受體共激活劑家族成員,這類輔助激活物并不直接綁定DNA發生作用,而是通過招募核激素受體啟動子來發揮作用[18]。NCOA1基因與NCOA3基因協同作用,通過調控過氧化物酶體復合物受體家族維持胎盤形成和妊娠中期胚胎存活,同時敲出這兩個基因會導致胎盤形成受阻和妊娠中期的高致死率[4]。這說明NCOA1基因與胚胎存活有關。本研究檢測出NCOA1基因對杜洛克和大白群體的繁殖性能影響表現為:AA>AB>BB型,在長白群體內只檢測到AA型。在杜洛克和大白群體內NCOA1基因AA型 NBA比 BB型的多,達到極顯著水平(P<0.01)。這與魯慧文等[19]、李永輝[20]、Melville等[21]的研究結果一致。
3.3 ADAMTS1基因對豬繁殖性能的影響
ADAMTS1為細胞外蛋白酶,在孕酮受體激活時由顆粒細胞中合成和分泌,從而促進排卵[22-23]。ADAMTS1基因不僅參與排卵,也在體外卵母細胞成熟過程中起重要作用[24]。本研究檢測ADAMTS l基因在杜洛克、長白和大白群體內AB型TNB和NBA比AA型的高,達到差異顯著水平(P<0.05);在長白和大白群體內AB型BLW比AA型的高,未達到差異顯著水平(P>0.05)。本研究結果顯示ADAMTS l基因在3個群體的繁殖性能影響表現為AB>BB>AA趨勢。這與徐珊珊等證實AB型屬于優勢基因型的結果一致[25]。
3.4 FUT1基因對豬繁殖性能的影響
張引紅等[26]研究表明FUT1基因多態性對豬TNB影響顯著(P<0.05),對NBA影響不顯著(P>0.05)。Kyungtae等[7]證實FUT1基因AA型斷奶仔豬成活率高于BB型的。有報道指出FUT1基因GG和AG型對ETEC F18敏感,AA型為抗性[27]。本研究結果顯示AA型TNB、NBA和BLW均高于AB型和BB型。盡管FUT1基因AA型仔豬成活率較高的原因未明,但FUT1基因可以抵抗傳染病的研究報道已被證實[28,5]。這可能與豬的抗病性能提高從而提高NBA有關,所以在育種過程有必要關注FUT1基因AA型的比例。
3.5 RBP4基因對豬繁殖性能的影響
RBP4基因在胚胎發育時期發揮作用[29]。大多數有關RBP4基因報道證實AA型和A等位基因對豬TNB和NBA有正向影響[30-34],但也有學者指出BB型比AA型和AB型有高的產仔數,且B等位基因來源高產仔性能的中國豬種[35-37]。在本研究結果顯示:在杜洛克、長白和大白群體內AA型TNB、NBA、BLW比BB型的高,尤其在大白群體中AA型NBA比BB型的多1.1頭,達到差異極顯著水平(P<0.01)。RBP4基因不同基因型的繁殖性能表現為AA>AB>BB的趨勢,且A等位基因是影響3個豬群體繁殖性狀的優勢等位基因。
3.6 5個基因對3個豬群體繁殖性能影響效應比較
本研究結果表明5個基因對3個豬群體繁殖性能影響程度存在差異,影響較大的是NCOA1和RBP4基因,其次是FUT1基因和ADAMTS1基因,最小的是ESR基因。NCOA1基因對杜洛克群體的TNB和NBA、大白群體NBA影響達到差異極顯著水平(P<0.01),對大白群體TNB影響達到顯著水平(P<0.05)。RBP4基因對杜洛克和大白群體NBA影響達到差異極顯著水平(P<0.01),對3個豬群體TNB影響達到顯著水平(P<0.05)。FUT1基因對大白NBA影響達到差異極顯著水平(P<0.01),對杜洛克群體TNB、NBA和大白群體TNB影響達到顯著水平(P<0.05)。ADAMTS1基因對3個群體TNB、NBA影響達到顯著水平(P<0.05)。ESR基因對3個豬群體TNB、NBA影響未達到顯著水平(P>0.05)。
造成差異原因可能與基因效應大小有關,NCOA1基因可以直接和間接影響豬胚胎的存活。本研究顯示NCOA1基因和RBP4基因優勢基因型都是AA型,這2個基因AA型頻率在杜洛克、長白、大白群體中分別為0.59、1、0.78和0.68、0.41、0.25,可作為豬繁殖性能的輔助選擇標記。最新研究證實FUT1基因AA型仔豬有較高的成活率[25]。本研究顯示FUT1基因AA型頻率在杜洛克和大白群體中分別為0.16和0.03,而在長白群體中未檢測出AA型,這暗示著3個豬群體的斷奶仔豬成活率或許不同,但這一推論需要進一步試驗加以證實。ADAMTS l基因AB型為優勢基因型,在杜洛克、長白、大白群體內頻率分別為0.50、0.46和0.44,呈現中度多態,可作為豬繁殖性能的輔助選擇標記。ESR基因有利基因型BB型在大白群體中頻率0.03,在杜洛克和長白群體中未檢測出BB型,這也可能是ESR基因對3個豬群體繁殖性能影響不大的原因。總之,5個基因的不同基因型對3個豬群體TNB和NBA影響可達到差異顯著或者極顯著水平,但它們對BLW影響未達到差異顯著水平。
[1]ROTHSCHILD M F,JACOBOSON C,VASKE D,et al.The estrogen receptor locus is associated with amajor gene influence litter size in pigs[J].Proc Natl Acad Sci U SA,1996,93(1):201-205.
[2]SUNDARRAJAN C,LIAOW,ROY A C,et al.Association of estrogen receptor gene polymorphisms with outcome of ovarian stimulation in patients undergoing IVF[J].Mol Hum Reprod,1999,5(9):797-802.
[3]SHIMADAM,NISHIBORIM,YAMASHITA Y,etal.Down-regulated expression of A disintegrin andmetalloproteinasewith thrombospondin-like repeats-1 by progesterone receptor antagonist is associated with impaired expansion of porcine cumulus-oocytecomplexes[J].Endocrinology,2004,145(10):4603-4614.
[4]CHEN X,LIU Z,XU J.The cooperative function of NCOA1 and NCOA3 in placental development and embryo survival[J].Mol Endocrinol,2010,24(10):1917-1934.
[5]BAOW B,YE L,PAN Z Y,et al.Beneficial genotype of swine FUT1 gene governing resistance to E.coli F18 is associated with important economic traits[J].JGenet,2011,90(2):315-318.
[6]V?GELIP,KUHN B,KüHNE R,et al.Evidence for linkage between the swine L blood group and the loci specifying the receptorsmediating adhesion of K88 Escherichia coli pilus antigens[J].Anim Genet,1992,23(1):19-29.
[7]KIM K,NGUYEN D T,CHOIM,et al.Alpha(1,2)-fucosyltransferase M307A polymorphism improves piglet survival[J].Anim Biotechnol,2013,24(3):243-250.
[8]YELICH JV,POMP D,GEISERT R D.Detection of transcripts for retinoic acid receptors,retinol-binding protein,and transforming growth factors during rapid trophoblastic elongation in the porcine conceptus[J].Biol Reprod,1997,57(2):286-294.
[9]HARNEY JP,OTT T L,GEISERT R D,et al.Retinol-binding protein gene expression in cyclic and pregnant endometrium of pigs,sheep,and cattle[J].Biol Reprod,1993,49(5):1066-1073.
[10]TROUTW E,MCDONNELL J J,KRAMER K K,et al.The retinol-binding protein of the expanding pig blastocyst:molecular cloning and expression in trophectoderm and embryonic disc[J].Mol Endocrinol,1991,5(10):1533-1540.
[11]李明洲,張凱,李學偉,等.中國地方豬種 ESR基因 PvuⅡ多態性及其產仔數關系的研究[J].四川農業大學學報,2003,21(3):258-262.
[12]田勇,李學偉,李芳瓊,等.四川省外種豬ESR基因對繁殖及生長性狀的影響[J].黑龍江畜牧獸醫,2004(10):12-14.
[13]張曉東,王恒,丁月云,等.安徽省4個地方豬種ESR和FSHβ基因多態性及其與產仔數的關聯分析[J].安徽農業大學學報,2014,41(4):579-584.
[14]SHORT T H,ROTHSCHILD M F,SOUTHWOOD O I,et al.Effect of the estrogen receptor locus on reproduction and production traits in four commercial pig lines[J].Anim Sci.1997,75(12):3138-3142.
[15]HUNYADI-BAGIá,BALOGH P,NAGY K,et al.Association and polymorphism study of seven candidate geneswith reproductive traits in three pig breeds in Hungary[J].Acta Biochim Pol,2016,63(2):359-364.
[16]蘇一,邊連全,劉顯軍.豬品種 ESR和 PRLR基因的 PCR-SSCP多態性分析[J].湖北農業科學,2010,49(7):1557-1561.
[17]ROTHSCHILD M F,LARSON R G,JACOBSON C,et al.Pvu II polymorphisms at the porcine oestrogen receptor locus(ESR)[J].Anim genet,1991.22(5):448.
[18]QIN L,LIU Z,CHEN H,et al.The steroid receptor coactivator-1regulates twist expression and promotes breast cancer metastasis[J].Cancer Res,2009,69(9):3819-3827.
[19]魯慧文,趙志超,徐夢思,等.NCOA1基因型對長白豬繁殖性能的影響[J].江蘇農業科學,2012,42(9):175-177.
[20]李永輝.NCOA1基因與豬產仔性狀的相關及序列分析[D].長沙:湖南農業大學,2005.
[21]MELVILLE JS,GIBBIN A M,ROBINSON JA,et al.Meishang positire QTL for prolificacy traits found at the NCOA1 locus on SSC3[C].Montpellier:Proceedings of the 7 th World Congress on Genetics Applied to Livestock Production.France.2002:19-23.
[22]RUSSELL D L,DOYLE K M,OCHSNER S A,et al.Processing and localization of ADAMTS-1 and proteolytic cleavage of versican during cumulusmatrix expansion and ovulation[J].JBiol Chem,2003,278(43):42330-42339.
[23]BREEN SM,ANDRIC N,PING T,et al.Ovulation involves the luteinizing hormone-dependent activation of G(q11)in granulosa cells[J].Mol Endocrinol,2013,27(9):1483-1491.
[24]BLAHA M,NEMCOVA L,KEPKOVA K V,et al.Gene expression analysis of pig cumulus-oocyte complexes stimulated in vitro with follicle stimulating hormone or epidermal growth factor-like peptides[J].Reprod Biol Endocrinol,2015,13:113:1-17.
[25]徐珊珊,于麗麗,樂凱.豬 ADAMTS1基因對繁殖性狀的遺傳效應分析[J].安徽農業科學,2008,36(24):10374-10376.
[26]張引紅,周忠孝,曹果清.FUT1基因多態性及其與產仔性狀的關聯性研究[J].遺傳,2007,29(1):52-56.
[27]MEIJERINK E,NEUENSCHWANDER S,FRIESR,et al.A DNA polymorphism influencing alpha(1,2)fucosyltransferase activity of the pig FUT1 enzyme determines susceptibility of small intestinal epithelium to Escherichia coli F18 adhesion[J].Immunogenetics,2000,52(1/2):129-136.
[28]WANG S J,LIUW J,YANG LG,etal.Effectsof FUT1 genemutation on resistance to infectious disease[J].Mol Biol Rep,2012,39(3):2805-2810.
[29]ROTHSCHILD M F,MESSER L,DAY A,et al.Investigation of the retinol-binding protein 4(RBP4)gene as a candidate gene for increased litter size in pigs[J].Mamm Genome,2000,11,(1):75-77.
[30]SP?TTER A,MüLLER S,HAMANN H,et al.Effect of polymorphisms in the genes for LIF and RBP4 on litter size in two German pig lines[J].Reprod Domest Anim,2009,44(1):100-105.
[31]MARANTIDISA,LALIOTIS G P,AVDIM.Retinol binding protein 4 gene and reproductive traits in pigs[J].Genet Res Int,2007,50:181-185.
[32]MESSER L A,WANG L,YELICH J,etal.Linkagemapping of the retinol-binding protein 4(RBP4)gene to porcine chromosome14[J].Mamm Genome,1996,7(5):396.
[33]孫延曉,曾勇慶,唐輝,等.豬 PRLR和 RBP4基因多態性與產仔性能的關系[J].遺傳,2009,31(1):63-68.
[34]GON?ALVES ID V,GON?ALVESPB D,SILVA JC,et al.Interaction between estrogen receptor and retinol-bindingprotein-4 polymorphisms as a tool for the selection of prolific pigs[J].Genet Mol Biol,2008,31(2):481-486.
[35]KORWIN-KOSSAKOWSKA A,KAPELANSKIW,BOCIAN M,et al.Preliminary study of the RBP4,EGF and PTGS2 genes polymorphism in pigs and its association with reproduction traits of sows[J].Anim Sci Pap Rep,2005,23(2):95-105.
[36]ISLER B J,IRVIN KM,NEAL SM,etal.Examination of relationship between estrogen receptor gene and reproductive traits in pig[J].JAnim Sci,2002,80(9):2334-2339.
[37]ALFONSO L.Use ofmeta-analysis to combine candidate gene association studies:application to study the relationship between the ESR PvuII polymorphism and sow litter size[J].Genet Sel Evol,2005,37(4):417-435.
