一個受青枯病菌誘導的煙草功能基因NtCNGC1的克隆與表達分析
2018-03-18 05:17:22林世鋒王仁剛余婧任學良張潔余世洲
中國煙草學報 2018年6期
林世鋒,王仁剛,余婧,任學良,張潔,余世洲
貴州省煙草科學研究院,煙草行業分子遺傳重點實驗室,貴州貴陽龍灘壩路29號 550081
青枯勞爾氏菌(Ralstonia solanacearum)引起的煙草青枯病是世界范圍的毀滅性土傳病害,給煙草生產帶來巨大損失[1]。目前國內外對煙草抗青枯病的分子機理研究甚少,對控制青枯病的抗性基因數目、作用機制、煙草與青枯菌互作機理還不清楚。從轉錄組水平上分析和挖掘煙草品系中可能存在的抗青枯病基因,不僅能從分子水平上加深對煙草與青枯菌互作的認識,還將有助于青枯病抗源合理利用和抗病育種工作。
環核苷酸(cAMP/cGMP)是生命體重要的信號分子,環核苷酸門控離子通道(Cyclic nucleotidegated channel,CNGC)是環核苷酸主要受體之一,參與調控植物的生長、發育以及抗病等反應[2]。目前已在植物中克隆并鑒定了多個環核苷酸門控離子通道基因,Clough等[3]在篩選植物抗病突變體時發現AtCNGC2參與了植物響應病原微生物誘導的信號轉導,病原菌(Pseudomonas syringae)在cngc2突變體上生長速度變慢,不能產生相應的過敏性反應(Hypersensitive response,HR),原因是cngc2突變體中水楊酸(Salicylic acid,SA)含量持續上升,導致病程相關(pathogenesis-related,PR)基因的持續表達,從而造成病原菌不能正常生長。電生理學分析顯示,cngc2突變體胞內cAMP激活的鈣離子流缺失,從而破壞胞內NO(nitric oxide)的產生,造成植物免疫反應的改變[4-5]。AtCNGC4也參與植物對病原微生物的抗性反應,AtCNGC4與AtCNGC2的同源性最高,其突變體與cngc2的表型一致,表現出PR基因的持續表達,水楊酸含量處于高水平等現象[6-7]。敲除AtCNGC11和AtCNGC12可降低植物對病原菌(Hyaloperonospora arabidopsidis)的抵抗能力,表明二者在植物抗病信號途徑中是正調控因子[8]。此外,大麥NEC1基因的序列與AtCNGC4基因相似,其突變體表型與cngc4的相同,表現出PR基因的上調表達[9]。因此可以認為,CNGC既參與雙子葉植物的抗病反應,又參與單子葉植物的抗病反應。
基于前期發現的CNGC基因家族成員在抗病煙草材料中上調表達的轉錄組結果,本研究克隆了該CNGC基因家族成員NtCNGC1,熒光定量PCR發現NtCNGC1在煙草DB101根中高水平表達,對比抗、感煙草材料接種青枯病菌后NtCNGC1表達量,進一步證明了該基因與煙草抗青枯病相關。
1 材料與方法
1.1 試驗材料
強致病力煙草青枯病菌由貴州省煙草科學研究院煙草行業分子遺傳重點實驗室分離鑒定并繼代保存于TTC培養基。煙草青枯病抗病品種DB101和易感品種紅花大金元由貴州省煙草科學研究院良種繁育中心提供,采用漂浮育苗,待煙苗長至2~3片真葉時,移栽至花盆中(泥炭土:蛭石:珍珠巖=1:1:1)培養。
1.2 NtCNGC1基因的克隆
根據GenBank數據庫中的煙草CNGC1基因序列(登錄號:XM_016655150)設計克隆引物CNGC1-F和CNGC1-R(表1)。提取DB101煙草葉片總RNA,并反轉錄為cDNA,作為模版對煙草CNGC1基因進行PCR擴增、克隆和測序。同時,從DB101煙草葉片中提取基因組DNA,作為模版進行PCR擴增、克隆和測序。
1.3 NtCNGC1基因的生物信息學分析
采用SMART在線工具http∶//smart.embl-heidelberg.de/檢索保守模體;采用PlantCARE在線工具http∶//bioinformatics.psb.ugent.be/webtools/plantcare/html/預測啟動子的順時作用元件;采用ClustalX 2.0軟件對齊所有CNGC序列;采用MEGA4.0軟件構建進化樹,方法為鄰接法,自舉檢測次數為1000。
1.4 NtCNGC1基因的熒光定量PCR表達分析
當煙苗長至5~6片真葉時,選取DB101和紅花大金元生長狀況一致的煙苗,參照Marco等[10]的方法接種青枯病菌,以蒸餾水處理煙苗作為對照。分別在接種后3、6、12、24和48 h取接種葉片作為檢測樣品;并在DB101苗期和旺長期,取中部葉、莖和根作為檢測樣品,所有樣品液氮速凍保存。采用Trizol試劑盒(Invitrogen公司)提取各樣品的總RNA,DNaseⅠ除去基因組DNA后,采用反轉錄試劑盒(TaKaRa公司)將其反轉錄成cDNA,熒光定量PCR進行煙草CNGC1基因的表達分析。基因擴增上游引物為CNGC1-rF、下游引物為CNGC1-rR,以煙草β-actin基因作為內參,內參基因擴增中所用上下游引物為Actin-rF和Actin-rR(表1)。各處理樣品進行3次重復。采用2-ΔΔCT法進行基因相對定量分析[11]。
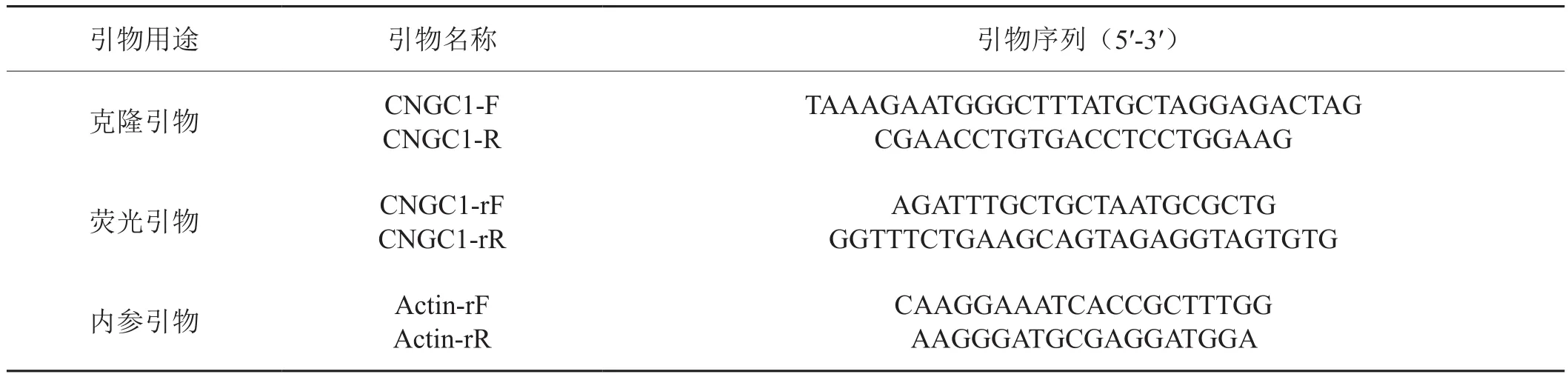
表1 NtCNGC1基因的克隆和表達分析引物Tab. 1 Primers used for cloning and expression analysis of NtCNGC1 gene
2 結果與分析
2.1 NtCNGC1基因的序列特征
分別以煙草基因組DNA和cDNA為模板,用引物對CNGC1-F/CNGC1-R擴增出目的條帶(圖1)。測序結果表明,NtCNGC1基因組DNA擴增產物長度為6519 bp,cDNA擴增產物長度為2445 bp,其中編碼區全長為2154 bp。該基因具有7個內含子,第7內含子最長,為952 bp,第6內含子次之,為659 bp,第2內含子最短,僅86 bp;8個外顯子中,第2外顯子最長,為565 bp,其次為第7外顯子,長度為476 bp,第5外顯子最短,僅112 bp(圖2)。
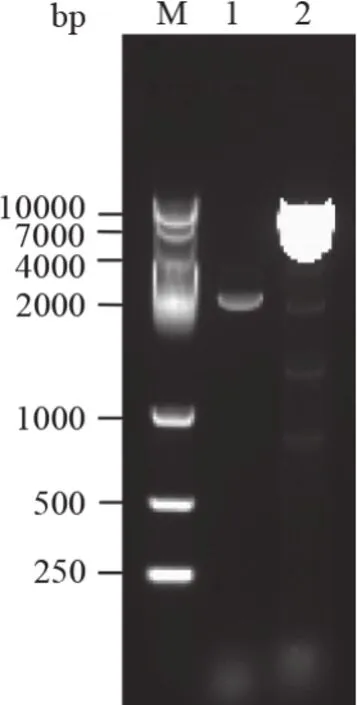
圖1 煙草NtCNGC1基因的PCR擴增結果Fig.1 PCR amplification results of NtCNGC1 from Nicotiana tabacum
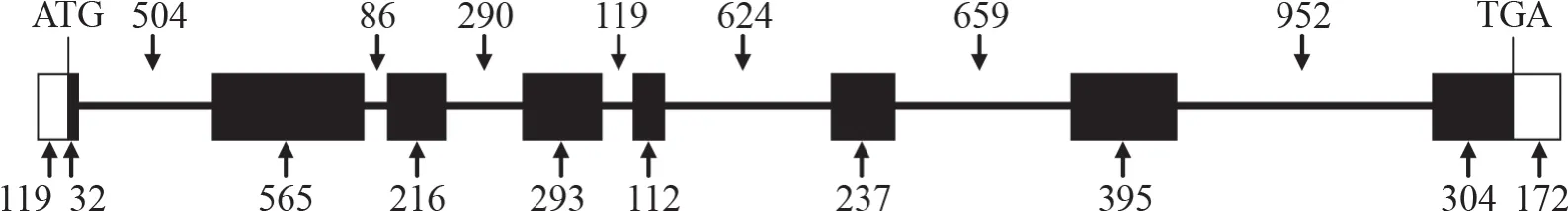
圖2 NtCNGC1的基因結構示意圖Fig.2 Gene schematic structure of NtCNGC1
2.2 NtCNGC1編碼蛋白的特征
NtCNGC1編碼717個氨基酸(圖3),等電點為9.35,分子量為83 kDa。SMART在線工具檢索發現,NtCNGC1編碼蛋白有6個保守結構域(圖4),分 別 位 于 97~118,133~152,185~207,257~279,380~402和488~619氨基酸殘基位置,N端為5個連續的跨膜結構域(Transmembrane region),C端為環化核苷酸結合域(Cyclic nucleotide-monophosphate binding domain,cNMP)。
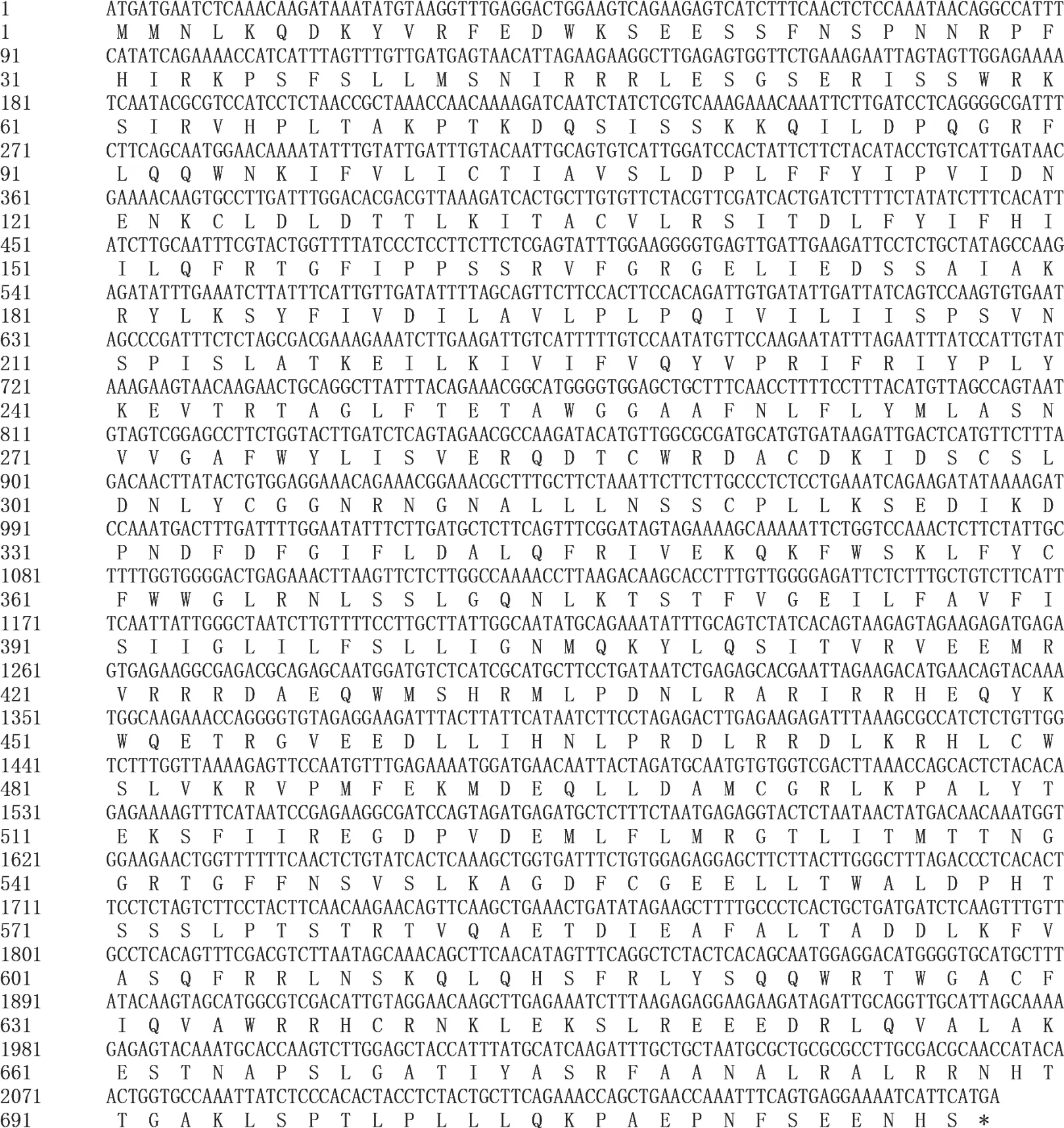
圖3 NtCNGC1的編碼區及推導的蛋白質序列Fig.3 Coding sequence and its deduced protein sequence of NtCNGC1
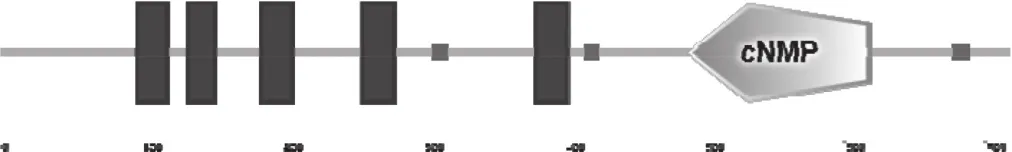
圖4 NtCNGC1的保守模體Fig.4 Conserved motifs of NtCNGC1
2.3 NtCNGC1基因的上游啟動子分析
啟動子預測結果表明,位于NtCNGC1基因起始密碼子ATG的5′區上游 1500 bp的啟動子核苷酸序列具有多種順式作用元件(表2),包括與光響應、真菌激發子響應、熱響應、干旱響應、傷口響應、水楊酸響應、厭氧誘導、防御與應激反應、病原菌應答、木質部誘導性表達和韌皮部阻遏性表達、代謝調節及器官發育等相關元件。

表2 NtCNGC1基因上游調控區順式作用元件Tab. 2 Cis-elements in the upstream regulation region of NtCNGC1 gene
2.4 NtCNGC1的系統進化分析
根據CNGC序列中的2個功能結構域,比較NtCNGC1與全部擬南芥CNGC蛋白家族成員和部分煙草、大麥、蒺藜苜蓿、水稻中假定的CNGC之間的親緣關系,結果表明(圖5),CNGC 主要分成5個亞組(Ⅰ、Ⅱ、Ⅲ、Ⅳa、Ⅳb),其中,Ⅰ、Ⅱ、Ⅲ親緣關系很近,它們與另外兩個亞組Ⅳa和Ⅳb的關系較遠。NtCNGC1與擬南芥抗病正調控因子AtCNGC11、AtCNGC12和AtCNGC11/12嵌合體同屬于CNGC家族的第Ⅰ亞組,而擬南芥抗病負調控因子AtCNGC2和AtCNGC4屬于CNGC家族的第Ⅳb亞組。
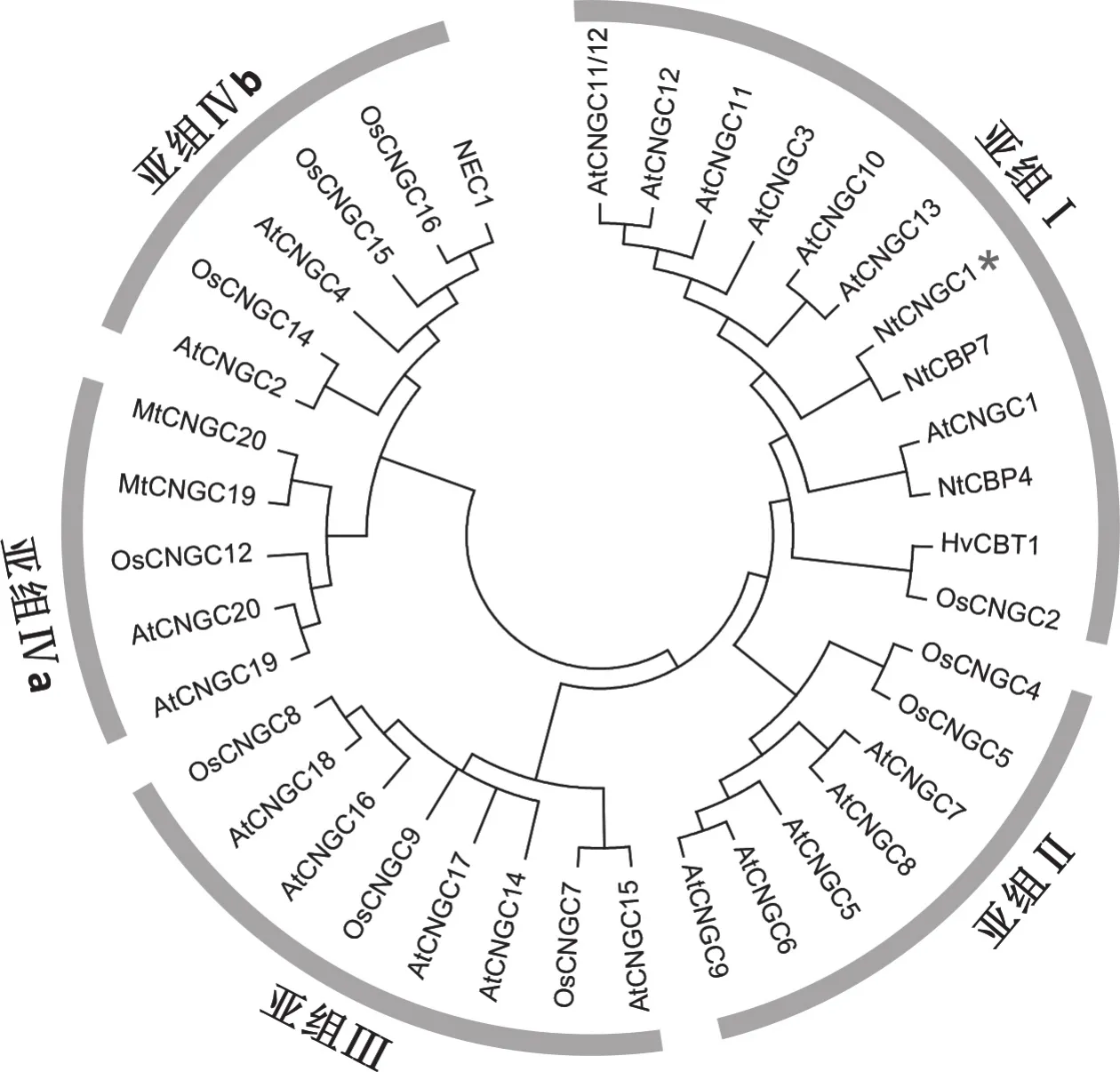
圖5 植物CNGCs的系統進化樹Fig.5 Phylogenetic tree of plant cyclic nucleotide-gated channels
2.5 NtCNGC1基因的表達分析
為了驗證NtCNGC1基因的轉錄組表達結果,利用熒光定量PCR檢測NtCNGC1響應青枯病菌侵染的表達情況。在煙草青枯病菌侵染后,抗病品種DB101中NtCNGC1基因的表達量從3 h 開始就有上調表達趨勢,而感病品種紅花大金元在12 h才表現出上調表達,且表達量前者明顯高出后者(圖6),這一表達變化趨勢與轉錄組結果基本一致。熒光定量PCR進一步檢測NtCNGC1基因在抗病煙草DB101中的組織表達情況,發現在苗期和旺長期NtCNGC1在根中表達量最高,莖中其次,葉中最低(圖7)。
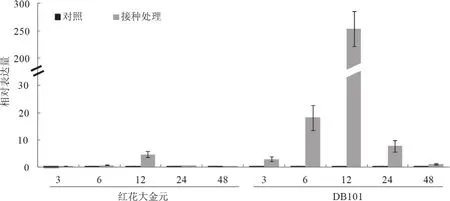
圖6 煙草接種青枯病菌病后NtCNGC1基因的表達分析Fig.6 Expression level of NtCNGC1 gene in response to Ralstonia solanacearum
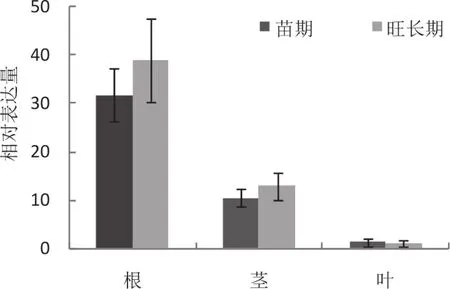
圖7 NtCNGC1基因在不同組織中的表達分析Fig.7 Expression level of NtCNGC1 gene in different organs
3 討論與結論
本課題組在前期煙草響應青枯病菌侵染的轉錄組研究中發現NtCNGC1基因(XM_016655150)可能參與抵抗煙草青枯菌侵染的防衛反應。為進一步驗證NtCNGC1基因的這一功能,本試驗克隆了NtCNGC1基因。熒光定量PCR證實NtCNGC1基因在抗病煙草品種DB101中上調表達,而在感病煙草品種紅花大金元中無明顯表達變化,初步說明NtCNGC1基因在抗病煙草中可以響應青枯菌的侵染。而且NtCNGC1基因的表達具有組織特異性,主要在根中表達,這與青枯菌在煙草根部侵染的特點相吻合。基于上述研究結果推測NtCNGC1是煙草抗青枯病相關基因。
大量研究表明,激素信號在植物抗病反應中具有重要的作用,一些抗病基因通過一個或幾個激素信號通路的調控發揮其重要功能[12-13]。本研究結果顯示,NtCNGC1基因的啟動子區域含有多個與植物抗病或激素應答相關的順式作用元件,如病原菌應答元件W-Box、真菌誘導元件Box-W1、防御及脅迫響應元件TC-rich repeats、以及水楊酸響應元件TCA-element,暗示NtCNGC1基因可能通過響應水楊酸信號參與煙草對青枯菌的抵御過程。有關CNGC在響應水楊酸信號調控植物防御反應中的作用研究已有報道,Moeder等[14]的研究結果表明,擬南芥AtCNGC2和AtCNGC4的表達受非致病性病原菌Pst DC3000(AvrRps4)和水楊酸的負調控,而擬南芥AtCNGC11和AtCNGC12的表達受非致病性病原菌Pst DC3000(avrRpt2)和水楊酸的正調控,推測前兩者在水楊酸和R基因介導的防御信號途徑中為負調控因子,而后兩者為正調控因子。考慮到煙草NtCNGC1與擬南芥AtCNGC11和AtCNGC12同屬于CNGC家族的第Ⅰ亞組,且有報道顯示水楊酸誘導煙草對煙草青枯菌的抗性[15-16],推測煙草NtCNGC1基因作為正調控因子參與水楊酸和R基因介導的煙草青枯病抗性。
綜上,本研究從煙草中克隆到NtCNGC1,生物信息學和表達分析表明該基因的表達具有組織特異性,并受青枯菌誘導,未來將通過轉基因和基因敲除方法對該基因的功能進行驗證。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
當代化工研究(2016年9期)2016-03-20 16:22:08
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
少兒科學周刊·兒童版(2015年1期)2015-07-07 04:12:52
聲屏世界(2014年6期)2014-02-28 15:18:09
