雷竹PvJ3基因克隆及功能分析
2018-03-23 09:51:09陳曉沛郭小勤曹友志徐英武
浙江農林大學學報 2018年2期
陳曉沛,施 泉,郭小勤,曹友志,徐英武
(浙江農林大學 省部共建亞熱帶森林培育國家重點實驗室,浙江 杭州311300)
開花是高等植物進入生殖生長的標志,是植物完成生活史的必要環節。在擬南芥Arabidopsis thaliana中的研究發現,開花過程受多種內源因素和外源環境因素的影響,由復雜基因網絡嚴格調控[1]。光周期途徑中植物開花受季節性晝長變化影響[2];赤霉素(GA)途徑是在非誘導光周期途徑的條件下,在開花過程中發揮作用[3];春化途徑中植物開花要受到低溫積累的影響[4];自主途徑中通過感知植物自身內部發育狀態而調控植物開花[5],多種途徑的成花調節信號最終都集中于2個整合因子:開花促進因子FLOERING LOCUS T(FT)和 SUPPRESSPR OF OVEREXPRESION OF CONSTANS 1(SOC1)[6-8]。 在擬南芥中,過表達FT基因會明顯促進SOC1基因的表達量;同時在ft突變體中,SOC1的表達量明顯下降[9-11],但是對ft和soc1雙突變體與ft單突變體植株的開花時間進行統計,結果顯示沒明顯差異,可知除FT蛋白調控SOC1的表達外,還存在另外的途徑影響SOC1基因的表達[10-13]。其他開花控制因子可以通過FT和SOC1參與開花途徑的調控,例如開花抑制因子SHORT VEGETATIVE PHASE(SVP)可以結合SOC1和FT的調控序列,對FT和SOC1產生抑制作用[14],進而下調它們的表達量。近年,發現擬南芥中屬于J蛋白家族一員的J3蛋白,可以與SVP蛋白相互作用調控SOC1和FT基因的表達量,而實現對成花信號的調節,在調控開花途徑中發揮著十分重要的作用[15]。 J蛋白(J-domain protein)是廣泛存在于動植物中的一個龐大的分子伴侶家族,該類蛋白N末端包含1個約由70個氨基酸構成的保守序列——J結構域而被命名為J蛋白[16-17]。J蛋白的J結構域由4個α-螺旋組成,第2和第3個螺旋之間有一個極為保守的由組氨酸、脯氨酸、天冬氨酸組成的HPD三肽,這是J結構域的重要特征[18];鄰近J結構域的是G/F結構域(富含甘氨酸和苯丙氨酸);之后是鋅指結構域。根據結構域的不同J蛋白可分為4類[19]:Ⅰ類J蛋白包含所有常見的結構;Ⅱ類J蛋白缺少鋅指結構;Ⅲ類J蛋白只有J結構域;Ⅳ類J蛋白也被稱作J-like蛋白,這類蛋白的J結構域沒有HPD模塊[20-21]。目前發現擬南芥中共有120個J蛋白,其中Ⅰ類J蛋白8個,Ⅱ類J蛋白16個,Ⅲ類J蛋白92個,Ⅳ類J蛋白4個[22]。在植物中,有關J蛋白的研究主要集中在擬南芥中,現有的結果表明,J蛋白可以參與多種生物學過程,包括葉綠體的發育[23-25]、植物的抗逆性[26-27]、信號轉導[28-29]和成花途徑[15]等。在擬南芥開花調控中,J3蛋白通過與SVP等其他開花關鍵因子相互作用而影響植物成花過程,SVP的突變體svp-41表現為早花現象,J3的突變j3-1表現為晚花現象,同時在j3-1和svp-41的雙突變體中,擬南芥表現出早花現象,與svp-41的單突變體的表型相似,這一結果表明J3基因通過與SVP的相互作用實現對開花時間的調控[30-31],進一步研究發現在細胞核中J3基因通過與營養生長短期(SVP)蛋白直接相互作用,衰減SVP蛋白與SOC1和FT調控序列的結合從而上調SOC1和FT基因的表達量[15]。竹類植物開花周期較長并具有時間上的不確定性,而且很多竹子開花后會成片死亡,雷竹Phyllostachys violascens是一種優良的筍用竹種,近年來,雷竹普遍開花,開花后部分雷竹會死亡,導致雷竹筍產量大幅度下降,帶來嚴重的經濟損失。有關竹類植物開花的研究已經很多,但是竹子成花機制和花期調控模式至今尚未完全清楚[32]。目前,竹子中還未見有關J3基因生物學功能的研究,本研究以雷竹為研究對象,通過同源克隆技術獲得1個J3同源基因,經過亞細胞定位、表達模式和功能分析,以期能為深入研究J3同源基因在竹類植物開花調控機制中的作用提供科學依據。
1 材料與方法
1.1 實驗材料
雷竹材料來源于浙江農林大學智能溫室大棚,選取位于同一竹鞭上的筍、花芽、花,開花和不開花雷竹的幼葉、成葉、莖稈,標記分裝后放入液氮中速凍,保存在-80℃超低溫冰箱。
1.2 核糖核酸(RNA)的提取及互補脫氧核糖核酸(cDNA)的合成
用全 RNA提取試劑盒(鼎國)分別提取采集的雷竹材料的RNA,以提取的RNA(少于0.5 g·L-1)為模板,用Prime ScriptTMRT Master Mix(Perfect Real Time)試劑盒(Clontech),反轉錄獲得雷竹各組織的cDNA,檢測合格后,于-20℃保存備用。
1.3 目的基因的克隆及序列分析
通過本地Blast,從毛竹Phyllostachys edulis基因組文庫中找到一個候選基因序列號(bphylf027h21),根據該候選基因的基因序列設計引物(J3,表1),雷竹cDNA為模板,進行聚合酶鏈式反應(PCR)擴增,瓊脂糖凝膠電泳檢測PCR擴增條帶,用膠回收試劑盒(上海生工生物公司)回收純化電泳條帶,將獲得的目的片段與pMD19(simple)-T載體置于連接儀上16℃連接1.0~3.0 h,之后轉化大腸埃希菌 Escherichia coli DH5α感受態細胞(熱轉化法),37℃恒溫培養箱培養6.0~8.0 h,挑取白色圓形單菌落,菌落PCR鑒定出陽性單克隆,送測序,得到堿基序列。
用Vector NTI預測基因序列的開放閱讀框(ORF)區,據此設計引物(PvJ3,表1),重復上述實驗過程,最終測序得到雷竹PvJ3基因序列。運用TMHMM和ProtScale分析PvJ3蛋白的結構特征和理化性質;Smart在線分析其蛋白的結構域;通過Clustal X2比對雷竹PvJ3與其他物種中的J3同源基因的氨基酸序列的相似性;并用Mega 5.0構建系統發育樹,選擇鄰近法(NJ),bootstrap回復次數為1 000次。
1.4 組織特異性表達檢測
用雷竹各組織部位的cDNA為模板,通過實時定量聚合酶鏈式反應(qRT-PCR),分析雷竹PvJ3基因的表達模式。根據實驗要求設計目的基因的定量引物(PvJ3-1,表1),毛竹的PeNTB基因作為內參基因[33]。各種雷竹組織做3次重復,避光條件下操作,參照SYBR Premix Ex TaqTMII(Takara)試劑盒要求加樣。PCR擴增程序(兩步法):預變性95℃,5 min;95℃,30 s;60℃,30 s,循環數為40。反應結束后,分析確定數據的可靠性,選擇合適的數據,用2-ΔΔCT方法[34]對數據進行分析。
1.5 亞細胞定位
根據PvJ3基因ORF區序列(不含終止密碼子)和pCaM35S-sGFP載體序列上的酶切位點,在目的基因ORF區的上下游分別引入酶切位點SmaI和BamHI,設計引物(PvJ3-2,表1),提取pMD19-PvJ3質粒做為模板,進行PCR反應,膠回收純化得到帶有酶切位點的基因片段與pMD19(simple)-T載體連接轉化后,陽性單克隆送測序。將測序正確的單克隆進行擴大培養,用質粒提取試劑盒(上海生工生物公司)提取質粒,用限制性內切酶SmaI和BamHI分別對pMD19-PvJ3和pCaM35S-sGFP載體質粒進行雙酶切實驗,之后用T4 DNA連接酶在16℃條件下過夜連接膠回收的酶切產物,連接產物轉化大腸埃希菌DH5α感受態細胞,經測序正確,瞬時表達載體PvJ3-GFP構建成功。通過基因槍轟擊洋蔥Allium cepa表皮,25℃下避光暗培養1~2 d后,用熒光顯微鏡觀察綠色熒光蛋白信號,金粉的準備和樣品的制備參照文獻[35]。
1.6 構建轉基因過表達載體
根據PvJ3基因ORF區序列和pC1301載體(含有35 S啟動子)的序列設計引物(PvJ3-3,表1),在目的基因ORF區的上下游同樣分別引入酶切位點SmaI和BamHI,構建過表達載體pC1301-PvJ3,實驗方法參照1.5節。
1.7 擬南芥轉基因植株檢測
通過熱轉化法將構建成功的pC1301-PvJ3過表達載體轉入農桿菌Agrobacterium tumefaciens GV3101感受態細胞中,提取陽性單克隆的質粒,重新轉化大腸埃希菌DH5α感受態細胞,再次測序檢驗轉入重組質粒是否正確。用已正確轉入pC1301-PvJ3重組載體的農桿菌配制侵染液,侵染進入花期的野生型擬南芥花序[36],侵染3次,間隔約1周·次-1,侵染后的擬南芥即T0代植株,將消毒處理后的T0代種子均勻撒在加入潮霉素的1/2 MS(Murashige and Skoog)固體培養基上,潮霉素質量濃度為100.00 mg·L-1,按照V(潮霉素溶液)∶V(1/2MS培養基)=1.50∶1 000.00比例加入培養基中,篩選抗性植株,以采用十六烷基三甲基溴化銨(CTAB)法[37]提取的抗性植株的基因組 DNA為模板,PCR鑒定出T1代陽性植株,并對植株表型進行初步統計。

表1 本研究所用的引物Table 1 PCR primers used in this study
2 結果與分析
2.1 基因的克隆和序列分析
根據在毛竹轉錄組數據庫中挑選的基因設計引物,利用同源克隆的方法,從雷竹中分離得到1個J3同源基因,命名為PvJ3,分析PvJ3基因的堿基序列,可知,其ORF區由1 260 bp個堿基組成,能編碼419個氨基酸。經蛋白結構域和氨基酸序列分析(圖1~2),可知PvJ3蛋白的N端有J蛋白家族典型的J結構域,同時具有鋅指結構域,C末端有法尼基化信號CAQQ序列。用ClustalX2軟件對不同物種的 J3同源蛋白的氨基酸進行序列比對,結果發現:J3蛋白在各類物種中的相似性很高,與擬南芥J3蛋白的一致性可達80.90%(圖2)。采用臨近法構建系統進化樹,結果顯示:雷竹J3蛋白與單子葉植物高粱Sorghum bicolor和玉米Zea mays的親緣關系最近,水稻Oryza sativa,大麥Hordeum vulgare和小麥Triticum aestivum次之,與雙子葉擬南芥和亞麻薺Camelina sativa的親緣關系最遠(圖3)。
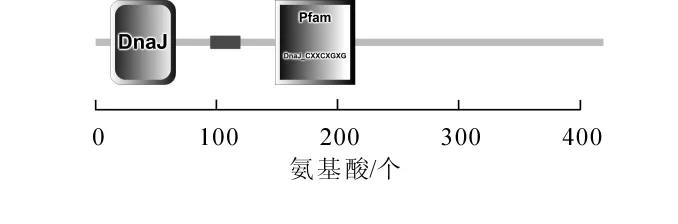
圖1 PvJ3蛋白結構示意圖Figure 1 Structure diagram of PvJ3
2.2 組織特異性表達分析
通過qRT-PCR技術,對同一竹鞭上開花和不開花雷竹中PvJ3基因在不同組織部位的表達水平進行分析。結果表明:PvJ3基因在同一竹鞭的筍、花芽、花,開花與不開花的莖稈、成葉、幼葉中均有表達。在開花雷竹的莖稈中PvJ3基因的表達量最高,莖、花芽、花、幼葉和成葉中的表達依次降低。在不開花雷竹的莖稈中PvJ3基因的表達量同樣也是最高,幼葉、成葉中表達量基本一樣(圖4)。
2.3 亞細胞定位結果分析
用基因槍轟擊洋蔥表皮細胞,通過熒光顯微鏡觀察瞬時表達載體PvJ3-GFP綠色熒光蛋白的信號,確定PvJ3蛋白的定位情況(圖5)。pCaM35S-GFP對照在整個洋蔥細胞里均有熒光信號,而PvJ3-GFP在細胞質核細胞核里有熒光信號,因此推斷PvJ3蛋白主要定位在細胞質和細胞核中。
2.4 PvJ3基因在擬南芥中的過量表達分析
構建過表達載體pC1301-PvJ3轉化野生型擬南芥收取T0代種子,潮霉素篩選抗性植株,以抗性植株基因組DNA為模板,經PCR檢測得到T1代擬南芥共37株。觀察統計T1代植株中長勢較好的30株轉基因植株的表型,發現PvJ3基因的過表達植株開花時間明顯早于野生型(圖6B)。統計結果顯示:轉基因植株開花天數比野生型植株顯著提早(P=6.51E-15<0.01),平均提早3.50 d(圖6C),開花時蓮座葉數目比野生型植株顯著減少(P=9.24E-09<0.01),平均減少4.43葉(圖6D),且表現出多主莖現象(圖6A)。
3 討論
高等植物成花過程存在多種誘導途徑,大量基因參與這些途徑的調控,成花關鍵基因的研究對于開花過程及花發育調控有著重要意義[38]。擬南芥中研究發現,SOC1和FT是感知不同開花途徑信號的整合因子,SVP是直接調控 FT和 SOC1的轉錄調控因子[8,39],J3蛋白通過與SVP直接相互作用而衰減SVP與SOC1和FT調控序列的結合從而上調SOC1和FT的表達量,參與擬南芥的成花過程,這一發現使得擬南芥成花調控網絡更加完善[15]。本研究成功從雷竹中克隆得到J3同源基因并命名為PvJ3,雷竹PvJ3所編碼的氨基酸序列與擬南芥J3蛋白氨基酸序列一致性達80.90%,對其蛋白結構域分析可知,PvJ3擁有相對保守的J結構域以及鋅指結構域,可以說明PvJ3屬于J家族中的一員。
為了研究雷竹PvJ3蛋白的功能,本研究通過qPCR技術和亞細胞定位分析該蛋白在雷竹中的表達模式和表達部位。對雷竹PvJ3基因進行了不同組織部位特異性表達分析,結果顯示雷竹PvJ3基因在同一竹鞭的不同組織部位都有表達,已發現雙子葉植物擬南芥J3基因同樣在根、莖、葉、花芽、花和果莢等不同組織中表達[29],說明J3基因在單雙子葉植物中均表現出全株表達的現象。在相對表達量上,PvJ3基因在開花雷竹莖稈中表達量最高,在花芽和花中的表達量降低,這與已報道的擬南芥中J3基因在花中表達量最高[15]的結果存在差異。PvJ3蛋白亞細胞定位結果顯示該蛋白定位在細胞質和細胞核,與擬南芥中J3蛋白的亞細胞定位結果相一致[15],擬南芥中和J3蛋白具有互作關系的SVP,SOC1和FT蛋白都是核蛋白,說明雷竹PvJ3可能在細胞核中與這些開花整合因子相互作用,從而對開花途徑進行調控。

圖2 PvJ3與其同源基因的氨基酸序列比對Figure 2 Alignment of the PvJ3 amino acid sequences from different plant species
將PvJ3基因導入野生型擬南芥植株驗證其功能,T1代轉基因植株的表型統計結果表明:轉基因擬南芥植株開花時間明顯早于野生型植株,開花時蓮座葉數量明顯減少(圖6 C,圖6D),而擬南芥J3基因的過表達植株和野生型植株的開花時間并沒有明顯差別[15],這說明雷竹PvJ3基因可以顯著影響花期,可能是一個開花促進因子。值得注意的是,轉雷竹PvJ3基因擬南芥植株同時表現出多主莖現象(圖6A),這一現象在擬南芥J3過表達植株中并沒有被觀測到[15]。 以上結果表明, 雷竹PvJ3基因不僅在成花調控中的功能和擬南芥J3基因存在一定差異,同時還可能參與莖的發育等其他生長發育過程,在基因功能上比擬南芥同源基因要復雜的多。
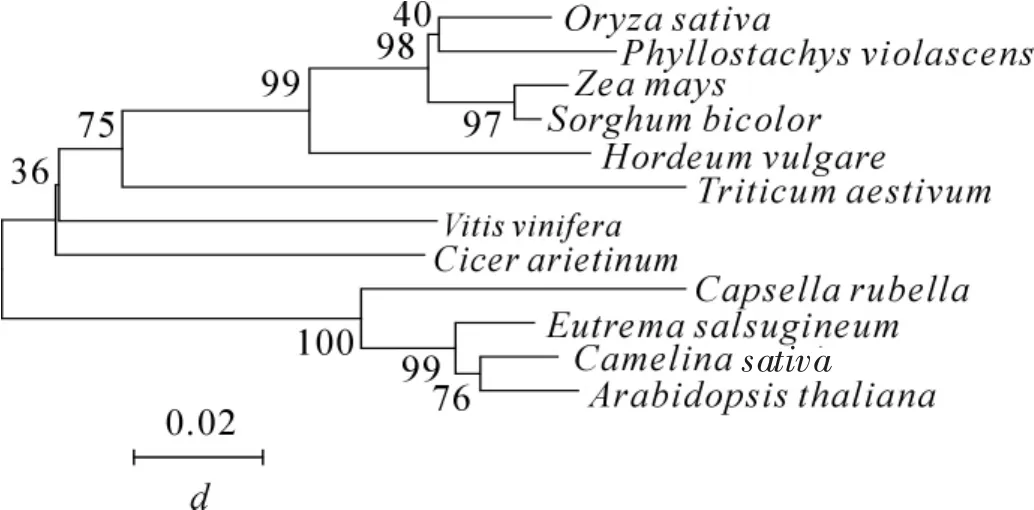
圖3 不同物種J3蛋白的系統進化樹分析Figure 3 Phylogenetic tree analysis of J3 in different plant species
目前,J3基因功能的有關研究主要以模式植物擬南芥為研究對象,單子葉植物中還未見報道。通過對比J3基因在雷竹和擬南芥中的表達模式不難發現:雷竹PvJ3基因和擬南芥J3基因都是全株表達,同時編碼蛋白都定位在細胞質和細胞核,這表明J3基因在不同植物中的表達部位不存在明顯差異,并且可能是一個核內作用蛋白。在相對表達量上,雷竹PvJ3基因和擬南芥J3基因存在一定差異,同一竹鞭上開花和不開花雷竹中PvJ3基因在莖稈中表達量最高,而擬南芥中J3基因表達量在花中最高,這可能與雷竹PvJ3基因參與莖的發育有關。轉基因實驗的結果表明:雷竹PvJ3基因不僅可以顯著提早擬南芥開花時間,同時還促使擬南芥出現多主莖現象,這表明雷竹PvJ3基因很可能參與除了開花調節以外的其他生長發育過程的調控。由于擬南芥和竹子之間的差異很大,雷竹PvJ3基因在竹子中是否可以促進植物提早開花,同時還參與哪些生物學過程,需要后續研究進一步驗證。
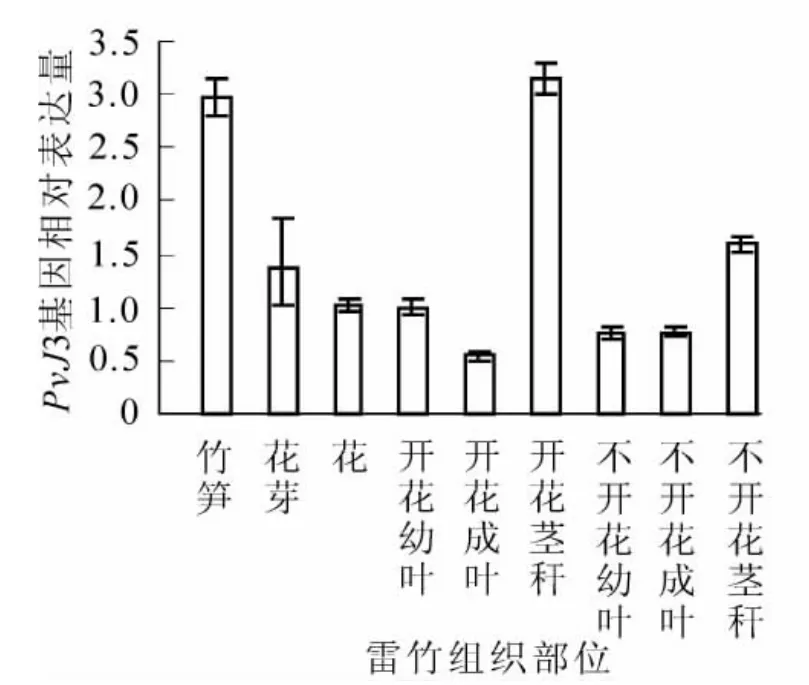
圖4 PvJ3在開花雷竹與不開花雷竹的不同部位表達水平Figure 4 Expression of PvJ3 in different tissues of Phyllostachys violascens
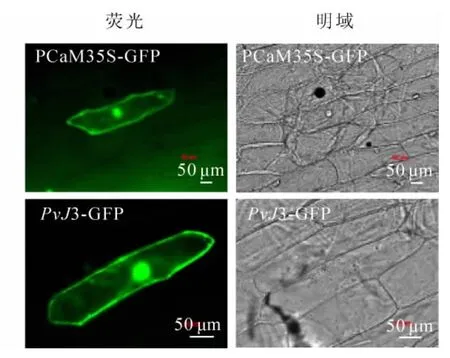
圖5 PvJ3蛋白亞細胞定位結果Figure 5 Subcellular localization of PvJ3 fusion protein
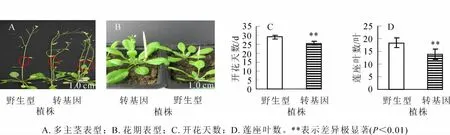
圖6 PvJ3過表達擬南芥T1植株表型Figure 6 Phenotypes of the PvJ3 transgenic Arabidopsis thaliana in T1generation
[1] QUESADA V,DEAN C,SIMPSON G G,et al.Regulated RNA processing in the control of Arabidopsis flowering [J].Int J Dev Biol,2005,49(5/6):773-780.
[2] SIMPSON G G,DEAN C.Arabidopsis,the Rosetta stone of flowering time? [J].Science,2002,296(5566):285-289.
[3] LIU Chang,CHEN Hongyan,HONG Linger,et al.Direct interaction of AGL24 and SOC1 integrates flowering signals in Arabidopsis [J].Development,2008,135(8):1481-1491.
[4] DENNIS E S,PEACOCK W J.Epigenetic regulation of flowering [J].Curr Opin Plant Biol,2007,10(5):520-527.
[5] BOSS P K,BASTOW R M,MYLNE J S,et al.Multiple pathways in the decision to flower:enabling,promoting,and resetting [J].Plant Cell,2004,16(suppl):S18-S31.
[6] WANG Cunxi,TIAN Qing,HOU Zhenglin,et al.The Arabidopsis thaliana AT PRP39-1 gene,encoding a tetratricopeptide repeat protein with similarity to the yeast pre mRNA processing protein PRP39,affects flowering time [J].Plant Cell Rep,2007,26(8):1357-1366.
[7] MOURADOV A,CREMER F,COUPLAND G.Control of flowering time interacting pathways as a basis for diversity[J].Plant Cell,2002,14(suppl):S111-S130.
[8] PENG Zhenhua,LU Ying,LI Lubin,et al.The draft genome of the fast-growing non-timber forest species moso bamboo (Phyllostachys heterocycla) [J].Nat Gen,2013,45(4):456-461.
[9] TORTI S,FORNARA F.AGL24 acts in concert with SOC1 and FUL during Arabidopsis floral transition [J].Plant Sign Behav,2012,7(10):1251-1254.
[10] NAVARRO C,CRUZ-ORó E,PRAT S.Conserved function of FLOWERING LOCUS T (FT) homologues as signals for storage organ differentiation [J].Curr Opin Plant Biol,2015,23(1):45-53.
[11] YAMASHINO T,YAMAWAKI S,HAGUI E,et al.Clock-controlled and FLOWERING LOCUS T (FT)-dependent photoperiodic pathway in Lotus japonicus I:verification of the flowering-associated function of an FT homolog [J].Biosci Biotechnol Biochem,2010,77(4):747-753.
[12] NAKANO Y,HIGUCHI Y,YOSHIDA Y,et al.Environmental responses of the FT/TFL1 gene family and their involvement in flower induction in Fragaria × ananassa [J].J Plant Physiol,2015,177(24):60-66.
[13] LEE J,LEE I.Regulation and function of SOC1,a flowering pathway integrator [J].J Exp Bot,2010,61(9):2247-2254.
[14] LEE J H,YOO S J,PARK S H,et al.Role of SVP in the control of flowering time by ambient temperature in Arabidopsis [J].Genes Dev,2007,21(4):397-402.
[15] SHEN Lisha,KANG Y G G,LIU Lu,et al.The J-domain protein J3 mediates the integration of flowering signals in Arabidopsis [J].Plant Cell,2011,23(2):499-514.
[16] RAJAN V,D’SILVA P.Arabidopsis thaliana J-class heat shock proteins:cellular stress sensors [J].Funct Int Genom,2009,9(4):433-446.
[17] MIERNYK J A.The J-domain proteins of Arabidopsis thaliana:an unexpectedly large and diverse family of chaperones [J].Cell Stress Chap,2001,6(3):209-218.
[18] QIAN Yanqiu,PATEL D,HARTL F U,et al.Nucleic magnetic resonance solution structure of the human Hsp40(HDJ-1) J-domain [J].J Mol Biol,1996,260(2):224-235.
[19] CRAIG E A,HUANG P,ARON R,et al.The diverse roles of J-proteins,the obligate Hsp70 co-chaperone [J].Rev Physiol Biochem Pharmacol,2006,156(1):1-21.
[20] MARTINEZ-YAMOUT M,LEGGE G B,ZHANG Ouwen,et al.Solution structure of the cysteine-rich domain of the Escherichia coli chaperone protein DnaJ [J].J Mol Biol,2000,300(4):805-818.
[21] MIERNYK J A.The J-domain proteins of Arabidopsis thaliana:an unexpectedly large and diverse family of chaperones [J].Cell Stress Chap,2001,6(3):209-218.
[22] SALAS-MU?OZ S,RODRíGUEZ-HERNáNDEZ A A,ORTEGA-AMARO M A,et al.Arabidopsis AtDjA3 null mutant shows increased sensitivity to Abscisic acid,salt,and osmotic stress in germination and post-germination stages[J].Plant Sci,2016,7(2):220.doi:10.3389/fpls.2016.00220.
[23] VITHA S,FROEHLICH J E,KOKSHAROVA O,et al.ARC6 is a J-domain plastid division protein and an evolu-tionary descendant of the cyanobacterial cell division protein Ftn2 [J].Plant Cell,2003,15(8):1918-1933.
[24] CHEN Kunming,HOLMSTROM M,RAKSAJIT W,et al.Small chloroplast-targeted DnaJ proteins are involved in optimization of photosynthetic reactions in Arabidopsis thaliana [J].BMC Plant Biol,2010,10(1):43.doi:10.1186/1471-2229-10-43.
[25] SHIMADA H,MOCHIZUKI M,OGURA K,et al.Arabidopsis cotyledon-specific chloroplast biogenesis factor CYO1 is a protein disulfide isomerase [J].Plant Cell,2007,19(10):3157-3169.
[26] SEKI M,NARUSAKA M,ABE H,et al.Monitoring the expression pattern of 1300 Arabidopsis genes under drought and cold stresses by using a full-length cDNA microarray [J].Plant Cell,2001,13(1):61-72.
[27] CHAI Tuanyao,ZHANG Yuxiu,ZHAO Wenming.Cloning of cDNA and expression analysis of DnaJ-like gene under heavy metal stress in bean [J].Progr Nat Sci,2000,10(2):135-140.
[28] SEDBROOK J C,CHEN Rujin,MASSON P H.ARG1 (altered response to gravity) encodes a DnaJ-like protein that potentially interacts with the cytoskeleton [J].Proc Natl Acad Sci USA,1999,96(3):1140-1145.
[29] GUAN Changhui,ROAEN E S,BOONSIRICHAI K,et al.The ARG1-LIKE2 gene of Arabidopsis functions in a gravity signal transduction pathway that is genetically distinct from the PGM pathway [J].Plant Physiol,2003,133(1):100-112.
[30] THEIβEN G,KIM J T,SAEDLER H.Classification and phylogeny of the MADS-box multigene family suggest defined roles of MADS-box gene subfamilies in the morphological evolution of eukaryotes [J].J Mol Evol,1996,43(5):484-516.
[31] WIGGE P A,KIM M C,JAEGER K E,et al.Integration of spatial and temporal information during floral induction in Arabidopsis [J].Science,2005,309(5737):1056-1059.
[32] de CARVALHO A L,NELSON B W,BIANCHINI M C,et al.Bamboo-dominated forests of the southwest Amazon:detection,spatial extent,life cycle length and flowering waves [J].PLoS One,2013,8(1):e54852.doi:10.1371/journal.pone/0054852.
[33] 齊飛艷,胡陶,彭鎮華,等.毛竹實時熒光定量PCR內參基因的篩選及成花基因PheTFL1表達分析[J].西北植物學報, 2013, 33(1): 48-52.QI Feiyan,HU Tao,PENG Zhenhua,et al.Screening of reference genes used in qRT-PCR and expression analysis of PheTFL1 gene in moso bambo [J].Acta Bot Boreal-Occident Sin,2013,33(1):48-52.
[34] LIVAK K J,SCHMITTGEN T D.Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCTmd [J].Methods,2001,25(4):402-408.
[35] 王小利,吳佳海,劉曉霞,等.高羊茅春化基因FaVRN1亞細胞定位與差異表達分析[J].基因組學與應用生物學, 2011, 30(2): 152-158.WANG Xiaoli,WU Jiahai,LIU Xiaoxia,et al.Subcellular localization and differential expression analysis of vernalizational gene FaVRN1 in tall fescue [J].Genomics Appl Biol,2011,30(2):152-158.
[36] CLOUGH S J,BENT A F.Floral dip:a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana [J].Plant J Cell Mol Biol,1998,16(6):735-743.
[37] 高志民,范少輝,高健,等.基于CTAB法提取毛竹基因組DNA的探討[J].林業科學研究,2006,19(6):725-728.GAO Zhimin,FAN Shaohui,GAO Jian,et al.Extract genomic DNA form Phyllostachys edulis by CTAB-based method [J].For Res,2006,19(6):725-728.
[38] 張素芝,左建儒.擬南芥開花時間調控的研究進展[J].生物化學與生物物理進展,2006,33(4):301-309.ZHANG Suzhi,ZUO Jianru.Advance in the flowering time control of Arabidopsis [J].Prog Biochem Biophys,2006,33(4):301-309.
[39] LI Dan,LIU Chang,SHEN Lisha,et al.A repressor complex governs the integration of flowering signals in Arabidopsis [J].Dev Cell,2008,15(1):110-120.
