miR-451對肝癌細胞增殖、糖酵解及相關基因表達的影響
2018-03-23 03:08:06周艷華張鵬飛戴廣海
現代中西醫結合雜志 2018年9期
周艷華,茅 慧,張鵬飛,戴廣海
(解放軍總醫院,北京 100853)
腫瘤細胞生長增殖活躍,其代謝機制也相當復雜,其中活躍的糖酵解代謝是惡性腫瘤細胞顯著的生化特征。惡性腫瘤細胞主要以碳水化合物通過糖酵解產能,而這一現象也被稱為沃伯格效應,該效應是細胞惡性轉化過程中最根本的代謝改變之一[1]。這種代謝方式的轉變為針對糖酵解途徑進行抑制靶向治療腫瘤細胞提供了可能性[2-3]。microRNA(miRNA)為廣泛存在于細胞內的非編碼小RNA,主要作用是調控基因的表達[4]。新近的研究顯示,miRNA對腫瘤細胞糖酵解途徑的異常調控是影響腫瘤細胞生物學特性,促進其快速生長、增殖的重要原因之一,而miR-451作為抑癌基因在惡性腫瘤的發生發展中發揮著重要作用[5-6],其中miR-451在惡性腫瘤細胞糖酵解途徑中所起的調控作用尤其值得關注[7-8]。本研究通過轉染來增強或抑制肝癌細胞中miR-451的表達,旨在探討miR-451對肝癌細胞糖酵解以及相關信號通路和基因表達的影響,為miRNA在腫瘤代謝方面的研究提供理論依據。
1 實驗資料
1.1 主要材料與試劑
人肝癌細胞MHCC97購自上海復旦大學中山醫院肝癌研究所;四甲基偶氮唑鹽(MTT)購自Sigma公司;DMEM F12培養基、小牛血清購于Gibco公司;TIANscript RT Kit 購自天根生化科技(北京)有限公司;Hot Start Fluorescent PCR Core Reagent Kits(SYBR Green I) 購自Ambion公司;葡萄糖檢測試劑盒、乳酸檢測試劑盒購自美國Invitrogen公司;miR-451、U6引物序、miR-451 mimics和miR-451 inhibitors購自Applera公司;CO2培養箱購自美國Thermo公司;XL-71臺式高速冷凍離心機購自德國Eppendorf公司;Prism?7300實時熒光定量PCR儀購自ABI公司。
1.2方法
1.2.1細胞培養將MHCC97細胞接種于DMEM F12 培養液(10%小牛血清+青霉素100 IU/mL +鏈霉素100 IU/mL)中,置于細胞培養箱中培養(孵育條件:37 ℃+5% CO2); 0.25%胰蛋白酶消化、傳代,取對數生長期細胞進行實驗。
1.2.2細胞轉染 實驗分為普通組、增強組和抑制組。將MHCC97細胞接種于6孔板中,培養24 h(細胞融合度達60%左右)后,增強組和抑制組按脂質體Lipofectamine2000說明進行SP細胞miR-451 mimics和miR-451 inhibitors的轉染,轉染48 h后采用RT-PCR法檢測3組細胞中miR-451表達量。
1.2.3MTT實驗 實驗分為普通組、增強組和抑制組。將MHCC97細胞接種于96孔板培養,平均每孔5×104個細胞,培養24 h后,每孔加入20 μL MTT(5 mg/mL),置于37 ℃、5% CO2飽和濕度條件下孵育4 h。終止反應后棄上清液,加入150 μL DMSO,振蕩10 min。在全自動酶標儀上在490 nm波長處檢測各孔的吸光度(OD),計算肝癌細胞增殖情況并繪制細胞生長曲線圖。
1.2.4肝癌細胞糖酵解相關基因表達檢測 實驗分為普通組、增強組和抑制組,采用實時熒光定量PCR法檢測各組AMPK/mTOR、Ki-67、血管內皮生長因子(VEGF)、基質金屬蛋白酶9 (MMP-9)、胰島素樣生長因子(IGF-1)表達情況。 ①細胞總RNA提取:采用Trizol法抽提濃縮組織RNA(每1×106~1×107細胞加入1 mL Trizol試劑)。Agilent 2100 生物分析儀檢測RNA分子完整性,NanoDrop@ ND-1000測定RNA濃度和純度。② cDNA 合成:按照TIANscript RT Kit 的說明進行操作。反應體系:Total RNA 40 μg,10×DNase I buffer 5 μL, RNase Inhibitor 20 IU,DNase I(RNase-free) 2 μL (10 IU),加入RNase free dH2O至總體積50 μL。反應條件:42 ℃反轉錄50 min,85 ℃ 5 min滅活反轉錄酶。③ RT-PCR反應:構建20 μL的反應體系,即RT product 1 μL, 10 μmol/L 的PCR特異引物F 1 μL,10 μmol/L 的PCR特異引物R 1 μL,2×Master Mix 10 μL,Nucleasr-Free Water 7 μL。反應條件:95 ℃ 30 s;40個循環,95 ℃ 5 s;60 ℃ 34 s。以U6為內參基因,采用2-ΔΔCT法分析檢測基因表達的相對定量值。
1.2.5葡萄糖消耗和乳酸產量檢測 實驗分為普通組、增強組和抑制組,將轉染前后MHCC97細胞以1×105/孔接種于24孔板,培養24 h后收集培養基,3 000 r/min離心10 min去除細胞碎片,取上清。按葡萄糖檢測試劑盒和乳酸檢測試劑盒說明書檢測并計算各組MHCC97細胞葡萄糖消耗值和乳酸產量。
1.3 統計學方法
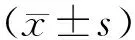
2 結 果
2.1
各組肝癌細胞中miR-451表達情況 普通組MHCC97細胞中miR-451的相對表達量為0.476±0.052,增強組MHCC97細胞中miR-451的相對表達量為1.153±0.242,明顯高于普通組(P<0.05);抑制組MHCC97細胞中miR-451的相對表達量為0.284±0.030,明顯低于增強組(P<0.05)。
2.2
各組肝癌細胞增殖情況 3組轉染前OD值比較差異均無統計學意義(P均>0.05)。轉染24 h和48 h后,增強組OD值均明顯低于普通組和抑制組(P均<0.05),抑制組OD值均明顯高于普通組(P均<0.05)。見表1。
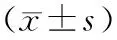
表1 3組轉染前后不同時間肝癌細胞增殖情況
注:①與增強組比較,P<0.05;②與普通組比較,P<0.05。
2.3
各組肝癌細胞糖酵解相關基因表達情況 增強組肝癌細胞AMPK mRNA表達量明顯高于普通組和抑制組(P均<0.05),而抑制組明顯低于普通組(P<0.05);增強組肝癌細胞mTOR、Ki-67、MMP-9、VEGF和IGF-1 mRNA表達量均明顯低于普通組和抑制組(P均<0.05),而抑制組均明顯高于普通組(P均<0.05)。見表2。
2.4
各組肝癌細胞糖酵解情況 轉染前3組肝癌細胞葡萄糖消耗值和乳酸分泌值比較差異均無統計學意義(P均>0.05)。轉染后增強組葡萄糖消耗值和乳酸分泌值均明顯低于轉染前及普通組和抑制組(P均<0.05),抑制組葡萄糖消耗值和乳酸分泌值均明顯高于轉染前和普通組(P均<0.05)。見表3。
表2 3組肝癌細胞糖酵解相關基因表達情況
注:①與增強組比較,P<0.05;②與普通組比較,P<0.05。
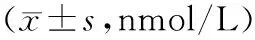
表3 3組轉染前后肝癌細胞葡萄糖消耗和乳酸分泌量比較
注:①與轉染前比較,P<0.05;②與增強組比較,P<0.05;③與普通組比較,P<0.05。
3 討 論
惡性腫瘤細胞存在明顯的能量代謝異常,而沃伯格效應是目前公認的惡性腫瘤細胞最重要的能量代謝特性,即在氧充足的條件下,腫瘤細胞無法通過線粒體氧化磷酸化方式獲取更多的能量,而只能通過糖酵解方式進行葡萄糖代謝以滿足其活躍的合成代謝需求。沃伯格效應的發現為惡性腫瘤的治療提供了新的途徑和思路,即針對糖酵解途徑的靶向干預來抑制腫瘤細胞的能量代謝[9-10]。miRNA是普遍存在于真核細胞,具有高度保守序列的小RNA。目前已知miRNA在細胞的生長、增殖、凋亡、分化、信號轉導及代謝等生物學進程中起著非常重要的調控作用[11],其能夠對糖酵解相關基因進行調控,導致糖酵解相關酶的表達及活性異常,最終影響腫瘤細胞的生長和增殖[12-13]。
miR-451為新近發現的具有抑癌作用的小RNA,與多種惡性腫瘤的增殖、侵襲和轉移密切相關[14-15]。Godlewski等[16]的研究顯示,miR-451在膠質瘤細胞中呈低表達,高葡萄糖條件下會使細胞miR-451表達增強,抑制AMPK功能,進而抑制膠質細胞瘤增殖遷移;而當葡萄糖缺乏時會激活AMPK信號,抑制miR-451表達,一方面增強膠質瘤細胞遷移能力,另一方面增加對mTOR的抑制,抑制膠質瘤細胞的增殖。進一步通過熒光素酶報告基因分析提示miR-451 通過直接抑制靶基因CAB39的表達,進而抑制LKB1/AMPK信號傳導。本研究中,通過脂質體miR-451 mimics和inhibitors轉染來增強和抑制肝癌細胞miR-451的表達,結果顯示增強miR-451表達能抑制肝癌細胞增殖,減少肝癌細胞葡萄糖消耗值和乳酸分泌值;抑制miR-451表達則促進肝癌細胞的增殖,會增加肝癌細胞葡萄糖消耗值和乳酸分泌值。提示過表達miR-451會抑制肝癌細胞糖酵解,抑制miR-451表達則會增強肝癌細胞糖酵解。
miR-451能夠通過抑制糖酵解來抑制腫瘤細胞的生長,但是其涉及的信號通路和基因目前尚未明確。AMPK/mTOR通路是已知的調控腫瘤細胞生長和能量代謝的一條重要的信號途徑[17]。mTOR基因活化能夠促進細胞糖酵解,進而促進細胞生長和增殖,AMPK則負向調控mTOR的功能。當細胞能量缺乏時,AMP/ATP比例升高,激活AMPK并磷酸化形成TSC1/2二聚體,抑制mTOR活性[18-19]。Ki-67、MMP-9、VEGF和IGF-1是目前公認的癌基因,這些指標高表達提示腫瘤生長增殖活躍,易發生轉移,且Ki-67、MMP-9、VEGF和IGF-1在腫瘤細胞中的表達與糖酵解過程中的信號通路存在密切相關性[20-21]。本實驗結果顯示,增強miR-451表達后肝癌細胞中mTOR、Ki-67、MMP-9、VEGF和IGF-1 mRNA表達量明顯降低,而AMPK mRNA表達量明顯增高;抑制miR-451表達后肝癌細胞中mTOR、Ki-67、MMP-9、VEGF和IGF-1 mRNA表達量增高,而AMPK mRNA表達量明顯降低。提示miR-451可能通過調控AMPK/mTOR傳導通路以及細胞中Ki-67、MMP-9、VEGF、IGF-1的表達,進而影響腫瘤細胞代謝途徑,最終達到抑制腫瘤生長增殖的作用。
綜上所述,miR-451參與調控肝癌細胞增殖糖酵解,其調控機制可能通過影響AMPK/mTOR信號通路,進而影響細胞中Ki-67、MMP-9、VEGF、IGF-1的表達而實現。未來可以針對腫瘤糖酵解相關miRNAs進行靶向性研究來干預腫瘤細胞能量代謝,進而為腫瘤的治療提供新的途徑和思路。
[1] Bensinger SJ,Christofk HR. New aspects of the Warburg effect in cancer cell biology[J]. Semin Cell Dev Biol,2012,23(4):352-361
[2] Ponisovskiy MR. Warburg effect mechanism as the target for theoretical substantiation of a new potential cancer treatment[J]. Crit Rev Eukaryot Gene Expr,2011,21(1):13-28
[3] Chiu M,Ottaviani L,Bianchi MG,et al. Towards a metabolic therapy of cancer?[J]. Acta Biomed,2012,83(3):168-176
[4] Huang Y,Shen XJ,Zou Q,et al. Biological functions of microRNAs[J]. Bioorg Khim,2010,36(6):747-752
[5] Liu X,Zhang A,Xiang J,et al. miR-451 acts as a suppressor of angiogenesis in hepatocellular carcinoma by targeting the IL-6R-STAT3 pathway[J]. Oncol Rep,2016,36(3):1385-1392
[6] Zhu S,Huang Y,Su X. Mir-451 correlates with prognosis of renal cell carcinoma patients and inhibits cellular proliferation of renal cell carcinoma[J]. Med Sci Monit,2016,22:183-190
[7] Suresh PS,Venkatesh T,Tsutsumi R. In silico analysis of polymorphisms in microRNAs that target genes affecting aerobic glycolysis[J]. Ann Transl Med,2016,4(4):69
[8] Wang J,Lei ZJ,Guo Y,et al. miRNA-regulated delivery of lincRNA-p21 suppresses beta-catenin signaling and tumorigenicity of colorectal cancer stem cells[J]. Oncotarget,2015,6(35):37852-37870
[9] Rao SG. Mitochondrial changes in cancer[J]. Handb Exp Pharmacol,2017,240:211-227
[10] Maximchik PV,Kulikov AV,Zhivotovsky BD,et al. Cellular energetics as a target for tumor cell elimination[J]. Biochemistry (Mosc),2016,81(2):65-79
[11] Jansson MD,Lund AH. MicroRNA and cancer[J]. Mol Oncol,2012,6(6):590-610
[12] Arora A,Singh S,Bhatt AN,et al. Interplay between metabolism and oncogenic process:Role of microRNAs[J]. Transl Oncogenomics,2015,7:11-27
[13] Chen B,Li H,Zeng X,et al. Roles of microRNA on cancer cell metabolism[J]. J Transl Med,2012,10:228
[14] Wang J,Zhao X,Shi J,et al. miR-451 suppresses bladder cancer cell migration and invasion via directly targeting c-Myc[J]. Oncol Rep,2016,36(4):2049-2058
[15] Zeng T,Peng L,Chao C,et al. miR-451 inhibits invasion and proliferation of bladder cancer by regulating EMT[J]. Int J Clin Exp Pathol,2014,7(11):7653-7662
[16] Godlewski J,Nowicki MO,Bronisz A,et al. MicroRNA-451 regulates LKB1/AMPK signaling and allows adaptation to metabolic stress in glioma cells[J]. Mol Cell,2010,37(5):620-632
[17] Sorokin DV,Scherbakov AM,Yakushina IA,et al. The mechanism of adaptation of breast cancer cells to hypoxia: Role of AMPK/mTOR signaling pathway[J]. Bull Exp Biol Med,2016,160(4):555-559
[18] Liu LL,Long ZJ,Wang LX,et al. Inhibition of mTOR pathway sensitizes acute myeloid leukemia cells to aurora inhibitors by suppression of glycolytic metabolism[J]. Mol Cancer Res,2013,11(11):1326-1336
[19] Masui K,Tanaka K,Akhavan D,et al. mTOR complex 2 controls glycolytic metabolism in glioblastoma through FoxO acetylation and upregulation of c-Myc[J]. Cell Metab,2013,18(5):726-739
[20] Yamashita S,Moroga T,Tokuishi K,et al. Ki-67 labeling index is associated with recurrence after segmentectomy under video-assisted thoracoscopic surgery in stage I non-small cell lung cancer[J]. Ann Thorac Cardiovasc Surg,2011,17(4):341-346
[21] Wang Q,Diao X,Sun J,et al. Regulation of VEGF,MMP-9 and metastasis by CXCR4 in a prostate cancer cell line[J]. Cell Biol Int,2011,35(9):897-904
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
海峽科技與產業(2016年3期)2016-05-17 04:32:12
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
中國當代醫藥(2015年16期)2015-03-01 02:03:11
