小黑麥不同品種劣變種子的胚培養(yǎng)效果研究
2018-03-27 09:50:25田新會(huì)杜文華
草原與草坪 2018年1期
關(guān)鍵詞:差異
李 雪,田新會(huì),杜文華
(甘肅農(nóng)業(yè)大學(xué) 草業(yè)學(xué)院/草業(yè)生態(tài)系統(tǒng)教育部重點(diǎn)實(shí)驗(yàn)室/甘肅省草業(yè)工程實(shí)驗(yàn)室/中-美草地畜牧業(yè)可持續(xù)發(fā)展研究中心,甘肅 蘭州 730070)
小黑麥(Triticalewittmack)為禾本科一年生草本植物,由小麥(Triticum)和黑麥(Secale)經(jīng)屬間有性雜交和雜種染色體加倍而成的新物種,不僅表現(xiàn)了小麥的豐產(chǎn)性和籽粒的優(yōu)良品質(zhì),又保持了黑麥抗逆性強(qiáng)和賴(lài)氨酸含量高的特點(diǎn),是一種很有前途的糧飼兼用型作物[1-3]。
種子的劣變是指因種子生存能力降低、導(dǎo)致種子喪失生活力和萌發(fā)力的不可逆轉(zhuǎn)變化,是伴隨著種子貯藏時(shí)間延長(zhǎng)而發(fā)生的、不可避免的過(guò)程[4]。在劣變過(guò)程中種子內(nèi)部發(fā)生一系列生理生化變化,種子的各種功能和結(jié)構(gòu)受到損害[5-7],其損害程度隨時(shí)間延長(zhǎng)而逐漸加劇,從而導(dǎo)致種子質(zhì)量下降,種子活力下降,甚至喪失生活力[8-9]。這主要是因?yàn)椋m然胚具有活力,但胚乳喪失了為胚萌發(fā)提供營(yíng)養(yǎng)物質(zhì)的能力,從而使胚由于缺乏營(yíng)養(yǎng)而失去活力[10]。
植物胚是一個(gè)具有全能性的多細(xì)胞結(jié)構(gòu),通常能夠正常發(fā)育成熟,而且可以直接播種形成完整植株。通過(guò)離體胚培養(yǎng),使可能敗育或退化的胚獲得再生植株,在植物育種中具有十分重要的理論與實(shí)踐意義[11-12]。
胚培養(yǎng)可以追溯到18 世紀(jì),Charles Bonnet分離并培養(yǎng)了菜豆和蕎麥的胚并獲得了植株[13-14]。較為系統(tǒng)的胚挽救技術(shù)開(kāi)始于20 世紀(jì)初,目前胚挽救技術(shù)已被廣泛地應(yīng)用于小麥[15]、大麥[16-17]、水稻[18]、茄子[19]、油菜[20]、百合[21]、以及眾多果樹(shù)如桃[22]、葡萄[23-24]、櫻桃[25]、蘋(píng)果[26]、柿[27]、李[28]、柑橘[29-30]等。Sirkka等最早進(jìn)行小黑麥胚挽救技術(shù)研究[31]。目前國(guó)內(nèi)外對(duì)小黑麥劣變種子胚挽救技術(shù)研究較少。為提高劣變種子的利用率和挽救種質(zhì)資源,試驗(yàn)以10份澳大利亞小黑麥品種的劣變種子為試材,在體外進(jìn)行胚培養(yǎng),以比較各品種的胚發(fā)育率、胚萌發(fā)率和成苗率,為小黑麥種質(zhì)資源挽救提供技術(shù)支持。
1 材料與方法
1.1 供試材料
供試材料為引自澳大利亞10個(gè)小黑麥品種(Cherokee,While M96-3182-1,33rd ITSN,DH265,AT315,Rhino,AT574,AT528,32rd ITSN,2090White)的劣變程度相同的種子。對(duì)照為甘肅農(nóng)業(yè)大學(xué)草業(yè)學(xué)院培育的小黑麥新品系C24(CK1)和C31(CK2),在室溫條件下保存一年的種子。
1.2 培養(yǎng)基
1/2MS+2%蔗糖+0.1%麥芽糖+0.5%瓊脂, pH 5.8,于121℃高壓滅菌20 min。
1.3 種子消毒及接種
先將參試材料的種子沖洗干凈,用次氯酸鈉消毒5~10 min,再用蒸餾水沖洗干凈。將沖洗干凈的種子浸泡在蒸餾水中,放置4℃冰箱24 h。
將浸泡的種子在無(wú)菌操作臺(tái)用次氯酸鈉消毒5~10 min,無(wú)菌水沖洗干凈并用無(wú)菌濾紙將種子干燥。用鑷子將種皮剝除,然后將胚從胚和胚乳連接處切出,完整的胚盾片向上接種到培養(yǎng)基中。
1.4 培養(yǎng)及轉(zhuǎn)接
將接種好的胚放置于17℃、光照16 h、黑暗8 h的培養(yǎng)箱中進(jìn)行培養(yǎng)。當(dāng)胚發(fā)芽至3片葉時(shí),轉(zhuǎn)移到花盆。
1.5 試驗(yàn)設(shè)計(jì)
將引自澳大利亞10個(gè)劣變小黑麥品種(Cherokee,While M96-3182-1,33rd ITSN,DH265,AT315,Rhino,AT574,AT528,32rd ITSN,2090White)和對(duì)照材料經(jīng)種子消毒及接種后在培養(yǎng)箱中培養(yǎng),每個(gè)處理設(shè)3次重復(fù),每天定時(shí)進(jìn)行觀察,每隔5 d統(tǒng)計(jì)小黑麥的胚發(fā)育率、成苗率,直至小黑麥的胚發(fā)育率、成苗率不在變化。
1.6 測(cè)定指標(biāo)及方法
接種的胚長(zhǎng)出綠芽或須根時(shí)為胚發(fā)育,胚發(fā)育率為發(fā)育胚數(shù)占接種胚珠數(shù)的百分率[32]。
胚發(fā)育率(%)=胚發(fā)育數(shù)/接種胚數(shù)×100%
成苗率為正常苗數(shù)占接種胚珠數(shù)的百分率。正常苗為具有正常的根、莖和葉等器官的植株[32]。
成苗率(%)=成苗數(shù)/接種胚數(shù)×100%
1.7 數(shù)據(jù)分析
用SPSS19.0軟件中二因素隨機(jī)區(qū)組試驗(yàn)設(shè)計(jì)的統(tǒng)計(jì)方法,分別分析不同小黑麥材料間、培養(yǎng)時(shí)間間和品種×培養(yǎng)時(shí)間交互作用間胚發(fā)育率、胚萌發(fā)率和成苗率方差的差異顯著性,進(jìn)行方差分析時(shí)將胚發(fā)育率和成苗率進(jìn)行數(shù)據(jù)轉(zhuǎn)換(反正弦轉(zhuǎn)換)。如果差異顯著,用Duncan法進(jìn)行多重比較。對(duì)小黑麥的胚發(fā)育率和成苗率進(jìn)行相關(guān)性分析。用Excel進(jìn)行數(shù)據(jù)整理和作圖。
2 結(jié)果與分析
小黑麥材料間,誘導(dǎo)培養(yǎng)天數(shù)間以及小黑麥材料間×誘導(dǎo)培養(yǎng)天數(shù)交互間的胚發(fā)育率和成苗率存在極顯著差異(P<0.01),需進(jìn)行多重比較(表1)。
2.1 胚發(fā)育率
2.1.1 小黑麥材料間胚發(fā)育率的差異 參試的12份小黑麥材料中,2個(gè)對(duì)照的胚發(fā)育率相近,顯著高于其余材料;Cherokee和DH265的胚發(fā)育率顯著高于其余參試材料;小黑麥材料2090White、AT528、While M96-3182-1、AT574和AT315的胚發(fā)育率依次降低,且有顯著差異;33rd ITSN和、32rd ITSN和Rhino的胚發(fā)育率為0(表2)。
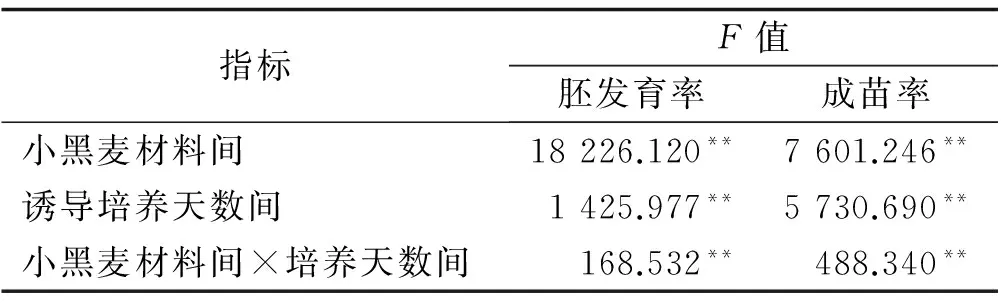
表1 小黑麥胚發(fā)育率和出苗率的方差結(jié)果分析
注:**達(dá)到極顯著水平(P<0.01),下同

表2 小黑麥材料間胚發(fā)育率和成苗率的差異
注:同列不同小寫(xiě)字母表示差異顯著(P<0.05),下同
2.1.2 培養(yǎng)天數(shù)間胚發(fā)育率的差異 由于小黑麥品種33rd ITSN、32rd ITSN和Rhino的胚發(fā)育率為0,所以從培養(yǎng)天數(shù)間胚發(fā)育率差異的影響中去除。隨著培養(yǎng)天數(shù)增加,小黑麥胚的發(fā)育率逐漸增加(表3)。0~7 d時(shí),小黑麥材料的胚發(fā)育率顯著低于其他培養(yǎng)天數(shù);7~12 d時(shí),胚發(fā)育率增加較快,增加率為25.4%;隨著誘導(dǎo)培養(yǎng)時(shí)間的繼續(xù)延長(zhǎng),胚發(fā)育率增加幅度減緩,27 d和32 d時(shí),胚發(fā)育率無(wú)顯著差異。
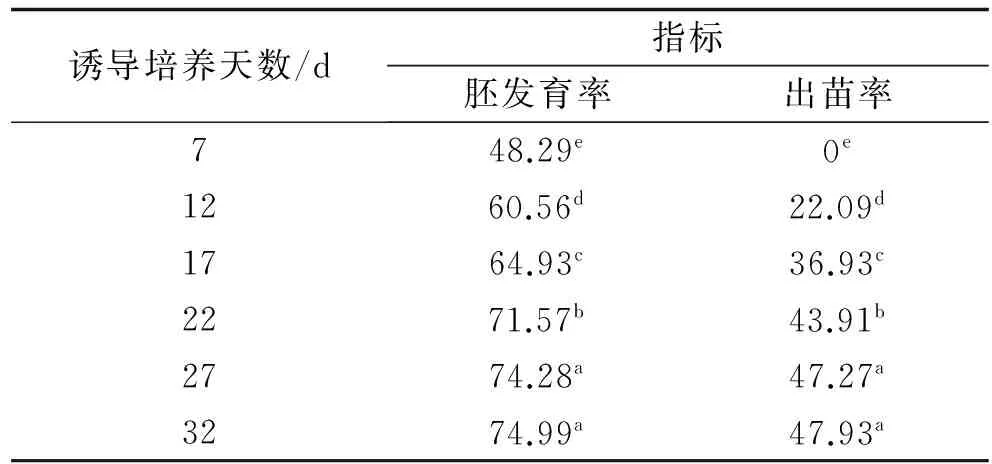
表3 誘導(dǎo)培養(yǎng)天數(shù)間胚發(fā)育率和成苗率的差異
2.1.3 小黑麥材料×培養(yǎng)天數(shù)交互作用間胚發(fā)育率的差異 CK1和CK2的胚發(fā)育率在誘導(dǎo)培養(yǎng)12 d時(shí)已經(jīng)完全發(fā)育,顯著高于其余小黑麥材料。隨著誘導(dǎo)培養(yǎng)天數(shù)增加,各參試小黑麥材料胚發(fā)育率的變化有差異,Cherokee誘導(dǎo)培養(yǎng)7 d的胚發(fā)育率較低,為57.65%,隨著誘導(dǎo)天數(shù)增加,胚發(fā)育率逐漸增加,27、32 d時(shí)胚發(fā)育率顯著增加;DH265誘導(dǎo)培養(yǎng)7 d的胚發(fā)育率較高(83.33%),胚發(fā)育率隨著誘導(dǎo)天數(shù)的增加而緩慢增加,22 d時(shí)胚發(fā)育率顯著增加,27,32 d時(shí)胚發(fā)育率還在增加,但無(wú)顯著差異;2090White誘導(dǎo)培養(yǎng)7 d時(shí),胚發(fā)育率較低(31.18%),12、17、22 d時(shí)胚發(fā)育率增加速度較快,差異顯著(P<0.05),27、32 d的增加緩慢,無(wú)顯著差異(P>0.05);AT528的胚發(fā)育率在誘導(dǎo)培養(yǎng)7 d時(shí)胚發(fā)育率為56.67%,12 d時(shí)胚發(fā)育率顯著增加,誘導(dǎo)培養(yǎng)12、17、22、27 d胚發(fā)育率增加緩慢,無(wú)顯著差異。AT315在誘導(dǎo)培養(yǎng)7 d時(shí),胚發(fā)育率較低(5%),12,17 d胚發(fā)育率顯著增加,17 d較12 d高118.95%,27,32 d胚發(fā)育率顯著增加;While M96-3182-1誘導(dǎo)培養(yǎng)7 d時(shí)胚發(fā)育為11.76%,12、17、22、27 d胚發(fā)育率增加,均差異顯著,27 d和35 d胚發(fā)育率無(wú)顯著差異。AT574誘導(dǎo)培養(yǎng)7 d時(shí)胚發(fā)育率為0,誘導(dǎo)培養(yǎng)12 d胚發(fā)育率為25.56%,誘導(dǎo)培養(yǎng)17,22和27 d胚發(fā)育率顯著增加,誘導(dǎo)培養(yǎng)35 d胚發(fā)育率增加緩慢,無(wú)顯著差異(圖1)。
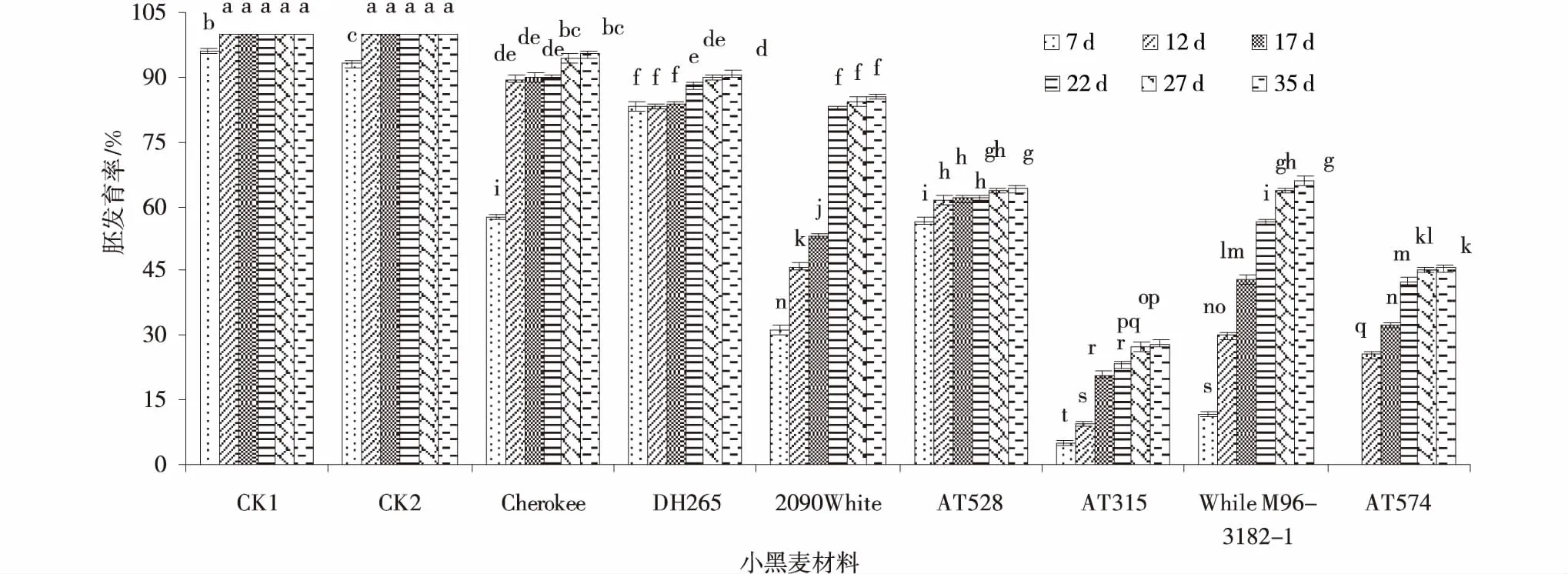
圖1 小黑麥材料×培養(yǎng)天數(shù)交互作用下的胚發(fā)育率Fig.1 Triticale materials×days of inducting culture of difference embryo development rate between interaction注:柱形圖間不同字母表示差異顯著(P<0.05)。下同
2.2 成苗率
2.2.1 小黑麥材料間成苗率的差異 對(duì)照材料的成苗率顯著(P<0.05)高于其余材料。參試小黑麥材料劣變種子的成苗率為8.23%~57.16%,Cherokee的成苗率最高,為57.16%,顯著高于其余參試小黑麥材料;DH265次之,其成苗率顯著高于除CK1、CK2和Cherokee之外的其他材料;2090White和AT528的胚成苗率相近,居第3位;其余材料的胚成苗率較低(表2)。
2.2.2 誘導(dǎo)培養(yǎng)天數(shù)間成苗率的差異 誘導(dǎo)培養(yǎng)7 d時(shí),小黑麥材料的成苗率為0。隨著誘導(dǎo)培養(yǎng)天數(shù)增加,成苗率逐漸增加,除27 d和32 d間小黑麥材料的成苗率無(wú)顯著差異外,其余天數(shù)間的胚成苗率均有顯著差異(P<0.05)。誘導(dǎo)培養(yǎng)17 d時(shí),小黑麥成苗率比12 d高67.18%,22 d的成苗率比17 d高18.90%,27 d的成苗率比22 d高7.65%;誘導(dǎo)培養(yǎng)時(shí)間27 d后,隨著培養(yǎng)時(shí)間延長(zhǎng),成苗率增加緩慢(表3)。
2.2.3 小黑麥材料×培養(yǎng)天數(shù)交互作用間成苗率的差異 CK1和CK2在誘導(dǎo)培養(yǎng)12 d時(shí)已經(jīng)全部成苗,其成苗率顯著(P<0.05)高于其余小黑麥材料。參試的小黑麥材料在誘導(dǎo)培養(yǎng)7 d時(shí)成苗率均為0。Cherokee誘導(dǎo)培養(yǎng)12 d時(shí)成苗率較低(6.47%),誘導(dǎo)培養(yǎng)17 d時(shí)成苗率顯著增加,但誘導(dǎo)培養(yǎng)22,27和32 d增加緩慢,差異不顯著;DH265在誘導(dǎo)培養(yǎng)12 d時(shí)成苗率為0,誘導(dǎo)培養(yǎng)17、22 d成苗率顯著增加,27、35 d成苗率增加緩慢,無(wú)顯著差異;2090White的成苗率在誘導(dǎo)培養(yǎng)12,17和22 d成苗率顯著增加,27、35 d成苗率增加緩慢,差異不顯著;AT528誘導(dǎo)培養(yǎng)12 d的成苗率較高(40.67%),17 d成苗率顯著增加,22,27和35 d的成苗率增加緩慢,差異不顯著;AT315在誘導(dǎo)培養(yǎng)12 d時(shí)成苗率為0,17、22 d成苗率顯著增加,27、35 d成苗率無(wú)差異顯著性;While M96-3182-1誘導(dǎo)培養(yǎng)12 d時(shí)成苗率為6.62%,誘導(dǎo)培養(yǎng)17 d成苗率顯著增加,高于241.24%,22 d時(shí)成苗率顯著增加,27、35 d成苗率增加差異不顯著;AT574誘導(dǎo)培養(yǎng)12 d、17 d成苗率為0,誘導(dǎo)培養(yǎng)22,27和35 d成苗率增加,但無(wú)差異顯著(圖2)。
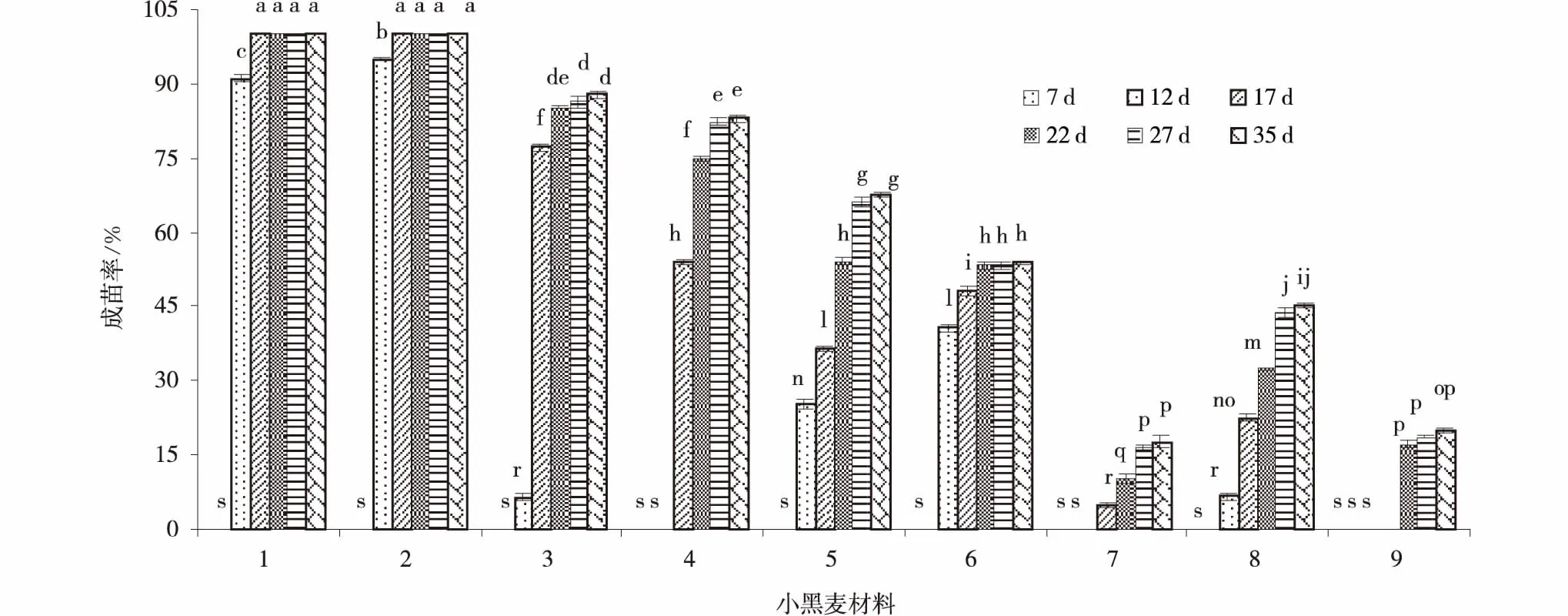
圖2 小黑麥材料×培養(yǎng)天數(shù)交互作用的成苗率Fig.2 Triticale materials×days of inducting culture of difference seedling rate among interaction
2.3 胚發(fā)育率和成苗率的相關(guān)性分析
小黑麥材料的胚發(fā)育率和成苗率相關(guān)性分析(表4)表明,小黑麥的胚發(fā)育率和成苗率呈極顯著正相關(guān)(R=0.97)。

表4 胚發(fā)育率和成苗率的相關(guān)性分析
2.4 再生苗移栽的幼苗成活率、株高和葉片數(shù)
對(duì)照CK1和CK2的成活率最高,顯著高于其他參試小黑麥材料。劣變的種子胚成活率為9.55%~69.42%,且不同品種差異顯著,其中Cherokee成活率最高,其次為DH265,AT315的成活率最低(表5)。
對(duì)照CK1和CK2的平均株高最高,顯著高于其余參試小黑麥材料。AT528平均株高較高,為16.40 cm,和CK2無(wú)顯著差異,其次為Cherokee,While和M96-3182-1,AT574的平均株高最低。
小黑麥品種AT528的平均葉片數(shù)最高,顯著高于其余參試小黑麥材料;其次為CK1、Cherokee和AT315,3品種的平均葉片數(shù)無(wú)顯著差異;AT574的平均葉片數(shù)最少,除與2090White無(wú)顯著差異外,顯著低于其余參試小黑麥。
3 討論
貯藏時(shí)間對(duì)種子的萌發(fā)具有較大的影響。張東暉等[33]研究表明,隨貯藏時(shí)間延長(zhǎng),種子劣變加劇,但有一定的安全貯藏期。貯藏14年喪失活力,貯藏7~8年失去種用價(jià)值,貯藏6年活力驟然下降,不宜作為播種用,貯藏4年保持較高的活力,適當(dāng)可作播種用,也可作為種質(zhì)資源保存期延長(zhǎng)至6年,貯藏2~3年活力表現(xiàn)最強(qiáng),說(shuō)明其種用價(jià)值最佳。試驗(yàn)選用不同程度劣變種子進(jìn)行研究,表明正常的種子胚的發(fā)育率和成苗率較高,并且由于劣變種子的劣變程度不同,其發(fā)育率和成苗率各異,發(fā)育率由高到低依次為:CK2>CK1>DH265>Cherokee>2090White>AT528>WhileM96-3182-1> AT754>AT315,成苗率依次為:CK2>CK1>Cherokee>DH265>2090White>AT528>WhileM96-3182-1> AT754>AT315。從而說(shuō)明DH265和Cherokee的劣變程度較輕,2090White和AT528居中,其余3個(gè)材料種子的劣變程度較重,而33rd ITSN,32rd ITSN和Rhino小黑麥種子已經(jīng)喪失了活力。和張東暉等[33]的研究結(jié)果類(lèi)似。不同植物種類(lèi)或品種胚培養(yǎng)存在較大差異。傅雪琳等[34]研究發(fā)現(xiàn),成熟稻材料間胚愈傷組織誘導(dǎo)率、質(zhì)地結(jié)構(gòu)和綠苗再生率有顯著差異。王丹菲等[35]用不同百合品種雜交獲得的幼胚為外植體,進(jìn)行離體胚培養(yǎng)發(fā)現(xiàn),不同雜交組合的胚發(fā)育率差異較大。試驗(yàn)材料DH265和Cherokee的劣變程度較輕,但2個(gè)材料間的胚發(fā)育率和成苗率還有顯著差異。由于成苗率和胚發(fā)育率極顯著正相關(guān),所以對(duì)其差異原因沒(méi)有單獨(dú)列出。
胚培養(yǎng)時(shí)間對(duì)胚挽救具有重要作用,張?jiān)虑俚萚36]研究表明,小麥成熟胚培養(yǎng)12 d就可以達(dá)到理想的分化效果,唐冬梅等[37]研究表明,無(wú)核葡萄不同基因型的胚培養(yǎng)時(shí)間不同。試驗(yàn)結(jié)果表明,隨著誘導(dǎo)天數(shù)增加,參試小黑麥材料的胚發(fā)育率逐漸增加。誘導(dǎo)培養(yǎng)7~12 d時(shí)胚發(fā)育率和成苗率增加幅度較大,誘導(dǎo)培養(yǎng)27 d和32 d時(shí)小黑麥材料的胚發(fā)育率和成苗率無(wú)顯著差異,說(shuō)明小黑麥胚培養(yǎng)的適宜誘導(dǎo)培養(yǎng)時(shí)間為27 d。
由小黑麥材料×誘導(dǎo)培養(yǎng)天數(shù)間的胚發(fā)育率和成苗率差異可知,CK1和CK2的胚發(fā)育率和成苗率在誘導(dǎo)培養(yǎng)12 d時(shí),均達(dá)100%,但劣變小黑麥材料種子在誘導(dǎo)培養(yǎng)27 d時(shí),胚發(fā)育率仍沒(méi)有達(dá)到100%,表明種子劣變對(duì)胚萌發(fā)有很大影響。參試的每種小黑麥材料的胚發(fā)育率和成苗率隨著時(shí)間的增加而增加,但增加的速度不同。材料Cherokee的胚發(fā)育率在7~12 d時(shí)增加較快,而2090White在17~22 d增加快,AT315和WhileM96-3182-1隨著時(shí)間的增加呈階梯式增加,其余材料的胚發(fā)育率顯著增加,但幅度不大。小黑麥材料2090White和AT528的成苗率在7-12 d時(shí)增加幅度較大,Cherokee和DH265在12-17d時(shí)增加快,其余材料增加勻速增加。趙占軍等[38]研究胚齡對(duì)小麥幼胚組織培養(yǎng)效果中表明,幼胚組織培養(yǎng)最適宜的胚齡為14~16 d。孫蘇陽(yáng)等[39]研究表明,小麥胚齡為16 d 時(shí),其萌發(fā)率和成苗率較高,是較為合適的幼胚培養(yǎng)時(shí)期。材料33rd ITSN,32rd ITSN,Rhino隨著誘導(dǎo)培養(yǎng)時(shí)間的增加而無(wú)變化,可能由于其種子劣變時(shí)間較長(zhǎng),而失去活力。
4 結(jié)論
小黑麥材料間胚發(fā)育率由高到低依次為:CK2>CK1>DH265>Cherokee>2090White>AT528> WhileM96-3182-1>AT754>AT315,成苗率的順序?yàn)椋篊K2>CK1>Cherokee>DH265>2090White>AT528> WhileM96-3182-1>AT754>AT315;小黑麥品種33rd ITSN、32rd ITSN和Rhino的胚發(fā)育率和成苗率均為0,完全失活;對(duì)劣變小黑麥的種子沒(méi)喪失活力的胚進(jìn)行培養(yǎng),最適宜的時(shí)間為27 d。
[1] 楊蕊菊.小黑麥抗旱生態(tài)適應(yīng)性研究[D].蘭州:甘肅農(nóng)業(yè)大學(xué),2003.
[2] 張微,李春艷,曹連莆,等.春性小黑麥材料抗旱性的比較研究[J].石河子大學(xué)學(xué)報(bào)(自然科學(xué)版),2009(5):529-535.
[3] 李冬梅,田新會(huì),杜文華.飼草型小黑麥新品系在甘肅臨洮灌區(qū)的生產(chǎn)性能研究[J].草原與草坪,2016(5):76-81.
[4] Anderson J D.Metabolic changes as sociated with senscence[J].Seed Science and Technology,1973,1:401-416.
[5] 董新紅,宋明.種子劣變的原因及其防止與修復(fù)[J].中國(guó)種業(yè),2002(1):39-40.
[6] 王彥榮,劉友良,沈益新.種子劣變的生理學(xué)研究進(jìn)展綜述[J].草地學(xué)報(bào),2001(3):159-164.
[7] 李青豐,易津,房麗寧,等.種子的劣變及劣變?cè)虻难芯縖J].內(nèi)蒙古農(nóng)牧學(xué)院學(xué)報(bào),1996(2):59-65.
[8] 韓建國(guó),浦心春,毛培勝.高羊茅種子老化過(guò)程中酶活性的變化[J].草地學(xué)報(bào),1998(2):84-89.
[9] 陳潤(rùn)政,周曉強(qiáng),傅家瑞.不同貯藏濕度對(duì)紅麻種子活力一些生理生化變化的影響[J].種子,1989(3):5-7.
[10] Bhaltacharrya S,Hazra A K,Sen Mandi S.Accelerated aging of seeds in hot water:germination characteristic-ccs of aged wheat seeds[J].Seed science and technology,1985,13(3):683-690.
[11] Gray D J,Fisher L C,Mortensen J A.Comparison of methodologies for inovule embryo rescue of seedless grapes[J].Hort Science,1987,22(6):1334-1335.
[12] Gray D J,Mortensen J A,Betten C.Ovule culture to obtain progeny fromhybrid seedless bunch grapes[J].J Amer Soc Hort Sci,1990,115(6):1019-1024.
[13] 陳香波,曹孜義.離體葡萄未成熟胚成苗途徑研究[J].果樹(shù)科學(xué),2000(4):261-264.
[14] 董曉玲.無(wú)核葡萄楊格爾的胚發(fā)育及胚珠培養(yǎng)[C]∥葡萄研究論文選集,2003(3):39-41.
[15] 鄧光兵,馬欣榮,張小萍,等.Phlb基因在普通小麥與簇毛麥雜種F1中的作用[J].應(yīng)用與環(huán)境生物學(xué)報(bào),1998(3):211-215.
[16] 李璋,劉翠云,閻正錄,等.普通小麥×栽培大麥雜種植株及其回交后代的產(chǎn)生和鑒定[J].遺傳學(xué)報(bào),1987(3):188-192.
[17] Xia G M,Xiang F N,Zhou A F.Asymmetric somatic hybridizationbetween wheat (TriticumaestivumL.)andAgropyronelongatum(Host)Nevishi[J].Theor Appl Genet,2003,107:299-305 .
[18] 遲全勃,Prapa Sripichitt,許紅春.通過(guò)胚胎拯救將野生稻耐旱特性轉(zhuǎn)移到栽培稻進(jìn)行遠(yuǎn)緣雜交初探[J].中國(guó)農(nóng)學(xué)通報(bào),2005(10):368-372.
[19] Buitendijk J H,Pinsonneaux N .Embryo rescue by half -ovule culturefor the production of interspecific hybrids in Alstroemeria[J].Scientia Horti culrurae,1995,64:65-75 .
[20] 王愛(ài)云,李栒,胡大有.生物技術(shù)在油菜種質(zhì)創(chuàng)新中的應(yīng)用[J].生命科學(xué)研究,2006(1):18-23.
[21] 楊利平,丁冰.百合種間遠(yuǎn)緣雜交種的育成[J].河北林果研究,1998(3):16-20.
[22] 莊恩及,姚強(qiáng),吳鈺良,等.早熟和特早熟桃胚珠培養(yǎng)研究[J].園藝學(xué)報(bào),1991(4):303-308.
[23] 潘春云,亓桂梅,湯小寧,等.葡萄三倍體育種初報(bào)[J].山東農(nóng)業(yè)大學(xué)學(xué)報(bào),1998(3):25-28.
[24] 徐海英,閻愛(ài)玲,張國(guó)軍.葡萄二倍體與四倍體材料間雜交胚珠的離體培養(yǎng)[J].果樹(shù)學(xué)報(bào),2001(6):317-320.
[25] Mu S K,Lena G.Rescue of hybrid embryos of Actinidia species[J].ScientiaHorticulturae,1990,44:97 -10 .
[26] Ramming D W.The use of embryos culture in fruit bre-eding[J].Hort-Science,1990,25(4):393-398 .
[27] 谷曉峰,唐仙英,羅正榮.羅田甜柿幼胚培養(yǎng)條件的研究[J].果樹(shù)學(xué)報(bào),2001(2):80-83.
[28] 劉用生,殷桂琴,汪濤.GA、6-BA、IBA與活性炭對(duì)李胚萌發(fā)及幼苗生長(zhǎng)影響[J].植物生理學(xué)通訊,1993(1):32-34.
[29] 裘文達(dá).園藝植物組織培養(yǎng)[M].上海:上海科技出版社,1986:195-197.
[30] 伊華林,鄧秀新.培養(yǎng)三倍體柑桔植株的研究[J].果樹(shù)科學(xué),1998(3):212-216.
[31] Fan Chuanzhu,Ma Yuansheng.Rescue and Characterization of Aged Seeds by Embryocultur-e[J].Biotechnology in Agriculture,1993(15):296-299.
[32] 王愛(ài)玲,王躍進(jìn),唐冬梅,等.提高無(wú)核葡萄胚挽救中幼胚成苗率的研究[J].中國(guó)農(nóng)業(yè)科學(xué),2010(20):4238-4245.
[33] 張東暉,云錦鳳,石鳳敏,等.不同貯藏時(shí)間披堿草種子劣變及活力測(cè)定[J].草業(yè)科學(xué),2008(4):116-118.
[34] 傅雪琳,盧永根,劉向東,等.幾個(gè)水稻粳型親秈系成熟胚離體培養(yǎng)研究初報(bào)[J].上海交通大學(xué)學(xué)報(bào)(農(nóng)業(yè)科學(xué)版),2006(3):230-234.
[35] 王丹菲,代漢萍,雷家軍.百合種間雜種胚離體培養(yǎng)研究[J].北方園藝,2006(4):157-159.
[36] 張?jiān)虑?陳耀鋒,王麗,等.小麥成熟胚培養(yǎng)條件的優(yōu)化及高效基因型篩選[J].干旱地區(qū)農(nóng)業(yè)研究,2014(5):100-105.
[37] 唐冬梅,蔡軍社,駱強(qiáng)偉,等.用于無(wú)核葡萄選育的胚挽救技術(shù)研究[J].果樹(shù)學(xué)報(bào),2008(3):316-321.
[38] 趙占軍,陳茂盛,王貴娟.胚齡和激素對(duì)小麥幼胚組織培養(yǎng)的影響[J].生物技術(shù),2003(5):7-8.
[39] 孫蘇陽(yáng),王永軍,李海軍,等.胚齡、激素對(duì)小麥單倍體萌發(fā)率及成苗率的影響[J].江蘇農(nóng)業(yè)學(xué)報(bào),2013(3):679-681.
猜你喜歡
英語(yǔ)世界(2023年10期)2023-11-17 09:19:16
汽車(chē)實(shí)用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂(lè)探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國(guó)特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國(guó)非營(yíng)利評(píng)論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語(yǔ)文(2016年21期)2016-05-25 13:13:50
