銅藻苗種繁育研究
2018-03-30 03:33:12王鐵桿鐘晨輝嚴興洪張立寧
水生生物學報 2018年2期
關鍵詞:生長
張 鵬 王鐵桿 鐘晨輝 嚴興洪 張立寧 劉 穎
(1. 浙江省海洋水產養殖研究所, 溫州 325005; 2. 浙江省近岸水域生物資源開發與保護重點實驗室, 溫州 325005;3. 福建省水產研究所, 廈門 361013; 4. 上海海洋大學水產與生命學院, 上海 201306)
銅藻[Sargassum horneri (Turn.) Ag.]隸屬于褐藻門(Phaeophyta), 墨角藻目(Fucales), 馬尾藻科(Sargassaceae Kuetzing), 馬尾藻屬(Sargassum)[1,2]。銅藻是北太平洋西部特有的暖溫帶性海藻, 不連續分布在我國沿海地區, 在浙江南麂島存在大量的銅藻海藻場, 俗稱為“丁香屋”, 因富含有益的生源要素而廣泛應用到醫藥、食品、飼料和有機肥料方面[3]。由于銅藻成體植株高大, 枝葉繁茂, 多生長在潮下帶淺海巖礁上, 成片漂浮水面, 堪稱“海中森林”, 為棲息在海藻場內的多種門類的海洋生物提供天然的避敵、索餌、產卵的場所。其次, 銅藻作為營造海藻場生境結構的關鍵藻種, 以其生長快速和生物量巨大獲得了海洋生態學家的關注與興趣,被列為重建海底藻場和修復近岸海洋生態系統的重要工具海藻之一[4—6]。
近年來, 由于陸源污染加劇了近海環境污染、銅藻自然繁衍場的海水養殖業和濱海旅游業的過度開發, 致使浙江省南麂島的野生銅藻自然資源量日趨減少, 天然銅藻場萎縮嚴重[7]; 另一方面, “漂浮銅藻”近年來在我國沿海的部分海域呈暴發性增長的趨勢, 對海上交通、海水養殖和生態環境帶來了嚴峻的挑戰[8]。為了保護好南麂島野生銅藻資源、實施退化藻場的生態恢復, 同時也為了有效應對“漂浮銅藻”帶來的危害, 了解環境因素與銅藻生長發育的關系, 開展銅藻繁殖生物學和苗種繁育等相關研究工作具有十分重要的意義。因此, 本研究以浙江省南麂島自然采集的野生銅藻作為親本, 結合銅藻繁殖生物學及海區養殖技術的研究和報道[9—12],研究了其生殖周期、受精卵發育模式, 并揭示了銅藻幼體早期發育與環境因子的關系等, 具有實際生產意義, 同時也為今后的銅藻增養殖、藻場生態修復、“漂浮銅藻”防治等提供了一定的參考。
1 材料與方法
1.1 育苗設施設備
育苗室內配備5個水泥池, 規格均為6 m×4 m×1.2 m (長×寬×深), 其中1個作為孢子收集池, 4個作為幼孢子體培育池。育苗室室溫可控, 通風順暢,安裝可調式布簾, 調節光照強和光照時間。配置用來收集銅藻受精卵的80目和200目網袋數個及用于清洗、播撒受精卵的塑料方桶、量筒、噴壺等釆苗器具。所用海水需經過暗沉淀和沙濾處理。
1.2 種藻挑選與保育促熟
2012年4月采自浙江省南麂島馬祖岙的野生銅藻成體(圖1)作為種藻(27°28′N, 121°03′E), 藻體規格: 株高(123.43±31.89) cm, 濕重(498.45±38.80) g,一級分支數為(115.88±16.98)個, 二級分支數(18±4.73)個, 雌雄比例控制在10∶1。采用黑暗沉淀的自然海水沖洗掉種藻表面的附著物后放置于水泥池內培養, 水深控制在1—1.5m, 鹽度24—28, 水溫15—20℃。每天施肥N濃度控制在10 mg/L, P濃度控制在1 mg/L, 促使藻體的成熟, 隔天更換全部海水并及時清理脫落的殘枝和污物。每天按時鏡檢成藻的雌雄生殖托發育情況, 當生殖托開始有大量黏液分泌, 且雌托有明顯的卵掛出時, 應當準備受精卵的收集。
1.3 受精卵收集方法

圖 1 銅藻成體Fig. 1 The Sargassum horneri (×10 cm)
觀察銅藻雌托掛卵發育情況, 當發現大部分卵已經受精且細胞開始分裂時, 進行銅藻受精卵的收集工作, 具體為: 分別將80目和200目網袋套在育苗池排水口上, 排放育苗池內的海水, 進行受精卵的收集, 同時用潛水泵(750 W)反復沖洗種藻, 加快受精卵從生殖托上脫落、放散。并不斷擠壓網袋, 排出內部的氣體, 加速小顆粒物質透過網袋。
當水中受精卵數量很少時, 應當停止受精卵的收集, 取下網袋, 用海水將80目網袋中的受精卵沖洗至200目網袋中, 然后將200目網袋中的受精卵轉移至50 L塑料方桶, 不斷加水攪拌、沉淀, 并倒掉方桶上層懸浮液, 反復多次至上清液澄清為止。收集方桶下層銅藻受精卵, 轉移至1000 mL玻璃量筒,沉淀2h定容, 以估算銅藻受精卵收集數量。銅藻受精卵放散量(C)的測算按如下公式計算:
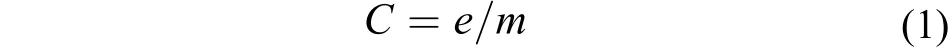
式(1)中, e為收集到的受精卵數, m為種藻的濕重。
1.4 受精卵發育觀察及溫度、鹽度試驗
在進行受精卵收集時, 應當對其發育情況進行實時觀察、拍照、記錄, 以了解胚胎發育情況, 以此作為受精卵收集、噴灑、移植的依據。
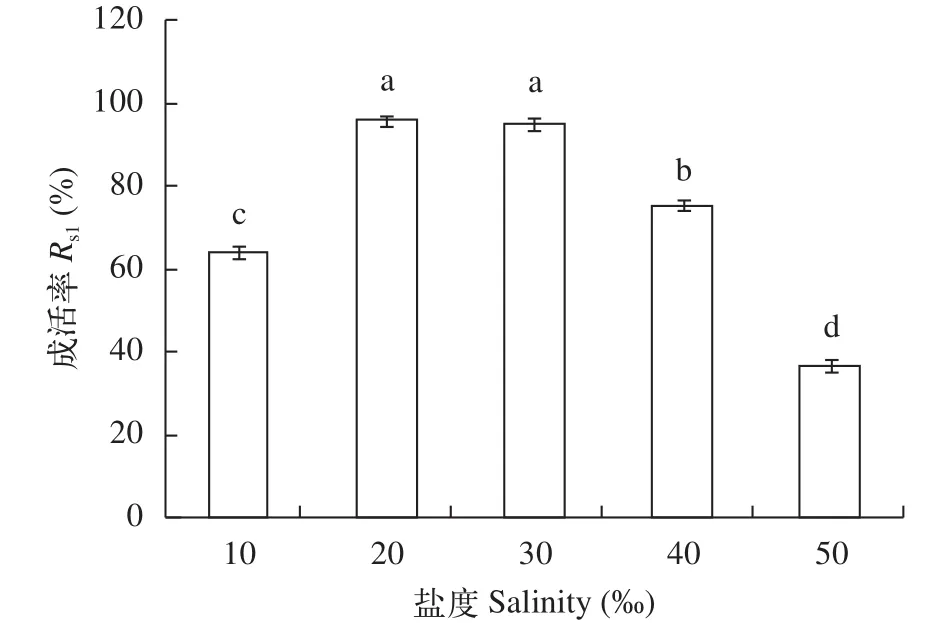
在1 L的燒杯中, 對銅藻胚胎發育進行鹽度、溫度試驗, 溫度設10、15、20、25和30℃等5組, 鹽度組分10、20、30、40和50等5組; 觀察各梯度下受精卵的成活率(試驗密度為8—10個/cm2), 試驗周期48h。試驗所用海水經過暗沉淀和二級沙濾處理,經煮沸冷卻后使用。使用的培養基為PESⅠ[13]。銅藻受精卵成活率(Rs1)按如下公式計算:
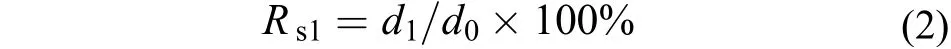
式(2)中, d0為試驗開始時的受精卵數, d1為試驗結束時已生成假根的銅藻幼孢子體個數。
1.5 幼孢子體培育
將量筒中測量好的銅藻受精卵依照培養密度要求倒入噴壺, 均勻噴灑至育苗池預先鋪設好的附著基上, 進行幼孢子體的培育。培育海水中加入N肥10 mg/L, P肥1 mg/L。培養密度控制在8—10個/cm2。
幼孢子體培育前期由于附著牢度不強, 應當盡量減少換水頻度和水流強度, 以促進銅藻幼孢子體的附著。待兩葉片期后, 銅藻幼孢子體附著逐漸牢固, 加大換水力度, 并根據育苗池中雜藻的生長情況,及時用水泵沖洗, 減少雜藻對幼孢子體生長的影響。觀察、測量和記錄幼孢子體生長、附著等情況。
1.6 幼孢子體培育的環境因子試驗
將銅藻受精卵接種至蓋玻片(24 mm×24 mm)上, 培養3d, 剔除死卵及多余的孢子, 密度控制在15個/cm2, 且保持均勻, 將蓋玻片轉移至500 mL的燒杯中, 進行環境因子試驗, 此時銅藻幼孢子體規格為(198.34±4.45) μm。環境因子試驗梯度按如下設定, 溫度設10、15、20、25和30℃等5組, 鹽度組設10、20、30、40和50等5組, 光照強度設500、1500、2500、3500、4500、5500、6500和7500 lx等8組, 光照周期設0L∶24D、6L∶18D、12L∶12D、18L∶6D、24L∶0D (L∶D, 明期∶ 暗期)等5種, 培養密度試驗設1、5、10、20、30和40棵/cm2等6組。除生態因子測試試驗外, 其他試驗的培養條件為(20±1)℃, 3500—4500 lx, 12L∶12D, 鹽度27—30,PESⅠ培養液。每個試驗設置3個重復組, 試驗周期為15d。所有試驗每3天更換1次培養液, 同時記錄孢子體生長發育情況。按如下公式計算幼孢子體的存活率(Rs2)和相對生長率(RGR)[14]:
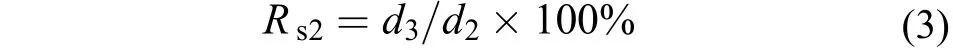
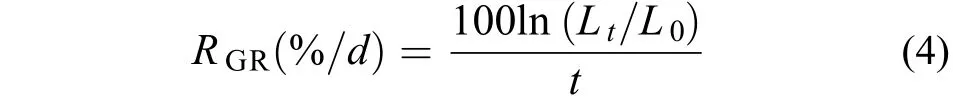
式(3)中, d2為實驗開始時的銅藻幼孢子數, d3為試驗結束時的幼孢子個數。式(4)中, L0為銅藻幼孢子體初始長度; Lt為試驗結束時銅藻幼孢子體長度;t為試驗周期。
1.7 幼孢子體附著基質選擇
為適應幼孢子體生長、后期養殖及銅藻場的重建等工作, 選擇扇貝殼、水泥磚、羊棲菜布簾、維尼龍繩簾、大理石磚等5種附著基, 進行銅藻附著基選擇試驗。觀察不同附著基對銅藻幼孢子體的附著率(RA)和相對生長率(RGR)的影響, 試驗周期為15d, 計算公式如下:
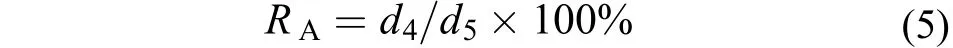
式(5)中, d5為試驗開始時的銅藻幼孢子數, d4為試驗結束時的幼孢子個數。相對生長率計算公式同公式(4)。
1.8 數據統計及分析
在試驗結束后, 利用Excel軟件將統計的不同處理組的Rs1、Rs2、RGR和RA等數據做成柱形圖, 并用SPSS19.0軟件進行Duncan’s多重比較, 以說明不同處理間的差異。
2 結果
2.1 繁殖習性初步觀察
根據觀察, 浙南海區銅藻每年繁殖一次, 自然繁殖盛期在每年的3月下旬至5月上旬, 此時海水溫度一般為13—18℃。銅藻為雌雄異體, 藻體分為主枝、側枝, 側枝上可再分出大量小枝, 雌、雄生殖托生于小枝上或葉腋間 (圖版Ⅰ-1、 2); 雌雄生殖托充分成熟時, 雌托呈短粗狀, 長度為14.3—46.0 mm (圖版Ⅰ-3、 5), 而雄托呈細長狀, 稍部略有彎曲, 長度為12.3—72.0 mm (圖版Ⅰ-4); 生殖托直徑從基部至頭部呈減小趨勢。成熟的生殖托重量可占整個藻體重量的20%—30%。
成熟雌性生殖托橫切后, 顯微鏡下觀察發現,生殖托內卵顆粒清晰, 核質均勻, 具有明顯的細胞膜; 縱切發現, 同一生殖托內卵具有發育不同步性,且稍部比中部和基部成熟晚(圖版Ⅰ-6)。生殖托即將排卵時, 表面出現大量黏液, 且有明顯的“拉絲”現象, 隨后卵子從生殖窩孔緩慢地逐個排出, 并黏附于生殖托表面, 等待受精(圖版Ⅰ-7)。在卵子受精后, 生殖托表面發育生成假根后幼孢子體隨即脫落, 進行附著生活。未受精卵子在生殖托上維持一段時間后, 脫落、死亡。
雄性生殖托成熟時, 精子以“精子團”的形式由生殖窩排出, 并黏附聚集在生殖窩外(圖版Ⅱ-8), 后“精子團”脫離生殖托, 迅速放散出單個的精子(圖版Ⅱ-9), 精子具有較強的游動能力。
2.2 受精卵的收集
受精卵定量將收集到的銅藻受精卵置于1000 mL玻璃量筒中, 沉淀2h, 體積不再變化時, 移除上清海水, 吸取沉淀好的受精卵沉淀混合液的上、中、下層各1 mL, 統計受精卵的含量(表 1), 從受精卵的數量來看, 以中層最高, 達到(5.31±0.37)×104個/mL, 其次是下層數量為(4.79±0.58)×104個/mL, 上層最低為(4.18±0.24)×104個/mL, 且差異顯著(P<0.05); 雜質數量下層最高, 中層次之, 上層最少,且差異顯著(P<0.05); 而受精卵比例方面上層最高,中層次之, 下層最少, 且差異顯著(P<0.05)。沉淀后的銅藻受精卵經測算含量為(4.76±0.54)×104個/mL。
收集結果一般受精卵應連續收集3—4d,以確保受精卵的足夠數量。2011—2013年銅藻育苗過程中受精卵的收集結果發現(圖 2), 受精卵的放散多集中在收集的中間2d, 雌性銅藻的受精卵收集量每天可以達到40—65 mL/kg, 經過4d左右的收集, 雌性銅藻受精卵的收獲量總共可以達到100—160 mL/kg, 可滿足大約50—90 m2的室內采苗需要。
2.3 受精卵的發育
發育過程剛釋放的受精卵無可見的凝聚核物質, 卵的長軸為(310.1±23) μm, 短軸為(245.7±18) μm, 呈橢圓型; 大約30min左右就會出現凝聚的核物質, 最終形成8個核(圖版Ⅱ-1、2); 卵子一旦受精, 卵內的多核在受精卵的中央融合成一個大核(圖版Ⅱ-3—8)。受精卵的第1次橫分裂發生在8核融合結束后, 形成2細胞的幼孢子體(圖版Ⅲ-1、2)。隨后, 在幼孢子體較窄一端的近頂端處, 進行第2次橫分裂, 形成“假根原細胞”(圖版Ⅲ-3、4)。然后, 孢子體經過多次橫縱分裂后, 形成多細胞的幼孢子體; 與此同時, 基部的“假根原細胞”也進行多次分裂, 最終形成假根芽 (圖版Ⅲ-5—7), 假根芽不斷發育形成假根(圖版Ⅲ-8)。受精卵經過48h左右繁育形成具有假根的幼孢子體, 在風浪、水流等外力作用下大量脫離生殖托, 開始獨立生活。
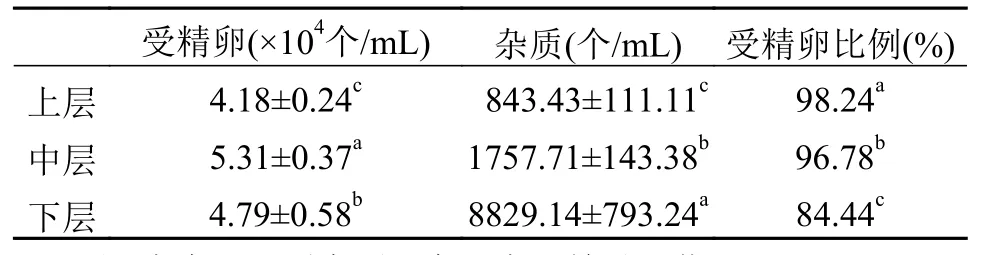
表 1 不同取樣位置受精卵的含量Tab. 1 Amounts of zygotes at different sampling positions
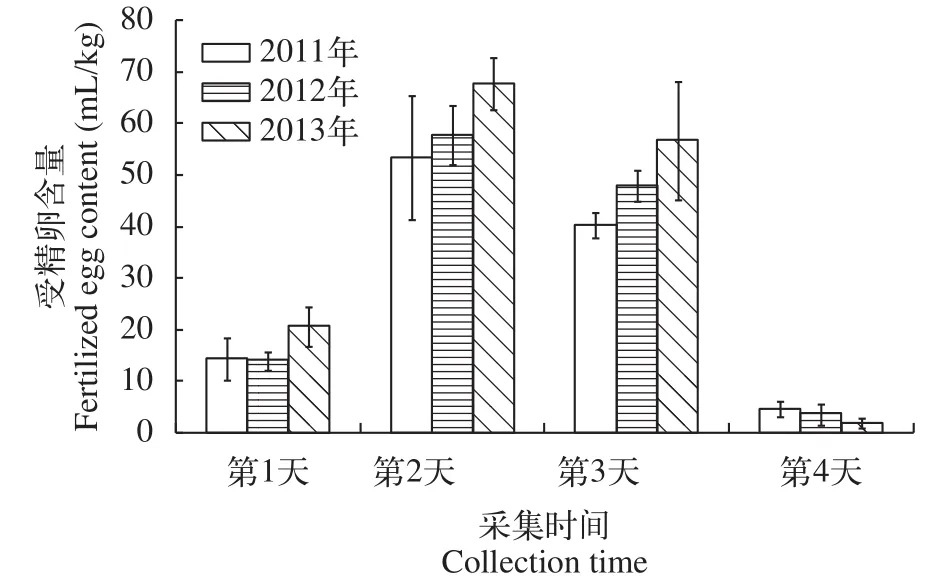
圖 2 2011—2013年銅藻受精卵收集情況Fig. 2 Collection of zygotes from 2011 to 2013
受精卵發育對溫度的要求如圖 3所示, 在10—20℃, 銅藻受精卵的成活率受溫度影響不大,不同溫度下的成活率差異不顯著(P>0.05); 當溫度超過20℃時, 隨著溫度的升高, 受精卵的成活率隨著溫度的升高而呈現下降趨勢, 且25℃和30℃的成活率明顯低于10℃至20℃(P<0.05), 因此, 10—20℃是銅藻受精卵發育的適宜溫度。
受精卵發育對鹽度的要求如圖 4所示, 鹽度10、40時, 受精卵發育緩慢, 成活率明顯低于鹽度20和30組(P<0.05), 鹽度為50的試驗組受精卵出現了大量死亡現象, 成活率為最低。由此可知, 銅藻受精卵發育適宜的鹽度范圍是20—30。
2.4 幼孢子體的發育
銅藻完成受精并形成假根之后(圖版Ⅳ-1), 幼孢子體生長加快, 外部形態也發生明顯變化。適宜環境下, 培養2—3d, 幼孢子體的頭部發育呈突起狀,長度達到(375±21) μm (圖版Ⅳ-2), 此時假根部更加發達, 假根長度和數量也不斷增加; 幼孢子體前期(1—4d)發育較慢, 日增長35—40 μm; 之后(5—8d)生長速度加快, 日增長達到144—170 μm, 孢子體頂端漸呈扁平狀, 此時長度達到(984±38) μm(圖版Ⅳ-3); 培育10—11d, 第一葉片基本形成, 生長放緩, 此時基部350—500 μm處逐漸形成小突起, 第二葉片開始發育, 此時長度達到1.2 mm左右(圖版Ⅳ-4); 第二葉片不斷發育伸長, 同時第一葉片伸長的過程中不斷變寬, 且邊緣呈一定的波浪形(圖版Ⅳ-5); 經過18—20d的發育, 第二葉片發育至第一葉片大約2/3長度時, 葉腋處開始出現第三葉片的突起,此時第一葉片基本停止伸長, 繼續變寬, 第二葉片伸長的過程中重復第一葉片的發育情況, 以此形成幼孢子體的前期發育模式(圖版Ⅳ-6、7)。
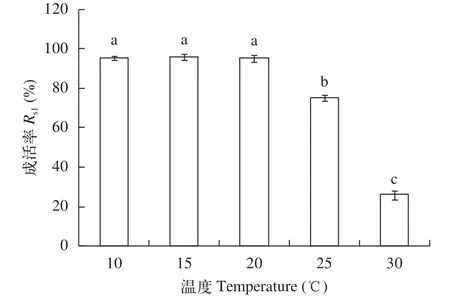
圖 3 溫度對銅藻受精卵成活率的影響Fig. 3 The effect of temperature on zygote survival
幼孢子體對溫度的要求如圖 5所示, 銅藻幼孢子體對溫度的適應范圍較廣, 10—25℃都可以存活, 且以10—20℃存活率較高, 明顯大于其他試驗組(P<0.05), 溫度升高至25℃時, 存活率和生長率呈明顯下降趨勢, 溫度30℃時, 幼孢子體死亡率較高且基本停止生長。因此, 溫度20℃最適合銅藻幼孢子體生長。
幼孢子體對鹽度的要求如圖 6所示, 鹽度20—30內銅藻幼孢子體存活率明顯高于其他組(P<0.05), 而生長率以20—30最為適宜(P<0.05), 過高和過低的鹽度都不利于銅藻幼孢子體的生長。
幼孢子體對光照強度的要求如圖 7所示,光強500—8500 lx內銅藻幼孢子體存活率明顯高于其他組(P<0.05), 而生長率以4500 lx最為適宜(P<0.05), 在黑暗條件下幼孢子體雖能保持一定的存活率(試驗周期內), 但基本停止生長, 當光照強度超過8500 lx時, 幼孢子體的存活率和生長率都呈明顯的下降趨勢(P<0.05)。這說明4500 lx的光照條件對銅藻幼孢子體生長較為適宜。
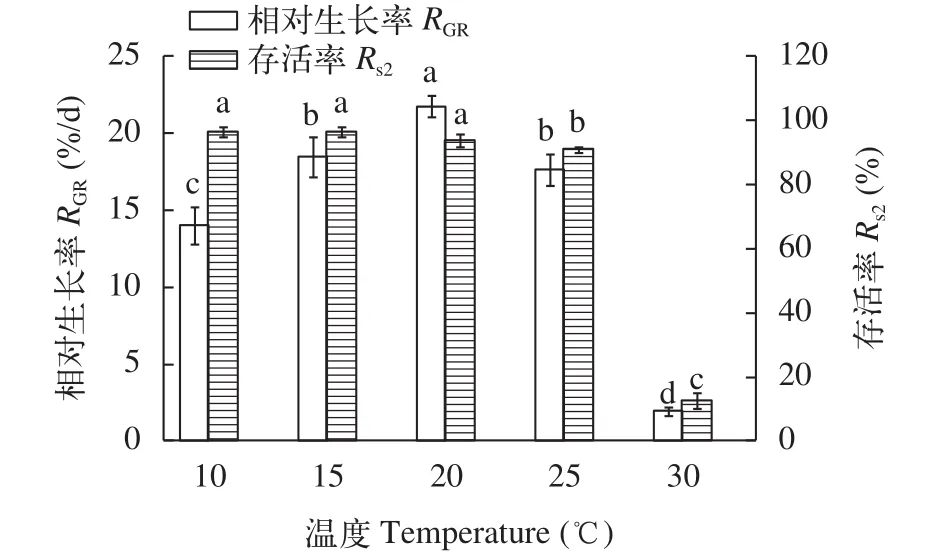
圖 5 溫度對幼孢子體生長和存活的影響Fig. 5 The effect of water temperature on germlings survival and growth
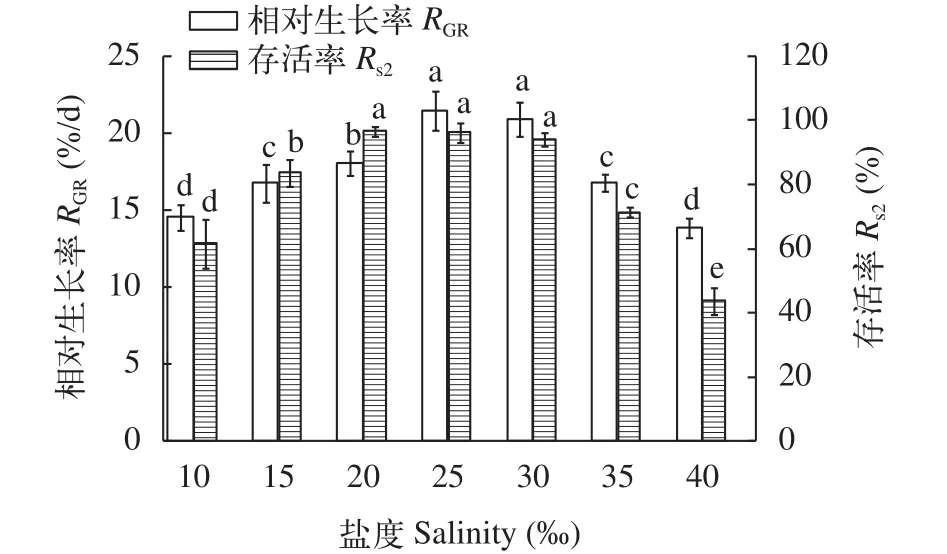
圖 6 鹽度對銅藻幼孢子體生長和存活的影響Fig. 6 The effect of salinity on germlings survival and growth
幼孢子體對光照周期的要求銅藻幼孢子在不同的光照周期下均能存活(圖 8), 在每天光照時間不低于6h下, 銅藻幼孢子體均能保持較高的存活率(P<0.05)。當全黑暗時, 銅藻幼孢子基本停止生長, 且存活率較低(P<0.05)。每天光照時間12—18h, 銅藻幼孢子的相對生長率較高(P<0.05), 比較適合其生長。

圖 7 光照強度對幼孢子體生長和存活的影響Fig. 7 The effect of light intensities on germlings survival and growth
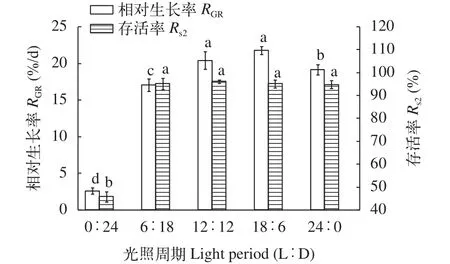
圖 8 光照周期對幼孢子體生長和存活的影響Fig. 8 The effect of light period on germlings survival and growth
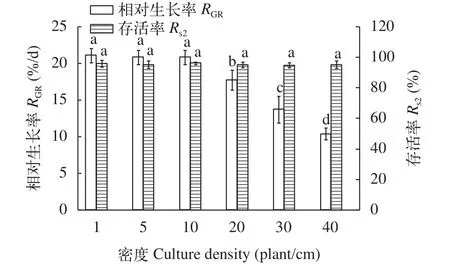
圖 9 培養密度對幼孢子體生長和存活的影響Fig. 9 The effect of culture density on germlings survival and growth
幼孢子體對培養密度的要求如圖 9所示,培養密度為1—40棵/cm2時, 幼孢子體均能保持較高的存活率且無明顯差異(P>0.05)。在培養密度1—10棵/cm2時, 幼孢子生長率相對較高(P<0.05),在培養密度超過10棵/cm2時, 幼孢子生長明顯受到影響(P<0.05), 且隨著密度的增加生長率呈現下降趨勢。因此, 幼孢子體培育階段培養密度應當控制在10棵/cm2內。
幼孢子體對附著基的要求如圖 10所示, 經過15d的培育, 扇貝殼和水泥磚附著基的附著率較高, 附著率超過70%, 其次為維尼龍繩簾, 附著率為65.1%, 羊棲菜布簾和大理石磚為最低(P<0.05)。而相對生長率方面, 水泥磚和維尼龍繩簾較高, 羊棲菜布簾和大理石磚次之, 扇貝殼最低(P<0.05)。因此, 質地堅硬表面粗糙的水泥磚較適合銅藻幼孢子體的的附著和生長。不同附著基效果如圖版Ⅴ所示。
3 討論
3.1 馬尾藻科育苗的研究比較
馬尾藻作為我國一類重要的大型藻類, 其主要養殖的種類有羊棲菜[15]、鼠尾藻[16,17]等; 其他種類如莫式馬尾藻(Sargassum maclurei)[18]瓦式馬尾藻(Sargassum vachellianum)[19]、海黍子(Sargassum muticum)[20]、半葉馬尾藻[21]、匍枝馬尾藻[22]等也有相關的人工育苗研究。比較發現, 雖然銅藻成體在外形特征上與其他馬尾藻類有所不同, 但是就其繁殖生物學而言, 差異不大, 受精卵到幼孢子體的各期形態也基本一致。
3.2 銅藻種藻的選擇、暫養和促熟排放
銅藻的生殖托發育與大多數馬尾藻類的生殖托發育過程較為相似, 雌性生殖托成熟后大量卵懸掛于雌性生殖托表面, 等待受精, 精子從成熟的雄性生殖托排出后, 游動到雌性生殖托表面, 與卵細胞結合形成受精卵, 然后從雌性生殖托表面脫落完成受精過程[15—22]。觀察發現, 南麂島銅藻生殖托有較為集中的形成時間, 一般為上半年的3月下旬至5月上旬。另外, 相同個體不同部位的生殖托形成時間和發育情況亦有不同, 野生型為頂部較先形成生殖托, 而養殖型靠近夾苗繩子的基部生殖托形成較早, 說明靠近水面的較高溫度、較強光照和較長光照時間等都是促進銅藻成體性成熟的關鍵因素所在。暫養期間應當加大換水量和換水次數, 制造水流以加快銅藻的成熟速度。由于成體生殖托形成的不同步性, 以及同一個生殖托內不同部位生殖細胞發育的不同步性等, 因此, 在選擇種藻的時候應當以較為成熟的側枝為選取對象, 人為控制其集中排放, 集中采苗。另外, 在銅藻促熟方面也應當加強類似于其他馬尾藻有性生殖同步化的相關研
究[23, 24]。
3.3 銅藻受精卵發育及其環境選擇
鼠尾藻、羊棲菜、莫氏馬尾藻、瓦氏馬尾藻、海黍子以及其他馬尾藻相比, 銅藻的生殖托最長[1]。而且銅藻受精卵也存在馬尾藻“掛托分裂”的現象, 卵子由雌性生殖托排放后先黏附在生殖托表面, 當完成受精后繼續在托上進行分裂直至長出假根, 隨后自然脫落。該結果與張婧等[25]有關枸杞島銅藻受精卵的早期發生發育觀察相類似。南麂島銅藻的卵發育方式屬于8核1卵型[26], 與其他馬尾藻的卵型發育方式類似[16—20]。溫度10—20℃, 鹽度20—30是銅藻受精卵發育的適宜環境。
3.4 銅藻幼孢子體培育關鍵
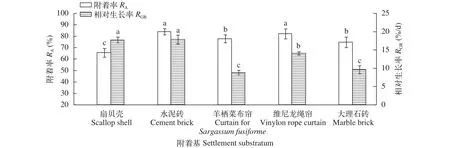
圖 10 附著基對銅藻幼孢子體生長和附著的影響Fig. 10 The effect of different settlement substratum on germlings survival and growth
環境因子本研究發現, 銅藻幼孢子體階段,有較廣的環境適應范圍。溫度方面, 在10—25℃條件下幼孢子體均能存活并能保證一定的生長率。該結論與孫建璋[5,11]、張婧等[25]通過對南麂列島的銅藻生態學實地調查以及枸杞島銅藻幼孢子體室內培育試驗, 得出銅藻的生存水溫范圍較廣(8—30℃), 在受精卵只發育成無主干和側枝的幼孢子體的階段, 可以耐受惡劣環境因子的結論相類似。而本研究通過統計分析, 得出的幼孢子體最適宜生長溫度為20℃。光照方面, 光強500—8500 lx銅藻幼孢子體存活率較高, 在黑暗條件下幼孢子體雖能保持一定的存活率, 但基本停止生長, 當光照強度超過8500 lx時, 幼孢子體的存活率和生長率都呈明顯的下降趨勢(P<0.05)。該結論與孫建璋等[5,11]認為銅藻合子附著后的幼孢子體對強光的耐受力下降的結論類似, 張婧等[25]也發現, 日齡小于30d的幼孢子體對強光耐受力較差, 且苗體頂端分生組織較易潰爛, 因此該階段銅藻幼孢子體的培養, 宜采用4500 lx左右的光照強度。
有關鹽度、光照周期、培養密度等環境因子對銅藻幼孢子體生長存活影響的相關研究報道較少, 因此, 本研究得出的銅藻幼孢子體適宜鹽度范圍、光照周期、培養密度等對銅藻的前期培育和人工育苗等工作有比較現實的參考價值。
附著基質銅藻作為一種較為高等的大型海藻, 已經分化出類似于高等植物根的“假根”附著器, 附著器是保證其抵抗風浪、選擇棲息環境、營附著生活的重要器官[2]。而影響銅藻營附著生活的最重要因素, 就是附著基。因此, 附著基的選擇是決定銅藻人工培育的關鍵所在。研究發現, 不同類型的孢子適宜附著和生長的附著基也不同[26,27]。韓軍軍[19]、孫修濤[28]、賈檉[29]等都進行過有關馬尾藻的附著基選擇的相關研究, 發現水泥板和棕繩較適合馬尾藻幼苗的生長發育。而本研究也有相同發現, 在室內培育條件下, 質地堅硬表面粗糙的水泥磚比較適合銅藻幼孢子體生長發育。且該附著基易于搬運、便于沖洗, 更適合后期海區藻場重建的藻礁投放等需要。
4 結論
綜上所述, 南麂島銅藻的自然繁殖盛期為每年3月下旬至5月上旬, 海水溫度為13—18℃, 較適宜于雌雄生殖托的形成。
在人工條件下, 有利于銅藻受精卵發育的適宜溫度為10—20℃, 鹽度是20—30, 在該培育條件下,經過48h的孵化, 受精卵可以大批量同步化發育成具有假根的幼孢子體。
人工育苗結果表明, 適宜的環境因子(溫度20℃, 鹽度20—30, 光照強度4500 lx, 光照時間12—18h/d)較利于幼孢子體的生長發育。銅藻幼孢子體的前期培育連續流水和充氣為宜, 經18—20d的室內培育, 幼孢子體可發育成具有2—3葉片的幼苗。另外, 銅藻幼孢子體的培養密度應當控制在10棵/cm2范圍內, 質地堅硬、表面粗糙的水泥磚較適合銅藻幼孢子體的附著和生長, 可滿足后期藻場修復的要求。
參考文獻:
[1]Zeng C K, Lu B R. Chinese Journal of Seaweed [M].Beijing: Science Press. 2000, 96—97 [曾呈奎, 陸保仁.中國海藻志, 第三卷第二冊, 墨角藻目. 北京: 科技出版社. 2000, 96—97]
[2]Qian S B, Liu D Y, Sun J. Haizaoxue.[M]. Qingdao:Press of Ocean University of China. 2005, 2—6 [錢樹本,劉東燕, 孫軍. 海藻學. 青島: 中國海洋大學出版社.2005, 2—6]
[3]Zhang P, Cai Y F, Wang T G, et al. AFLP analysis of different geographic populations of Sargassum horneri along the coast of Zhejiang Province [J]. Acta Agriculturae Zhejiangensis, 2015, 27(9): 1586—1592 [張鵬, 蔡一凡, 王鐵桿, 等. 浙江沿海不同地理群體銅藻Sargassum horneri的AFLP分析. 浙江農業學報, 2015, 27(9):1586—1592]
[4]Choi C G, Kim H G, Sohn C Y. Transplantation of young fronds of Sargassum horneri for construction of seaweeds beds [J]. Journal of the Korean Fisheries Society,2003, 36(5): 469—473
[5]Sun J Z, Chen W D, Zhuang D G, et al. In situ ecological studies of the subtidal brown alga Sargasssum horneri at Nanji Island of China [J]. South China Fisheries Science,2008, 4(3): 58—63 [孫建璋, 陳萬東, 莊定根, 等. 中國南麂列島銅藻Sargassum horneri 實地生態學的初步研究.南方水產, 2008, 4(3): 58—63]
[6]Chen L R. Morphological traits of Sargassum horneri and its influences on habitat structure of the seaweed bed [D].Thesis for Master of Science. Shanghai Ocean University,Shanghai. 2015 [陳亮然. 銅藻形態學特征及其對海藻場生境構造的影響. 碩士學位論文, 上海海洋大學, 上海.2015]
[7]Zheng H Y, Rao D Z, Chen G F, et al. Consideration on protective exploitation of Sargassum horneri (Turn) Ag resource around Nanji Islands [J]. Modern Fisheries Information, 2008, 23(10): 25—27 [鄭海羽, 饒道專, 陳高峰, 等. 保護性開發南麂列島 Sargassum horneri (Turn)Ag. 資源的思考. 現代漁業信息, 2008, 23(10): 25—27]
[8]Lin S M, Huang R, Ogawa H, et al. Assessment of germling ability of the introduced marine brown alga,Sargassum horneri in Northern Taiwan [J]. Journal of Applied Phycology, published online: 11 March 2017
[9]Isamu Umezaki. Ecological Studies of Sargassum horneri(TURNER) C. AGARDH in Obama Bay, Japan Sea [J].Bulletin of the Japanese Society of Scientific Fisheries,1984, 50(7): 1193—1200
[10]Nanba N. Egg release and germling development in Sar-gassum horneri (Fucales, Phaeophyceae) [J]. Phycological Research, 1995, 43: 121—125
[11]Sun J Z, Zhuang D G, Chen W D, et al. Studies on sexual reproduction and seedling production of the brown alga Sargassum horneri [J]. South China Fisheries Science,2008, 4(2): 6—14 [孫建璋, 莊定根, 陳萬東, 等. 銅藻Sargassum horneri繁殖生物學及種苗培育的研究. 南方水產, 2008, 4(2): 6—14]
[12]Sun J Z, Zhuang D G, Sun Q H, et al. Artificial cultivation trials of Sargassum horneri at Nanji islands of China[J]. South China Fisheries Science, 2009, 5(6): 41—46[孫建璋, 莊定根, 孫慶海, 等. 銅藻人工栽培初步研究.南方水產, 2009, 5(6): 41—46]
[13]Wang S J. Marine Algal Biotechnology [M]. Shanghai:Shanghai Science and Technology Press. 1994, 20—21[王素娟. 海藻生物技術. 上海: 上海科學技術出版社.1994, 20—21]
[14]Hunt R. Plant Growth Analysis [M]. London: Edward Arnord. 1978, 2
[15]Li S Y. Studies on technique of industrial raising seedlings of Sargassum fusiforme (Harvey) setch [J]. Journal of Zhejiang Ocean University (Natural Science), 2001,20(3): 251—265 [李生堯. 羊棲菜生產性育苗技術研究.浙江海洋學院學報(自然科學版), 2001, 20(3):251—265]
[16]Wang F J, Sun X T, Li F. Studies on sexual reproduction and seedling-rearing of Sargassum thunbergii [J]. Marine Fisheries Research, 2006, 27(5): 1—6 [王飛久, 孫修濤,李鋒. 鼠尾藻的有性繁殖過程和幼苗培育技術研究. 海洋水產研究, 2006, 27(5): 1—6]
[17]Wang Z F, Liu J G. The sexual reproduction of Sargassum thunbergii and its application in seedling breeding[J]. Oceanologia Et Limnologia Sinica, 2007, 38(5):453—457 [王增福, 劉建國. 鼠尾藻(Sargassum thunbergii)有性生殖過程與育苗. 海洋與湖沼, 2007, 38(5):453—457]
[18]Yang B, Qu Y K, Xie E Y. Morphological observation of Sargassum maclurei sexual reproduction and juvenile sporophyte development [J]. Journal of Aquaculture,2013, 34(10): 30—34 [楊彬, 曲元凱, 謝恩義. 莫氏馬尾藻有性繁殖和幼孢子體發育的形態學觀察. 水產養殖,2013, 34(10): 30—34]
[19]Han J J, Zhang J W, Liu Y Y, et al. Studies on the largescale breeding of Sargassum vachellianum [J]. Acta Hydrobiologica Sinica, 2016, 40(1): 123—130 [韓軍軍, 張建偉, 劉媛媛, 等. 瓦氏馬尾藻規模化繁育技術研究. 水生生物學報, 2016, 40(1): 123—130]
[20]Cao S Q, Zhang Z Y, Wang G S, et al. Indoor artificial seeding of seaweed Sargassum muticum [J]. Journal of Dalian Fisheries University, 2008, 23(5): 359—364 [曹淑青, 張澤宇. 海黍子室內人工育苗技術的研究. 大連水產學院學報, 2008, 23(5): 359—364]
[21]Tian S X, Chen W D, Lin L, et al. Morphological observation and structural analysis of Sargassum hemiphyllum(Turn.) var. chinense J. Agardh in breeding season [J].Marine Fisheries, 2014, 36(2): 107—115 [田淑嫻, 陳萬東, 林利, 等. 繁殖期半葉馬尾藻中國變種的形態結構觀察. 海洋漁業, 2014, 36(2): 107—115]
[22]Zou X X, Lin Y, Zhu J, et al. Study on the artificial cultivation technology of Sargassum polycystum [J]. Fishery Modernization, 2016, 43(3): 29—33 [鄒瀟瀟, 林勇, 朱軍, 等. 匍枝馬尾藻人工栽培初步研究. 漁業現代化,2016, 43(3): 29—33]
[23]Pang S J, Fei X G, Xiao T, et al. Mass production of the artificial seeds of Hizikia fusiformis by controlling the laying out of the ovules and semens [J]. Marine Sciences,2001, 25(4): 53—54 [逄少軍, 費修綆, 肖天. 通過控制卵子和精子的排放實現羊棲菜人工種苗的規模化生產.海洋科學, 2001, 25(4): 53—54]
[24]Han J J, Bai K Q, Zhao M L, et al. Morphological observation of Sargassum vachellianum sexual reproduction and juvenile sporophyte development [J]. Acta Hydrobiologica Sinica, 2016, 40(6): 1215—1220 [韓軍軍, 白凱強,趙明林, 等. 瓦氏馬尾藻有性繁殖和幼孢子體發育的形態學觀察. 水生生物學報, 2016, 40(6): 1215—1220]
[25]Zhang J, Yan X H, Zhang S Y. Observations on embryology of zygotes and early development of juvenile sporophytes in Sargassum horneri [J]. Journal of Fisheries of China, 2012, 36(11): 1706—1716 [張婧, 嚴興洪, 章守宇. 銅藻受精卵的早期發生與幼孢子體發育觀察. 水產學報, 2012, 36(11): 1706—1716]
[26]Han J J, Zhang J W, Liu Y Y, et al. Studies on the largescale breeding of Sargassum vachellianum [J]. Acta Hydrobiologica Sinica, 2016, 40(1): 123—130 [韓軍軍, 張建偉, 劉媛媛, 等. 瓦氏馬尾藻規模化繁育技術研究. 水生生物學報, 2016, 40(1): 123—130]
[27]Zhang Z Y, Han Y X, Li X G, et al. Reproductive biology and indoor cultivation of seaweed Gloiopeltis tenax[J]. Journal of Dalian Fisheries University, 2009, 24(1):1—7 [張澤宇, 韓余香, 李獻剛, 等. 鹿角海蘿的繁殖生物學及室內培養的研究. 大連水產學院學報, 2009,24(1): 1—7]
[28]Sun X T, Wang F J, Wang W J, et al. Large scale breeding of Sargassum thunbergii based on sexual reproduction [J]. Progress in Fishery Sciences, 2010, 31(3):84—91 [孫修濤, 王飛久, 汪文俊, 等. 基于有性繁殖的鼠尾藻規模化繁育試驗. 漁業科學進展, 2010, 31(3):84—91]
[29]Jia C, Yang B, Xie E Y. Reproductive biology of seaweed Sargassum moclurei [J]. Fisherie Science, 2012,31(10): 616—619 [賈檉, 楊彬, 謝恩義. 莫氏馬尾藻繁殖生物學初步研究. 水產科學, 2012, 31(10): 616—619]

圖版 Ⅰ 銅藻生殖托發育Plate Ⅰ The development of Sargassum horneri receptacles
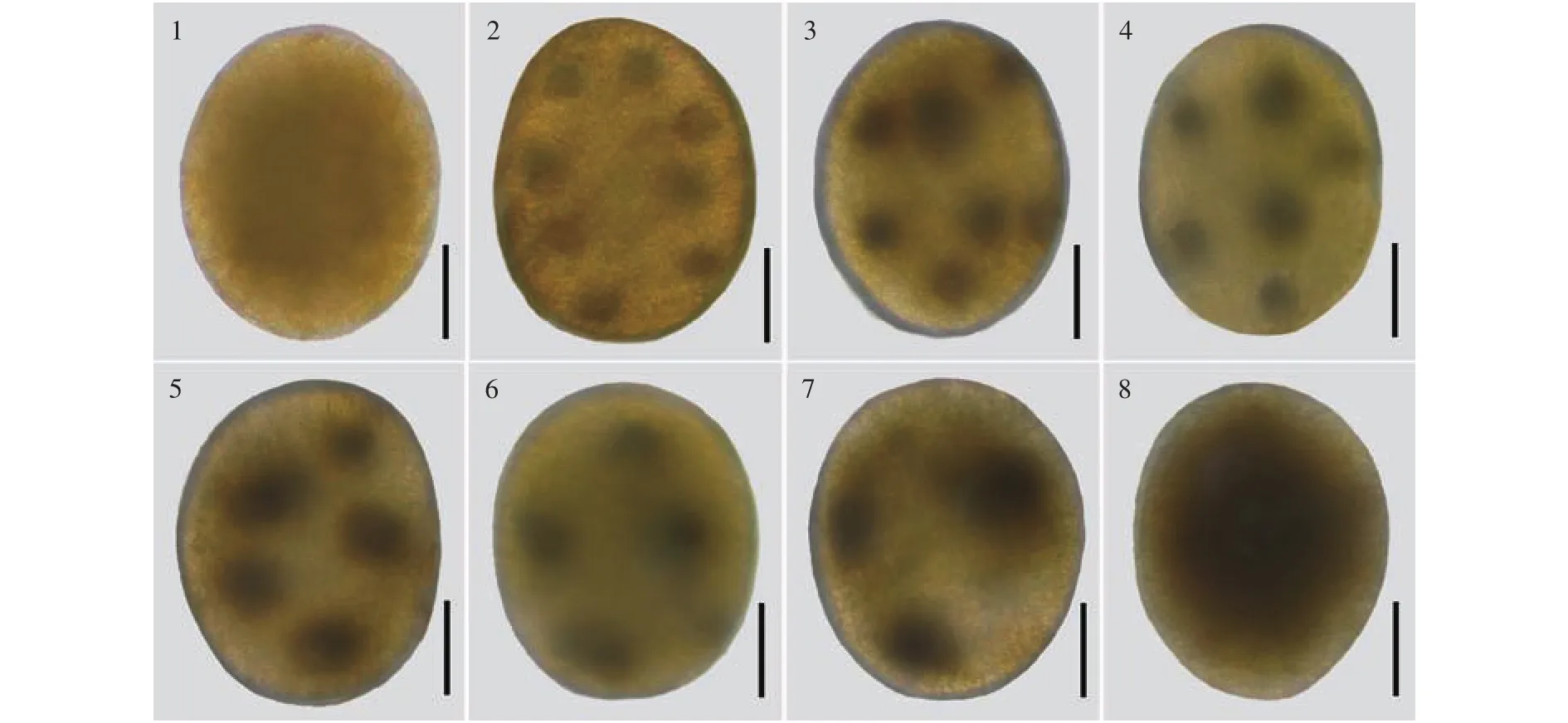
圖版 Ⅱ 銅藻卵子和受精卵內核物質的變化Plate Ⅱ The occurrence of nuclei fusion in the zygote of Sargassum horneri

圖版 Ⅲ 銅藻受精卵的發生過程Plate Ⅲ The embryology of zygote in Sargassaum horneri

圖版 Ⅳ 銅藻幼孢子體的生長發育Plate Ⅳ The development and growth of Sargassum horneri germlings

圖版 Ⅴ 不同附著基條件下銅藻幼孢子體附著發育情況Plate Ⅴ The effect of settlement substratum of Sargassum horneri germlings
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
