嗜酸乳桿菌-地衣芽孢桿菌融合子培養特性研究
2018-04-12 08:41:11要榮宇王靜慧楊喬木吳瑞妮霍乃蕊
食品研究與開發 2018年7期
要榮宇,王靜慧,楊喬木,吳瑞妮,霍乃蕊
(1.四川農業大學動物科技學院,四川成都611130;2.山西出入境檢驗檢疫局,山西太原030024;3.太原市食品藥品監督管理局,山西太原030031;4.山西農業大學動物科技學院,山西太谷030801)
嗜酸乳桿菌屬乳桿菌屬,是乳酸菌中非常重要的益生菌之一,被稱為第三代乳酸發酵劑菌種[1]。嗜酸乳桿菌具有調節腸道菌群、提高免疫力、降低血脂、降低產毒微生物的活性、防止乳糖不耐受等功能。地衣芽孢桿菌屬厚壁菌門,是一種常見的革蘭氏陽性細菌,廣泛存在于土壤、空氣和腐敗有機物中。地衣芽孢桿菌已經應用于醫藥行業,其商品名為整腸生,主要運用于腸道疾病的治療,對腸炎、菌痢等具有明顯療效[2]。
原生質體融合即細胞融合,是指將兩親本菌株的細胞壁通過酶解脫壁,形成球狀的原生質體后,在高滲條件下使其相互接觸,通過膜融合、細胞質融合、核融合,最終基因組之間發生接觸、交換,進而發生基因重組,最后在適宜的條件下再生出細胞壁來,獲得陽性重組子的過程。原生質體融合具有重組頻率高、遺傳穩定、可實現遠緣雜交、可集合雙親優良性狀的優點,所以在微生物育種領域得到廣泛應用[3-5]。王成濤等[6]利用原生質體融合技術將芽孢桿菌T12-1與嗜熱鏈球菌及嗜酸乳桿菌C3進行融合,得到2株高效降低膽固醇的乳酸菌融合子,可顯著降低蛋黃乳等食品中的膽固醇含量,同時兼具乳酸菌發酵食品的風味特征。呂兵等[7]將專性厭氧的短雙歧桿菌和典型的啤酒上面酵母進行原生質體融合,成功篩選出遺傳較穩定的BSF1和BSF2,改善了雙歧桿菌的耐氧特性,為厭氧性益生菌的改良提供了思路。王玉華[8]將2株不同來源的嗜酸乳桿菌La-w1和La-w2融合后獲得菌株La-F1,其耐酸性和耐膽鹽能力明顯高于原菌株。黎永學等[9]將雙歧桿菌和釀酒酵母進行原生質體融合可得到遺傳穩定的融合子,實現了厭氧菌和酵母的跨界融合,為雙歧桿菌生物學功能的開發提供了新途徑。
實驗室在前期成功制得嗜酸乳桿菌-地衣芽孢桿菌融合子,在此基礎上,本試驗進一步優化融合子的培養條件,考察培養基種類、培養溫度、pH值、氧氣濃度對融合子生成量的影響進行分析,確定融合子的適宜培養條件,為融合子的制劑化研究及工業化應用奠定基礎。
1 材料和方法
1.1 材料
1.1.1 菌株
嗜酸乳桿菌(Lactobacillus acidophilus),地衣芽孢桿菌(Bacillus licheniformis)購于中科院微生物研究所。嗜酸乳桿菌-地衣芽孢桿菌融合子由山西農業大學動科院動物衛檢實驗室制備。
1.1.2 試劑與設備
牛肉膏、蛋白胨、胰蛋白胨、葡萄糖、酵母浸膏、吐溫80、瓊脂粉:北京奧博星生物技術有限公司。
硫酸鎂、磷酸氫二鉀、氯化鈉、檸檬酸三銨、硫酸錳、醋酸鈉、溴甲酚紫(均為分析純):天津市化學試劑六廠。
722E型可見分光光度計:上海光譜儀器有限公司;PHS-25C型酸度計:上海大普儀器有限公司;HZQ-F160型全溫振蕩培養箱:哈爾濱市東連電子技術開發有限公司;XW-80A型漩渦混合器:北京科偉永興儀器有限公司;DF205型離心機:北京科偉永興儀器有限公司;LRHS-150型恒溫恒濕培養箱:上海躍進醫療器械廠。
1.1.3 培養基的配制
MRS培養基:蛋白胨10 g,牛肉膏10 g,酵母浸膏5 g,葡萄糖 20 g,Tween-80 1 mL,醋酸鈉 5 g,K2HPO42 g,檸檬酸銨2 g,MgSO4·7H2O 0.58g,蒸餾水1 000 mL,MnSO4·4H2O 0.2 g (固體培養基加入 15 g~20 g瓊脂)pH 6.2~6.6,將mgSO4·7H2O,MnSO4·4H2O,葡萄糖,Tween-80以外的成分溶解,冷至50℃。然后加入MgSO4·7H2O、MnSO4·4H2O,最后加入葡萄糖和 Tween-80。過濾分裝,121℃高壓滅菌15 min。在MRS液體培養基中添加1.5%瓊脂制備固體培養基。
CM培養基(肉湯培養基):蛋白胨10 g,葡萄糖5 g,酵母膏 3 g,牛肉膏 3 g,MgSO4·7H2O 2 g,蒸餾水1 000 mL,pH 7.2。固體培養基瓊脂粉用量為2%(15 g~20 g)。按照所需劑量,將配制好的培養基分裝在三角瓶中,121℃高壓滅菌30 min,備用。
BPM培養基(牛肉膏蛋白胨培養基):牛肉膏3 g,蛋白胨 10 g,NaCl 5 g,蒸餾水 1 000 mL,pH 7.0~7.2。固體培養基瓊脂粉用量為2%(15 g~20 g)。按照所需劑量,將配制好的培養基分裝在三角瓶中,121℃高壓滅菌,備用。
LB培養基:胰蛋白胨10 g,酵母浸膏5 g,NaCl 5 g,蒸餾水1 000 mL,pH 7.0。固體培養基瓊脂粉用量為2%(15 g~20 g)。按照所需劑量,將配制好的培養基分裝在三角瓶中,121℃高壓滅菌,備用。
1.2 試驗方法
1.2.1 融合子菌落形態觀察
將融合子接種到不同液體培養基中,37℃恒溫培養24 h,革蘭氏染色后進行鏡檢,觀察菌體在液體培養基中的個體形態。
將融合子劃線接種到固體培養基中,37℃恒溫培養36 h,長出單菌落,挑取菌體,革蘭氏染色后進行鏡檢,觀察菌體在固體培養基上的個體形態。
1.2.2 融合子培養條件的確定
1.2.2.1 培養基對融合子生長量的影響
按照配方分別配制MRS液體培養基、CM液體培養基、BPM液體培養基、LB液體培養基,高壓蒸汽滅菌。將活化好的融合子在無菌條件下按5%(體積分數)接種量接種于不同液體培養基中,37℃恒溫培養,每3 h取樣一次,測定OD600值。
1.2.2.2 起始pH值對融合子生長量的影響
分別配制pH值為4、5、6、7、8的4種液體培養基。將活化好的融合子以5%(體積分數)接種量在無菌條件下分別接種,37℃恒溫培養12 h,測定其OD600值。
1.2.2.3 培養溫度對融合子生長量的影響
分別配制4種培養基,將活化好的融合子以5%(體積分數)接種量在無菌條件下分別接種,并于20、25、30、35 ℃條件下培養 12 h,測定其 OD600值。
1.2.2.4 培養方式對融合子生長量的影響
1)通氣狀態搖床培養
將活化好的融合子以5%(體積分數)接種量在無菌條件下接種于不同液體培養基中,瓶口加蓋棉塞,37℃恒溫搖床培養,140 r/min,第12小時、第24小時取樣測定OD值。
2)隔氧狀態靜置培養
在無菌條件下,將每種培養基各吸取9.5 mL置于10 mL滅菌的EP管中,然后將活化好的融合子各取0.5 mL接種于4種培養基的EP管中,搖勻后石蠟密封,37℃靜置培養,每12 h取樣一次測定OD值。
1.2.3 融合子產酸性能分析
1.2.3.1 定性產酸實驗
將活化好的融合子以5%(體積分數)接種量在無菌條件下接種于不同液體培養基中,并分別滴入2滴溴甲酚紫溶液,37℃恒溫培養,觀察試管顏色變化。如果試管中的培養液的顏色由紫色變為黃色則說明菌種產酸,否則不產酸。
1.2.3.2 總酸測定
將活化好的融合子以5%(體積分數)接種量在無菌條件下接種于不同液體培養基中,37℃恒溫培養。每隔12小時取不同培養基的培養液10 mL,分別加入蒸餾水20 mL,滴加2滴~3滴0.5%酚酞指示劑,用0.1 mol/L氫氧化鈉標準溶液(現配現用)進行滴定,滴定至微紅為止,根據公式計算出乳酸生成量。
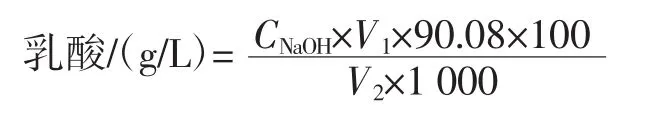
式中:C表示氫氧化鈉的濃度,mol/L;V1表示消耗氫氧化鈉的體積,mL;V2表示加入樣品的體積,mL;90.08表示乳酸的摩爾質量。
2 試驗結果
2.1 融合子的個體形態
2.1.1 融合子在不同液體培養基中的個體形態
圖1(A、B、C、D)分別為融合子在CM液體培養基、LB液體培養基、MRS液體培養基、BPM液體培養基中的個體形態。

圖1 融合子在不同液體培養基中的個體形態(L:10×100)Fig.1 Individual morphology of fusant in different liquid culture medium
經顯微鏡觀察可知,融合子為革蘭氏陽性菌。在CM液體培養基中為桿狀,呈長鏈狀排列;而在LB液體培養基中為短桿菌,單個排列或成短鏈狀排列;融合子在MRS液體培養基中菌數較少,可觀察到的融合子菌體呈長鏈狀排列;在BPM液體培養基中則為短桿菌,單個排列或成短鏈狀排列。
2.1.2 融合子在不同固體培養基中的個體形態

圖2 融合子在不同固體培養基中的個體形態(L:10×100)Fig.2 Individual morphology of fusant in different solid culture medium
圖2(E、F、G)分別為融合子在BPM固體培養基、LB固體培養基、CM固體培養基中的個體形態。融合子在BPM固體培養基中為短桿菌,多為單個排列,產芽孢;在LB固體培養基為短桿菌,多為單個排列或短鏈狀排列,產芽孢;在CM固體培養基中為單個排列或短鏈狀排列并產芽孢的桿菌。
融合子在BPM固體培養基中為短桿菌,多為單個排列,產芽孢;在LB固體培養基為短桿菌,多為單個排列或短鏈狀排列,產芽孢;在CM固體培養基中為單個排列或短鏈狀排列并產芽孢的桿菌。
2.2 不同培養基對融合子生長的影響
圖3為融合子在不同培養基中的生長曲線圖。
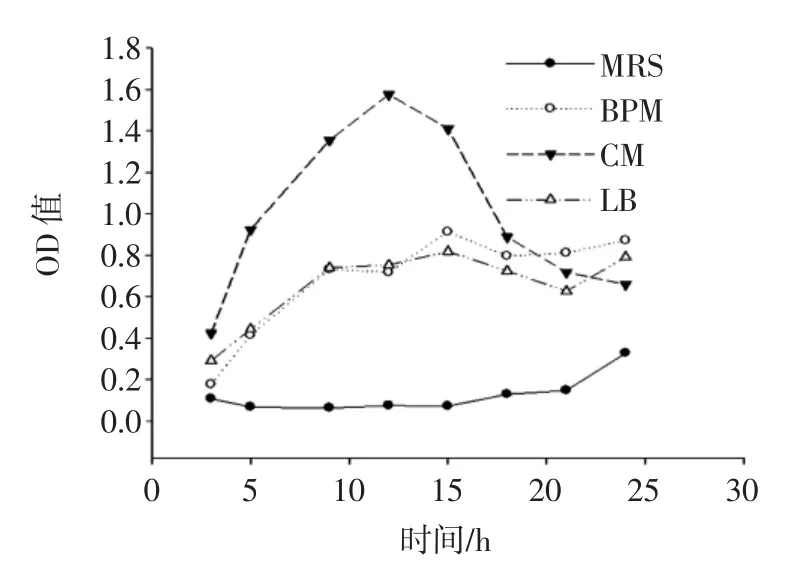
圖3 融合子在不同培養基中的生長曲線Fig.3 Growth curve of fusant in different medium
由圖3可知,融合子在MRS培養基中的OD值顯著低于其他培養基(P<0.05),且延滯期較長;融合子在BPM培養基和LB培養基中,3 h~9 h為對數生長期,之后進入穩定期;在CM培養基中,3 h~12 h為對數生長期,12 h后進入衰亡期,12 h時其OD值(1.576±0.011)顯著高于其他培養基(P<0.05),說明CM培養基對融合子的生長更加有利。
2.3 起始pH值對融合子生長量的影響
圖4為培養基的不同pH值對融合子生長量的影響。
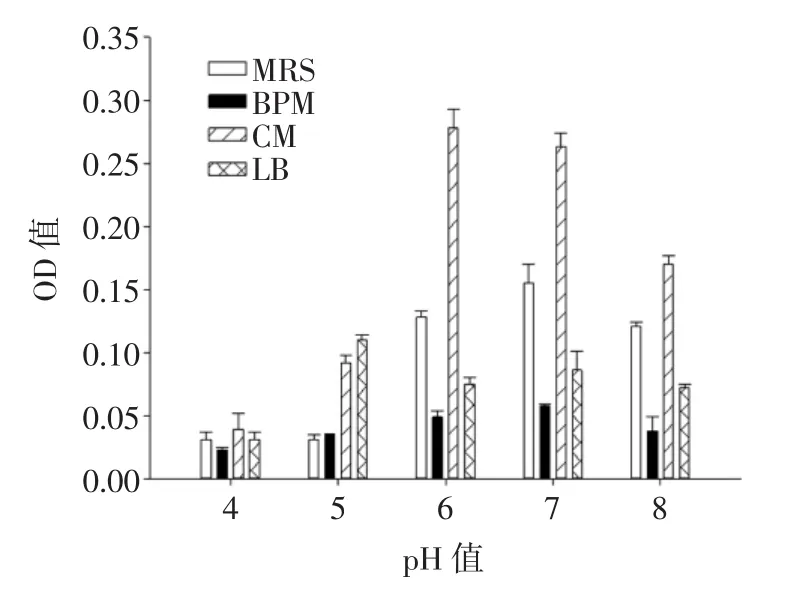
圖4 培養基不同起始pH值對融合子生長量的影響Fig.4 Effect of different initial pH values on the growth of the fusant
如圖4所示,融合子在MRS培養基、BPM培養基和LB培養基中生長,雖然pH值不同,OD值也不同,但是差異并不顯著(P>0.05)。而在CM培養基中,pH值的差異導致OD值變化顯著(P<0.05),隨著pH值的升高,OD值呈現先升高后降低的趨勢,在pH值為6時,融合子OD值最高,達到0.292。由圖4可知,在MRS培養基、BPM培養基、CM培養基和LB培養基中,融合子的最適生長pH值分別為7、7、6、5。
2.4 培養溫度對融合子生長量的影響
圖5為培養溫度對融合子生長的影響。
由圖5可知,融合子在MRS培養基和BPM培養基中,培養溫度對OD值的影響并不顯著(P>0.05);在LB和CM培養基中,隨著溫度的增高,OD值均呈現先升高后降低的趨勢,特別是溫度對生長在CM培養基中的融合子影響顯著,溫度為37℃時,OD值顯著增加并達到最高。由圖可確定,在MRS培養基、BPM培養基、CM培養基和LB培養基中,融合子的最適生長溫度分別為 37、35、37、35 ℃。
2.5 培養方式對融合子生長量的影響
將融合子接種于不同培養基中,分別采用有氧培養(通氧搖床)和厭氧培養(隔氧靜置)兩種方式進行培養,測定培養液OD值,結果如表1所示。
由表1可知,融合子在MRS培養基中隔氧靜置培養12 h和24 h,所測得的OD值均高于通氧搖床培養測定的OD值。融合子在BPM培養基、CM培養基及LB培養基中,通氧搖床所測定的OD值均高于隔氧靜置培養。說明在MRS培養基中,隔氧靜置培養更有利于融合子生長;而在BPM培養基、CM培養基及LB培養基中,通氧培養方式更為適合。
2.6 融合子產酸性能分析
2.6.1 定性產酸實驗結果
通過融合子溴甲酚紫顯色反應結果發現:MRS培養基和CM培養基均呈黃色,說明融合子在這兩種培養基中能夠產酸;BPM培養基和LB培養基呈紫色,說明融合子在這兩種培養基中產酸不佳。
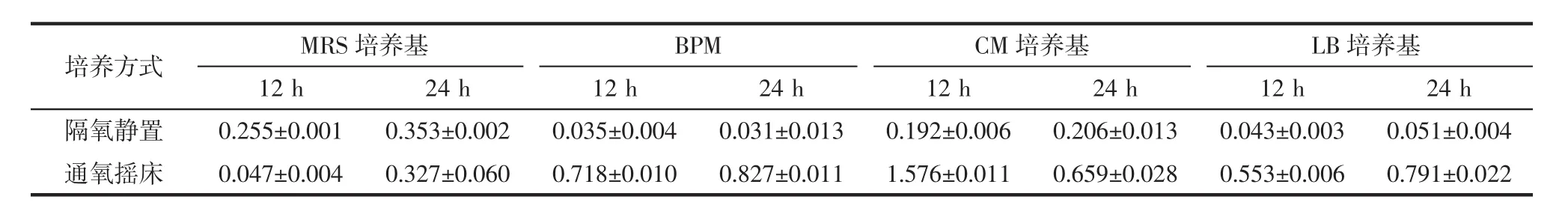
表1 培養方式對融合子生長量的影響Table 1 Effects of culture methods on the growth of fusant
2.6.2 融合子總酸測定結果
融合子在不同培養基中的乳酸生成量如圖6所示。
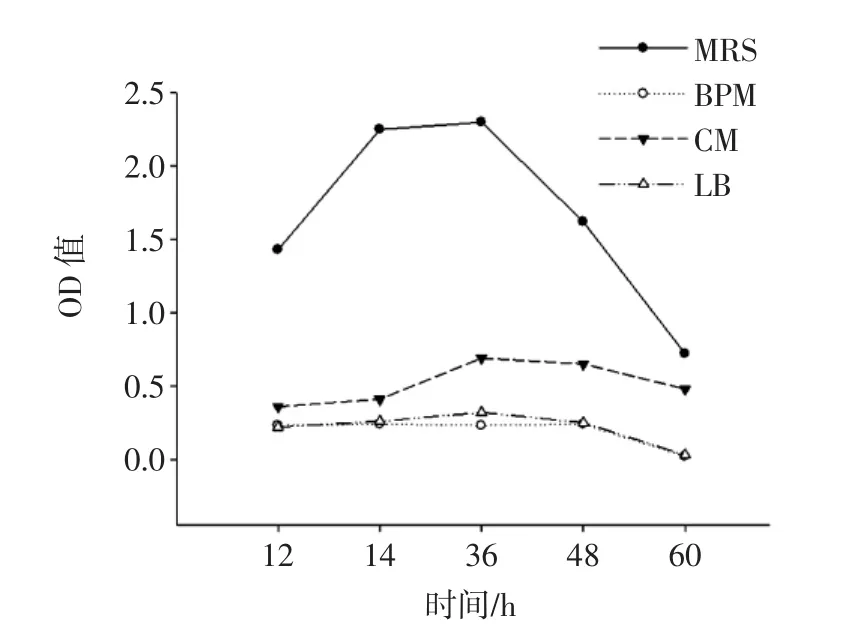
圖6 不同培養基中融合子乳酸生成量測定Fig.6 Determination of lactic acid production in different medium
由圖6可知,融合子在4種培養基中的乳酸生成量均呈現先升高后降低的趨勢。在培養前期,培養液中的總酸積累較少,酸度環境較為適宜融合子的生長,培養基中乳酸量不斷升高;培養一段時間后,培養液中總酸積累量較高,酸度上升,此時酸度環境可能會對融合子的生長產生抑制,甚至發生自溶,從而導致產酸量下降。在不同培養時段,融合子在MRS培養基中乳酸生成量顯著高于其他培養基,培養36 h,乳酸生成量最高,達到2.30 g/L。
3 討論與結論
對于微生物來說,即使是同一菌株,在不同培養條件下進行培養,其生長情況和代謝方式也不盡相同[10]。目前,關于嗜酸乳桿菌和地衣芽孢桿菌培養特性的相關報道已經較為詳盡,而嗜酸乳桿菌-地衣芽孢桿菌融合子的培養特性尚未見報道。嗜酸乳桿菌為乳酸菌屬,一般采用MRS培養基37℃培養。地衣芽孢桿菌的最佳生長溫度為35℃,最適pH值為7.0。本試驗發現CM培養基(肉湯培養基)成為最適合嗜酸乳桿菌-地衣芽孢桿菌融合子生長的培養基,其次為BPM培養基和LB培養基。CM培養基是最經典的細菌培養基之一,可以用于各種細菌的培養。BPM瓊脂培養基和LB培養基可為微生物生長提供氮源、碳源、磷酸鹽、維生素和無機鹽。MRS培養基是用于鑒別、分離乳酸菌的專用培養基[11]。本試驗通過鏡檢觀察發現融合子在液體和固體培養基上的菌落形態與親本的菌落形態明顯不同,這是基因重組的結果[12-13]。基因重組不僅導致融合子的菌落形態發生變化,而且最適培養基的種類也發生了改變。嗜酸乳桿菌在MRS培養基中生長良好,但是與地衣芽孢桿菌融合后,CM成為了最適培養基,培養方式也以通氧搖床培養為佳。但是就產酸性能分析,融合子在MRS培養基中的產酸量最大,說明MRS培養基更有利于融合子的繁殖代謝,產生包括乳酸在內的次級產物。
嗜酸乳桿菌和地衣芽孢桿菌的融合子兼具兩親本的優良特性。乳酸菌和地衣芽孢桿菌都是對人體有益的益生菌,融合子既具備了地衣芽孢桿菌較強的產蛋白酶、產淀粉酶能力以及形成芽孢的能力,也具有了乳酸菌耐受胃部的低酸環境以及腸道前段高膽鹽環境的能力,腸道定殖能力大大增強[14],同時還克服了其不易培養和制劑化的缺陷,該融合子在厭氧和有氧環境中都能很好生長,可用于發酵乳、發酵肉制品、奶酪等食品的生產,也可用于L-乳酸、蛋白酶、淀粉酶等制劑的發酵生產,并能作為新一代乳酸菌或地衣芽孢桿菌整腸生制劑生產用的菌株。
通過本試驗研究可知在常規培養條件下,CM培養基最有利于融合子的生長。CM、MRS和LB培養基的適宜初始pH值分別為6、7和5,BPM培養基初始pH值對融合子的生長繁殖影響不顯著。在CM培養基中,37℃更適于融合子生長,其他培養基的最適培養溫度均為35℃。除MRS培養基適合隔氧靜置培養外,其他3種培養基則配以通氧搖床培養方式。MRS培養基有利于融合子發酵產酸,第36小時時,乳酸生成量最高,達到2.30 g/L。
參考文獻:
[1]LI S,MA C J,GONG G G,et al.The impact of onion juice on milk fermentation by Lactobacillus acidophilus[J].LWT-Food Science and Technology,2016,65(1):543-548
[2]馬鑫,郭宏,張寶國,等.地衣芽孢桿菌作為飼料添加劑的研究進展[J].中國飼料添加劑,2011(2):10-13
[3]趙春苗,徐春厚.原生質體融合技術及在微生物育種中的應用[J].中國微生態學雜志,2012,24(4):379-382
[4]王登宇,臧威,孫劍秋,等.細菌原生質體融合育種技術及其應用進展[J].中國釀造,2008,27(7):1-5
[5]邱靜,羅水忠,姜紹通,等.高產L-乳酸米根霉的原生質體制備與再生條件研究[J].食品科學,2011,32(9):174-178
[6]王成濤,牛天貴,岳曉禹,等.應用原生質體融合技術構建高效降解膽固醇的乳酸菌[J].食品與發酵工業,2002,28(3):1-5
[7]呂兵,項建琳.應用原生質體融合技術改善雙歧桿菌的耐氧性[J].食品科學,2005,26(4):83-86
[8]王玉華,張桂榮,劉景圣.原生質體融合提高嗜酸乳桿菌耐酸及耐膽鹽能力[J].食品科學,2006,27(3):96-99
[9]黎永學,張德純,李代昆.雙歧桿菌和釀酒酵母原生質體融合子篩選方法的探討[J].食品科學,2006,27(2):84-86
[10]COURTIN P,RUL F.Interactions between microorganisms in a simple ecosystem:yogurt bacteria as a study model[J].Le Lait,2003,84(1/2):125-134
[11]杜連祥.工業微生物學實驗技術[M].天津:天津科學技術出版社,1992:70-90
[12]霍乃蕊,韓克光.細胞融合技術的發展及應用[J].激光生物學報,2006,15(2):209-213
[13]趙春苗,徐春厚.原生質體融合技術及在微生物育種中的應用[J].中國微生態學雜志,2012,24(4):379-382
[14]曾獻春,孟冬麗.乳酸菌原生質體制備與再生研究[J].食品科學,2006,27(10):269-272
猜你喜歡
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
今日農業(2021年19期)2022-01-12 06:16:36
中老年保健(2021年11期)2021-08-22 03:15:44
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化(高中版.高考數學)(2021年1期)2021-03-19 08:28:38
無線電工程(2020年11期)2020-10-29 01:25:46
現代出版(2020年3期)2020-06-20 07:10:34
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
