山西省旱柳古樹(shù)資源遺傳多樣性的SSR分析
2018-04-26 05:46:38王振龍高志文樊明瑞韓志校
山西林業(yè)科技 2018年1期
張 亮,王振龍,高志文,馬 晶,樊明瑞,韓志校
(1.山西省太原市林業(yè)科學(xué)研究所,山西 太原 030009;2.河北農(nóng)業(yè)大學(xué),河北 保定 071000 )
旱柳(SalixmatsudanaKoidz.),屬于楊柳科柳屬落葉大喬木,其分布廣泛,在全國(guó)各地都有栽培,主要品種與變種有饅頭柳、絳柳、龍須柳等。它是防護(hù)林、水土保持林、園林綠化林及用材林的重要組成樹(shù)種,具有較高的生態(tài)價(jià)值和經(jīng)濟(jì)價(jià)值。古樹(shù)是有生命的活化石,是大自然給人類的寶貴財(cái)富,具有重要的科研、歷史、文化價(jià)值,保護(hù)好它們有著十分重要的意義。
SSR已成為在生物上應(yīng)用最廣、最為重要的分子標(biāo)記。目前,SSR標(biāo)記已經(jīng)在眾多植物種類中得到證實(shí)和應(yīng)用,例如,葡萄、松樹(shù)等物種。而柳樹(shù)分子標(biāo)記的研究始于20世紀(jì)90年代,如,王輝等利用SSR標(biāo)記的方法對(duì)楊柳樹(shù)遺傳多樣性進(jìn)行了分析,劉恩英建立和優(yōu)化了柳樹(shù)SSR反應(yīng)體系,王源秀利用杞柳和簸箕柳候選雜交親本對(duì)其SSR指紋進(jìn)行了分析等。而對(duì)山西省不同地區(qū)旱柳古樹(shù)資源遺傳多樣性的SSR研究還未見(jiàn)報(bào)道。筆者嘗試用SSR分子標(biāo)記的方法對(duì)采自山西省7個(gè)地區(qū)的49份旱柳古樹(shù)材料進(jìn)行遺傳多樣性檢測(cè),旨在從DNA分子水平闡明其地區(qū)間的遺傳差異,為旱柳古樹(shù)資源的有效保護(hù),柳樹(shù)新品種的選育及遺傳改良抗性研究提供理論支持,并從分子水平上豐富對(duì)古樹(shù)的研究。
1 材料和方法
1.1 材料
2009年至2011年,太原市林業(yè)科學(xué)研究所柳樹(shù)課題組收集了山西省境內(nèi)的古旱柳品種(見(jiàn)圖1)。通過(guò)嫁接復(fù)壯、扦插等繁殖手段,至2012年已培育出49個(gè)合格的古旱柳品種2年生苗木各200株。于當(dāng)年秋季苗木生長(zhǎng)停止后,在樹(shù)冠1/3處,采集枝條、葉片等材料,封裝進(jìn)冷藏保溫箱并做好標(biāo)記,在4 h內(nèi)送至河北農(nóng)業(yè)大學(xué)進(jìn)行試驗(yàn)。
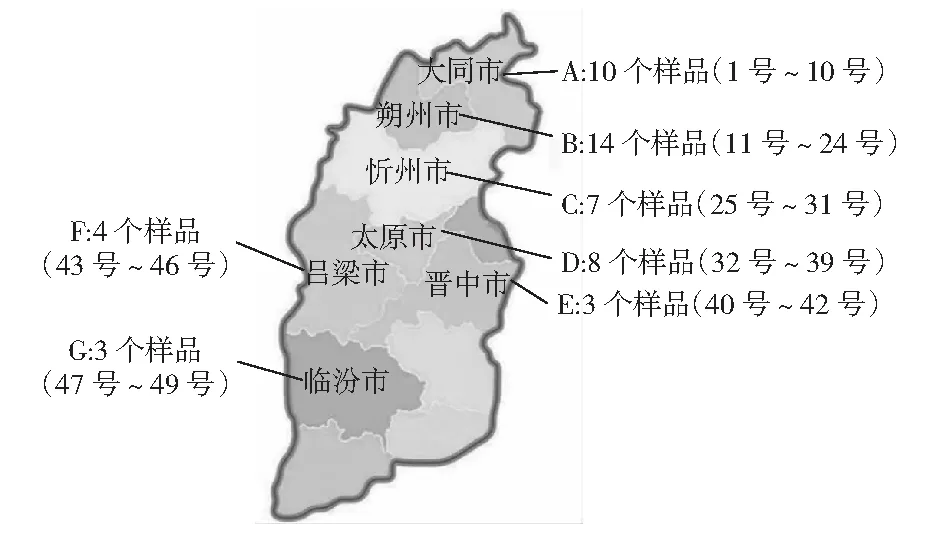
圖1 供試材料所在地理位置及取樣情況
1.2 方法
基因組DNA提取采用改良的CTAB法。用1.0%瓊脂糖凝膠電泳檢測(cè)DNA質(zhì)量,經(jīng)紫外分光光度計(jì)測(cè)定純度后,再用ddH2O稀釋至20 ng/μL左右。
試劑:所用引物均來(lái)自文獻(xiàn)[12-13],并由上海生工生物工程技術(shù)服務(wù)有限公司和北京天一輝遠(yuǎn)生物科技有限公司合成。2×Taq MasterMix購(gòu)自北京康為世紀(jì)生物科技有限公司。
PCR反應(yīng)體系:柳樹(shù)DNA基因組SSR-PCR 10 μL 反應(yīng)中最優(yōu)條件為2×Taq MasterMix 5μL,引物濃度為1.0 μmol/L,H2O為3μL,DNA為1μL.
PCR擴(kuò)增反應(yīng):PCR擴(kuò)增94 ℃預(yù)變性5 min后,95 ℃變性50 s,52 ℃復(fù)性50 s,72 ℃延伸1 min,35個(gè)循環(huán)。最后一個(gè)循環(huán)結(jié)束后,再72 ℃延伸7 min,得擴(kuò)增產(chǎn)物。
反應(yīng)產(chǎn)物采用8%非變性聚丙烯酰胺凝膠電泳,電泳40 min左右,取出進(jìn)行AgNO3染色。再用顯色液(1.5%NaOH,0.4%甲醛)顯色,直至條帶清晰為止,照相并記錄。
1.3 數(shù)據(jù)統(tǒng)計(jì)分析
選擇擴(kuò)增條帶清晰且有多態(tài)性的SSR標(biāo)記進(jìn)行數(shù)據(jù)統(tǒng)計(jì),假定膠上遷移率相同的條帶均來(lái)自同一位點(diǎn)上的同一等位基因。對(duì)每個(gè)樣品的擴(kuò)增電泳譜帶進(jìn)行統(tǒng)計(jì),有帶記為1,無(wú)帶或模糊的記為0,形成0/1數(shù)據(jù)矩陣輸入Excel中。SSR數(shù)據(jù)用POPGENE32和DPS軟件進(jìn)行遺傳多樣性數(shù)據(jù)計(jì)算。
2 結(jié)果與分析
2.1 古旱柳引物篩選與多態(tài)性分析
從25對(duì)引物中選取10對(duì)擴(kuò)增條帶清晰、重復(fù)性好、特異性高的引物。這10對(duì)引物在7個(gè)地區(qū)的49個(gè)旱柳古樹(shù)品種中共擴(kuò)增出條帶33條,其中多態(tài)性條帶26條,占78.79%(如表1).每對(duì)引物擴(kuò)增位點(diǎn)數(shù)為2~5,平均每對(duì)引物擴(kuò)增3.3個(gè)位點(diǎn)。
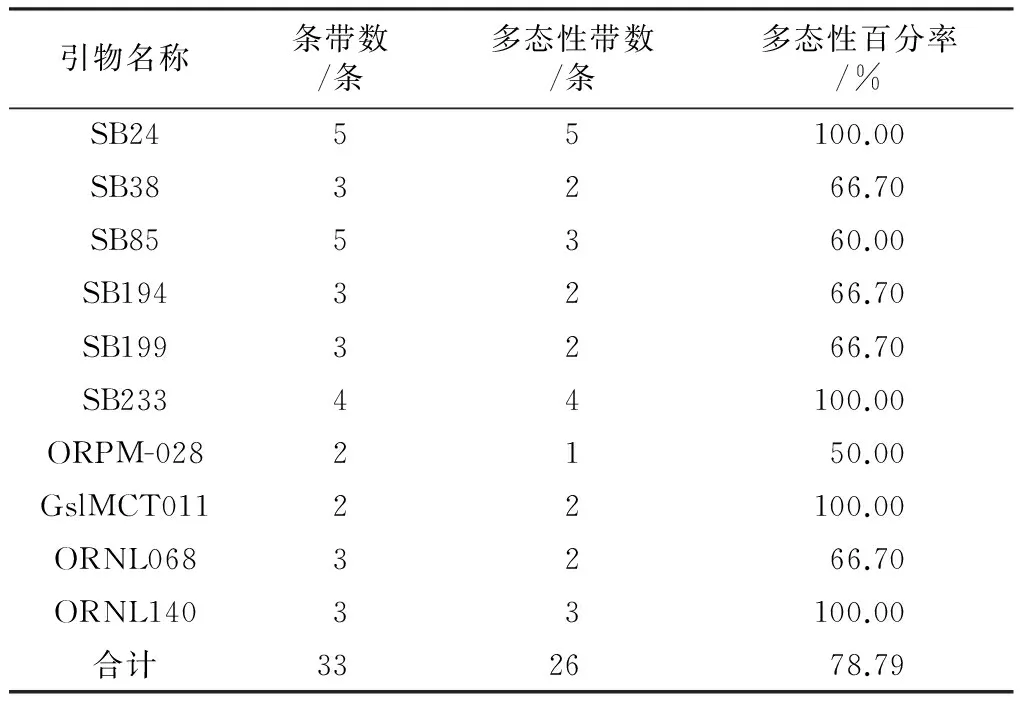
表1 引物多態(tài)性分析
從擴(kuò)增結(jié)果來(lái)看,SSR引物得到的擴(kuò)增產(chǎn)物片斷大小主要分布在80 bp~320 bp之間。其中引物SB24,SB233,GslMCT011,ORNL140多態(tài)性最高,達(dá)到100%;ORPM-028多態(tài)性最低,為50%;其余引物介于兩者之間。
圖2為引物SB24和引物ORPM-028擴(kuò)增出的部分樣品圖譜。
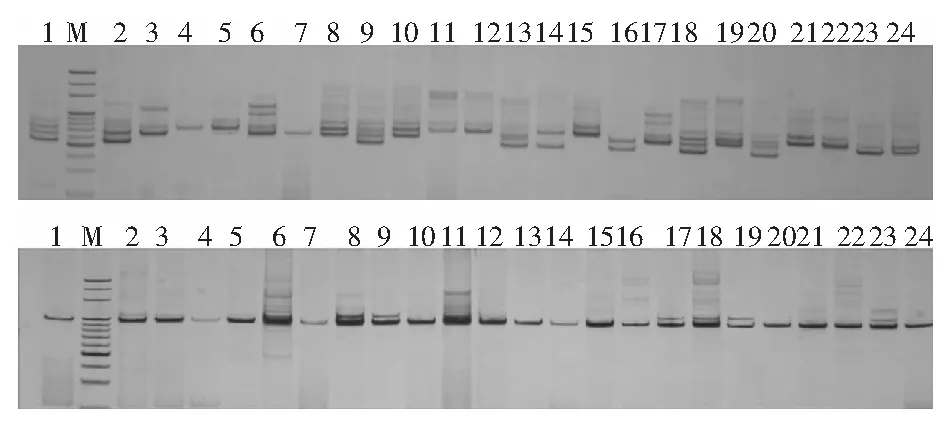
圖2 SB24引物和ORPM-028引物對(duì)供試的部分旱柳古樹(shù)的SSR擴(kuò)增譜帶
從圖2可以看出,不同引物在相同材料中產(chǎn)生的譜帶信息不同,同一引物在不同的材料中產(chǎn)生的譜帶信息也不相同。這說(shuō)明供試的旱柳古樹(shù)在DNA+水平上具有一定的差異。
2.2 古旱柳種群內(nèi)遺傳多樣性參數(shù)
地區(qū)間旱柳古樹(shù)的多態(tài)位點(diǎn)百分率、Nei′s基因多樣性和有效等位基因范圍分別為39.39%~69.70%,0.121 6~0.208 6和1.189 3~1.341 2,見(jiàn)表2.
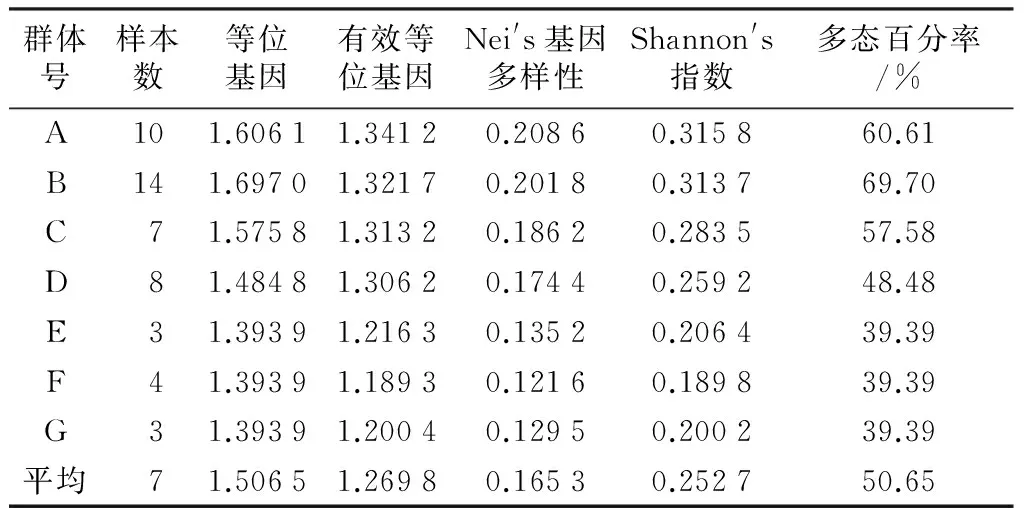
表2 7個(gè)地區(qū)間旱柳古樹(shù)資源的遺傳多樣性參數(shù)值
從表2中可以看出,7個(gè)地區(qū)間多態(tài)性比率差異較大,在39.39%~69.70%之間。其中朔州(地區(qū)B)地區(qū)的多態(tài)性比率最高,達(dá)到了69.70%;晉中(地區(qū)E)、臨汾(地區(qū)G)、呂梁(地區(qū)F)最低,為39.39%.7個(gè)地區(qū)的多態(tài)性高低為:B>A>C>D>E,G,F(xiàn).各群體間Nei′s基因多樣性和有效等位基因排列與多態(tài)位點(diǎn)百分率一致。
2.3 古旱柳群體間遺傳一致度及遺傳距離
古旱柳群體的遺傳一致度(I)與遺傳距離(D)見(jiàn)表3.
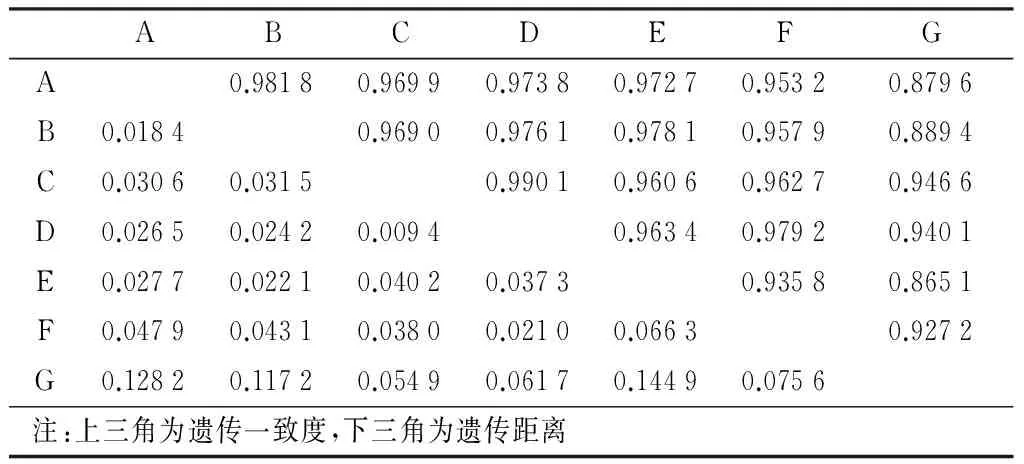
表3 旱柳古樹(shù)群體的遺傳一致度與遺傳距離
由POPGENE32軟件計(jì)算的7個(gè)地區(qū)間旱柳古樹(shù)的遺傳距離和遺傳一致度表明,古旱柳群體間的遺傳一致度很高(0.865 1~0.990 7),尤其是地區(qū)C與地區(qū)D之間遺傳相似度最大(0.990 7),接近于1,表明兩個(gè)地區(qū)幾乎沒(méi)有分化。地區(qū)E與地區(qū)G之間的遺傳一致度最低(0.865 1)。遺傳距離在地區(qū)E和地區(qū)G之間最大(D=0.144 9),地區(qū)C和地區(qū)D之間最小(D=0.009 4)。
不同地區(qū)旱柳古樹(shù)的遺傳聚類圖見(jiàn)圖3.
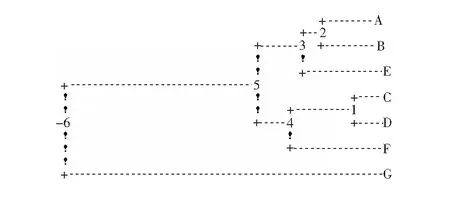
圖3 不同地區(qū)旱柳古樹(shù)的遺傳聚類圖
由圖3可以看出,7個(gè)地區(qū)的旱柳古樹(shù)資源通過(guò)聚類分成了兩大類,第Ⅰ類為:大同(地區(qū)A)、朔州(地區(qū)B)、忻州(地區(qū)C)、太原(地區(qū)D)、晉中(地區(qū)E)、呂梁(地區(qū)F);第Ⅱ類為:臨汾(地區(qū)G)。這也說(shuō)明了處在山西省中北部6個(gè)地區(qū)的旱柳古樹(shù)聚在了一起,而處在南部臨汾地區(qū)的單獨(dú)聚在了一起。
2.4 不同地區(qū)古旱柳材料的聚類分析
利用DPS軟件中的UPGMA法對(duì)分布在山西省7個(gè)地區(qū)的49份旱柳古樹(shù)材料進(jìn)行聚類分析,結(jié)果見(jiàn)圖4.
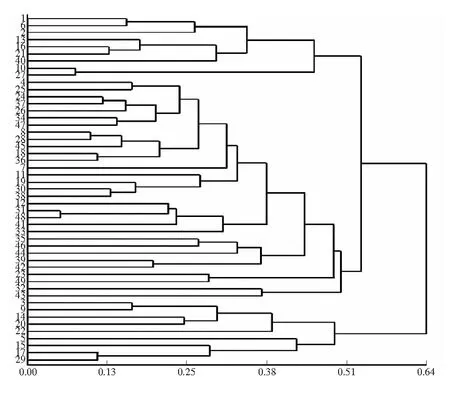
圖4 不同地區(qū)旱柳古樹(shù)植株的遺傳聚類分析
由圖4可以看出,在遺傳距離為0.53處,所有植株可以分為兩類:一類是包括所有地區(qū)的40個(gè)古旱柳品種;一類是包括大同、朔州、忻州3個(gè)地區(qū)的9個(gè)古旱柳品種。說(shuō)明這9個(gè)品種與上述40個(gè)古旱柳品種有一定的差異性,同時(shí),49個(gè)古旱柳品種個(gè)體的聚類結(jié)果與7個(gè)地區(qū)間的聚類有差別。
3 討論與結(jié)論
研究表明,一個(gè)物種遺傳多樣性越高,對(duì)環(huán)境變化的適應(yīng)能力就越強(qiáng),越容易擴(kuò)展其分布范圍和開(kāi)拓新環(huán)境。筆者用10對(duì)引物對(duì)采自山西省7個(gè)地區(qū)的49個(gè)旱柳古樹(shù)植株進(jìn)行了地區(qū)間遺傳多樣性分析,結(jié)果表明,其具有豐富的遺傳多樣性。選用的10對(duì)引物共擴(kuò)增出條帶33條,其中多態(tài)性條帶有26條,占78.79%.由此可見(jiàn),這7個(gè)地區(qū)的旱柳古樹(shù)材料具有比較豐富的SSR多態(tài)性。
Nei′s遺傳多樣性揭示了各地區(qū)間遺傳多樣性水平,利用SSR分子標(biāo)記方法測(cè)得的7個(gè)地區(qū)旱柳古樹(shù)的Nei′s遺傳多樣性范圍為0.121 6~0.208 6,均值為0.165 3,其排列順序依次是:朔州>大同>忻州>太原>晉中、臨汾、呂梁,它與地區(qū)間的多態(tài)性比率一致。說(shuō)明這7個(gè)地區(qū)間旱柳古樹(shù)具有較豐富的遺傳多樣性。
通過(guò)對(duì)這7個(gè)地區(qū)的旱柳古樹(shù)進(jìn)行聚類分析可知,地區(qū)間的遺傳一致度很高,尤其是地區(qū)C與地區(qū)D之間遺傳相似度最大,接近于1,表明兩地區(qū)幾乎沒(méi)有分化;地區(qū)E與地區(qū)G之間的遺傳一致度最低。7個(gè)地區(qū)間聚類結(jié)果得到兩類,并且具有地理相關(guān)性,一類是種植在山西省中北部6個(gè)地區(qū)的旱柳古樹(shù),一類是種植在山西省南部臨汾地區(qū)的旱柳古樹(shù)。
對(duì)來(lái)自這7個(gè)地區(qū)的49個(gè)品種個(gè)體之間的聚類結(jié)果表明,雖然柳樹(shù)主要是通過(guò)無(wú)性繁殖,但是來(lái)自山西省7個(gè)不同地區(qū)的49個(gè)旱柳古樹(shù)品種間卻存在遺傳差異,并非起源于同一無(wú)性系。另外,49個(gè)旱柳古樹(shù)材料聚類結(jié)果呈現(xiàn)混雜性,并不是同一種植地區(qū)的植株聚在一起。說(shuō)明這些古旱柳個(gè)體是在幾十年至幾百年前栽種的,由于時(shí)間不同,應(yīng)用的品種可能不同。雖然是在同一地區(qū)甚至同一地點(diǎn)收集的旱柳古樹(shù),其遺傳差異也可能會(huì)很大。從而也進(jìn)一步說(shuō)明了這些旱柳古樹(shù)具有廣泛的遺傳多樣性。
山西省旱柳古樹(shù)存在廣泛的遺傳多樣性,是寶貴的遺傳資源,應(yīng)加大保護(hù)和研究力度,為柳樹(shù)的遺傳改良提供育種材料。
參考文獻(xiàn):
[1] 韓 騫,王 輝,王進(jìn)茂,等.利用SSR引物通用性分析楊柳科樹(shù)種遺傳多樣性[J].分子植物育種,2009,7(5):904-911.
[2] 劉永華.瀏陽(yáng)市古樹(shù)資源調(diào)查與評(píng)價(jià)[J].湖南林業(yè)科技,2009,36(1):73-75.
[3] 李小龍.北沙柳種質(zhì)資源庫(kù)遺傳多樣性的SSR分子標(biāo)記分析[D].內(nèi)蒙古自治區(qū):內(nèi)蒙古農(nóng)業(yè)大學(xué),2011.
[4] ScottK D,Eggler P,Seaton G,et al.Analysis of SSRs derived from grape ESTs[J].Theor Appl Genet ,2000,100:723-726.
[5] Carol E Ritland,Yousry A Kassaby,Kermit Ritland.Single copy species transferable microsatellite markers developed from loblolly pine ESTs[J].Theor Appl Genet,2004,109:361-369.
[6] 王 輝.楊柳樹(shù)SSR遺傳多樣性分析及4種柳樹(shù)耐鹽性比較研究[D].保定:河北農(nóng)業(yè)大學(xué),2008.
[7] 劉恩英,王源秀,孫晨晨,等.柳樹(shù)SSR反應(yīng)體系的建立與優(yōu)化[J].江蘇林業(yè)科技,2008,35(5):8-11;16.
[8] 王源秀,徐立安,黃敏仁.杞柳和簸箕柳候選雜交親本SSR指紋分析[J].南京林業(yè)大學(xué)學(xué)報(bào):自然科學(xué)版,2008,32(2):1-5.
[9] 陳 靜,王文江.適用AFLP分析的核桃幼葉DNA提取方法[J].河北農(nóng)業(yè)大學(xué)學(xué)報(bào),2004,27(6):45-48.
[10] 王卓偉,余茂德,魯 成.PVP在桑葉總DNA提取中的應(yīng)用[J].西南農(nóng)業(yè)大學(xué)學(xué)報(bào),2001,23(1):61-65.
[11] ALLEN G C,F(xiàn)LORES-VERGARA M A,KRASYNANSKI S,et al.A modified protocol for rapid DNA isolation from plant tissues using cetyltrimethyl ammonium bromide[J].Nature Protocols,2006,1(5):2 320-2 325.
[12] 梁海永,劉興菊,楊敏生.利用RAMP-PCR技術(shù)對(duì)枸杞10個(gè)品種資源的分析[J].中國(guó)農(nóng)學(xué)通報(bào),2011,27(16):61-64.
[13] 盧欣石,何 琪.種群遺傳變異及基因多樣度分析[J].草業(yè)學(xué)報(bào),1999,8(3):76-82.
