綿羊MHC-DQB2 exon3單倍型的構建及其與布魯氏菌病易感性相關性
2018-06-07 02:47:34王元元齊江姣高劍峰
江蘇農業科學 2018年10期
嚴 國, 王元元, 羅 成, 齊江姣, 牟 云, 高劍峰
(石河子大學生命科學學院,新疆石河子 832003)
布魯氏菌病簡稱布病,是由布魯氏桿菌引起的嚴重危害人民健康和農牧業發展的人畜共患傳染病,是《中華人民共和國傳染病防治法》規定的乙類傳染病,羊為該病最主要的傳染源[1]。調查顯示,2012—2014年新疆牛羊布病陽性率呈逐年上升趨勢[2]。因此,這一疾病對新疆這一畜牧業主產區所造成的損失也逐年增長,遏制該疾病的傳播已成為亟待解決的重大問題。
主要組織相容性復合體(major histocompatibility complex,MHC)是脊椎動物的高度多態性基因群[3]。MHC蛋白的主要功能是抗原遞呈,在機體的免疫系統中發揮著至關重要的作用,與許多疾病的抗病性和易感性密切相關,MHC作為相關性研究的候選基因,受各研究領域的密切關注,已經成為現代分子遺傳研究的焦點。綿羊的MHC位于20號染色體,MHCⅡ區與人的相似,區域內的 DQ 和 DR 2個基因家族表現豐富的多態性[4],而綿羊的多態性更集中在 DQ 家族[5-7]。
單核苷酸多態性(single nucleotide polymorphism,SNP),是指由單個堿基的變化引起DNA序列發生改變的多態性,是基因組序列內最為常見的變異形式。SNP作為第三代分子遺傳標記,本身具有顯著特點,如數量多、分布廣泛、且穩定遺傳等[8]。這些特點在疾病相關基因的研究中發揮著尤為重要的作用,尤其是那些位于基因編碼區的SNPs,可能會使基因編碼的蛋白質的產量或質量發生改變,使其與某種疾病的發生直接相關,因而備受各研究領域的關注[9]。由于單個SNP在復雜疾病中所起到的作用微小[10-11],而由SNPs構建的單倍型提供的遺傳信息卻比單個的SNP更多、更準確、更符合多基因疾病的遺傳性,所以單倍型分析越來越成為復雜疾病關聯研究的有效工具,這為相關免疫性狀的關聯研究及抗病分子標記輔助選擇奠定了一定的基礎。因此,本研究通過 PCR-SSCP 技術結合測序的方法檢測哈薩克羊的MHC-DQB2 exon3的多態位點,探討SNPs與布魯氏菌病易感性相關性,此外構建其單倍型,并初步推測與布魯氏菌病易感性相關的單倍型,為今后開展布魯氏菌抗性分子標記輔助選擇研究提供了一定的依據,為進一步定位具有易感性的遺傳位點而最終進行抗病育種奠定基礎。
1 材料與方法
1.1 試驗材料
1.1.1 樣本來源 本試驗所用血液樣本部分(45支)來自于新疆維吾爾自治區畜牧獸醫站,全部為布魯氏菌病陽性;部分(100支)采自新疆烏蘇市3個羊場,以頸靜脈采血法采血 5 mL 于肝素鈉抗凝管,4 ℃保存,以備基因組DNA提取。
1.1.2 主要試劑 布魯氏菌檢測試劑盒購自哈藥集團生物疫苗有限公司;Tris平衡酚購自北京索萊寶科技有限公司;引物由北京六合華大基因科技股份有限公司合成;瓊脂糖為西班牙瓊脂糖(Biowest Agarose);2×TaqPCR Master Mix 和 DL 2000 marker購自北京天根公司;過硫酸銨購自美國Sigma公司;丙烯酰胺、甲叉雙丙烯酰胺、TEMED購自美國Promega公司;親和硅烷、剝離硅烷購自北京鼎國生物技術中心。
1.2 試驗方法
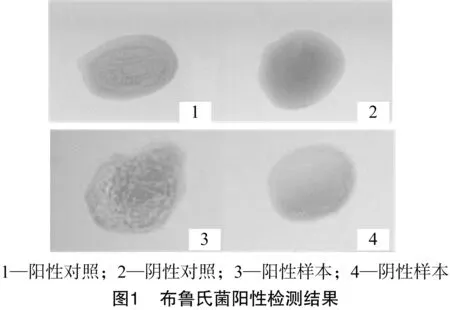
1.2.1 布魯氏菌陽性檢測 采用虎紅平板檢測法對100只哈薩克綿羊進行布魯氏菌檢測,在干凈載玻片上取30 μL被檢綿羊血清與等量的虎紅平板抗原混合均勻,在5 min內以陽性血清反應為對照,觀察并記錄結果,被檢血清出現較為明顯的顆粒狀或絮狀凝集則判定為布魯氏菌陽性,否則判定為陰性。以同樣的方法對所有樣品重復檢測2次,以確保檢測結果準確。
1.2.2 基因組DNA的提取與檢測 基因組DNA的提取:取2 mL無明顯血凝塊的新鮮血液,采用酚-氯仿法對其進行基因組DNA提取。
DNA純度與濃度的檢測:采用生物光譜計檢測DNA純度和濃度,用Plate assay法和瓊脂糖凝膠電泳法對DNA驗證的濃度及純度進行驗證,并對高濃度樣品進行適當稀釋,于 -18 ℃ 保存備用。
1.2.3 引物設計 參照綿羊MHC-DQB2的基因序列(GenBank登錄號:EU176819),利用引物設計軟件 Primer Premier 5.0設計該基因exon3的特異引物。引物序列為:F:5′-CCTCAGTGGAACCTACAGTGACC-3′;R:5′-TGCCC TTACTCCACTCCACCG-3′。
1.2.4 PCR擴增 PCR擴增體系為:上下游引物各 0.75 μL,模板3.5 μL,無菌雙蒸水7 μL,2×TaqPCR Master Mix 8 μL,總體積20 μL。PCR擴增程序:94 ℃預變性5 min;94℃變性30 s,55 ℃復性30 s,72 ℃延伸40 s,35 個循環;72 ℃ 延伸8 min;4 ℃保存。PCR產物用1%瓊脂糖凝膠電泳檢測,經凝膠成像系統拍照保存。
1.2.5 SSCP電泳 取3 μL的PCR產物與10 μL的變性上樣緩沖液(去離子甲酰胺9.5 mL;0.3%二甲苯青0.1 mL;0.5 mol/L EDTA 0.4 mL,pH值8.0)充分混勻,于PCR儀 98 ℃ 變性10 min,立即冰浴驟冷至少3 min,低溫恒溫循環系統溫度恒定至9 ℃,取10 μL變性產物上樣于10%的非變性聚丙酰胺凝膠(Acr ∶Bis=39 ∶1),300 V預電泳15 min,而后3 W恒定功率過夜電泳10 h。次日電泳結束后,銀染顯色并在X光片觀片燈上分析帶型并拍照保存。
1.2.6 PCR擴增及測序 經PCR -SSCP分析之后,選取不同基因型樣本,重新進行PCR擴增并電泳檢測后,送至北京六合華大基因公司進行測序。測序結果用DNASTAR軟件進行比對分析。
1.2.7 數據統計及分析 利用Excel整理統計數據,采用χ2適合性檢驗法分別分析病例組與正常組哈薩克羊MHC-DQB2基因exon3多態位點的基因頻率和基因型頻率是否符合Hardy-Weinberg(HWE)平衡定律,P>0.001表示無統計學差異,符合Hardy-Weinberg平衡定律,反之P<0.001 則說明不符合Hardy-Weinberg平衡定律。所有統計學分析均采用SPSS 17.0統計軟件包,通過比較分析每個SNP的基因頻率或基因型頻率分別在病例組和正常組中的差異性,從而分析其與布魯氏菌病易感性的相關性。
1.2.8 單倍型構建 去除等位基因頻率<0.05以及不符合Hardy-Weinberg平衡定律的SNPs位點,選擇同時符合以上2個條件的位點,用SHEsis在線軟件進行單倍型的構建,取連鎖不衡系數(D′值)作為連鎖不平衡指標。
2 結果與分析
2.1 布魯氏菌檢測
采用虎紅平板凝集試驗檢測綿羊種布魯氏菌陽性,結果在100個哈薩克羊個體中檢出布魯氏菌陰性個體84個,陽性個體為16個,布魯氏菌陽性檢出率為16%,檢測結果如圖1所示。
2.2 基因組DNA的提取電泳檢測
哈薩克羊全血中提取的基因組DNA經生物光譜計和Plate assay法檢測,發現基因組DNA濃度在40~1 000 μg/mL。純度經1%的瓊脂糖凝膠電泳檢測,出現整齊、清晰的單一條帶,符合后期試驗要求。
2.3 MHC-DQB2基因的PCR擴增結果檢測
將哈薩克羊MHC-DQB2 exon3 PCR擴增產物于1%的瓊脂糖凝膠檢測,檢測結果與DL 2000 marker比較與預期的相符,因而判斷為目的片段。
2.4 SSCP分析
分別對病例、正常組哈薩克羊個體MHC-DQB2 exon3的PCR擴增產物進行SSCP電泳,部分電泳結果見圖2。由于本試驗應用PCR-SSCP方法具有高度靈敏性,所以出現的基因型比較多,是否還有一些其他未檢測出的基因型,還有待于擴大樣本數量以進一步研究。

2.5 哈薩克羊MHC-DQB2基因exon3 SNPs分析
通過PCR-SSCP技術及測序的序列比對結果分析,哈薩克羊MHC-DQB2基因exon3在282 bp內共發現22個多態位點,經Hardy-Weinberg平衡定律檢驗無差異統計學意義,表明具有群體代表性。這些位點中不存在插入和缺失位點,有1個位點屬于四態性,1個位點屬于三態性,其余20個位點屬于二態性。這些二態性位點中轉換位點有16個,包括8個A/G轉換位點和8個C/T轉換位點;顛換位點有4個,包括A/T顛換位點2個,A/C、G/C顛換位點各1個。相比而言,轉換率為顛換率的4倍,這可能是因為CpG(堿基C、G以及連接兩者的磷酸酯鍵 p)結構上的胞嘧啶殘基是基因組中最易發生突變的位置,大多數是甲基化的,可自發地脫去氨基形成堿基T,因此導致了C→T(互補鏈上G→C)的突變增多,從而導致了轉換位點遠高于顛換位點[12]。
2.6 哈薩克羊MHC-DQB2基因exon3的單氨基酸多態性(SAP)分析
對哈薩克羊DQB2 exon3氨基酸序列進行分析,在93個氨基酸位點中,有20個屬于氨基酸多態位點,占總氨基酸位點數的21.5%。其中,有6個氨基酸突變為有義突變,其余均為無義突變(表1)。
2.7 哈薩克羊MHC-DQB2 exon3多態性與布魯氏菌病易感性的關系
許多疾病的抗性和易感性都與MHC有關聯,作為針對某些疾病的抗性和易感性的候選基因的MHC,已經成為現代分子免疫遺傳的研究熱點之一。本研究對MHC-DQB2基因的多態性與布魯氏菌病易感性進行關聯分析發現,MHC-DQB2可能是與布魯氏菌病易感性相關的基因之一。哈薩克羊MHC-DQB2 exon3 每個SNP的等位基因及基因型頻率在病例組和正常組中的分布差異見表2、表3,其中140 A/C/T/G位點、155T/C的等位基因在病例和正常樣本中的分布存在顯著差異(P<0.05),初步分析認為MHC-DQB2 exon3的140 A/C/T/G、155T/C與綿羊布魯氏菌病易感性相關,相關的研究內容目前尚未見報道。針對該基因每個多態位點的基因型進行關聯分析,發現位點AA/Aa/aa基因型在病例和正常樣本中的分布無顯著差異(P>0.05)。

表1 哈薩克羊MHC-DQB2基因exon3的SNPs位點及氨基酸變異位點

表2 中國美利奴羊MHC-DQB2基因exon3基因頻率
注:“*”表示病例組與正常對照組中國美利奴羊MHC-DQB2相同等位基因間差異顯著(P<0.05)。
2.8 病例和正常組MHC-DQB2 exon3的連鎖不平衡分析及單倍型的構建
對符合Hardy-Weinberg平衡定律的SNPs位點進行連鎖不平衡分析,通過SHSsis在線軟件[13]計算連鎖不平衡系數D′值,觀察各位點之間的連鎖不平衡程度,判斷是否存在連鎖不平衡。在MHC-DQB2 exon3的22個SNPs位點中,最終篩選出符合條件的18個位點進行單倍型的構建,連鎖不平衡圖見圖3。方格內數字就是連鎖不平衡系數D′值乘以100譬如99表示D′=0.99。D′值越大,意味著相關聯的2個SNPs間連鎖不平衡程度越強。通常認為D′>0.9為高度連鎖,D′>0.7 認為2個SNPs位點位于同一個Block區域,可以進行單倍型分析。圖3中各個方塊的顏色由淺至深,表示連鎖程度由低到高。經連鎖不平衡分析發現整個MHC-DQB2 exon3存在7個LD Block(表4),將其依次命名為Block1~Block7。
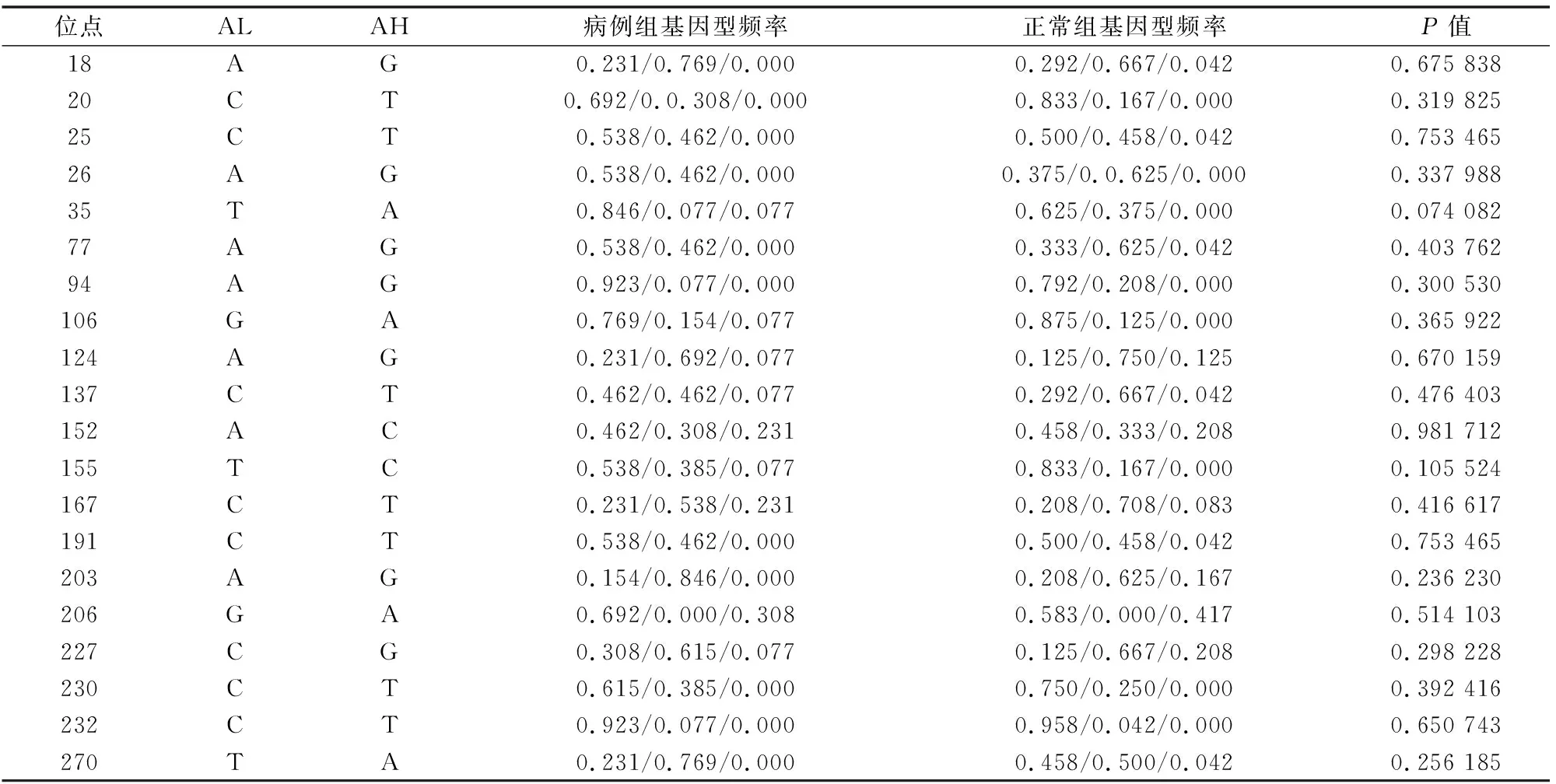
表3 中國美利奴羊MHC-DQB2基因exon3基因型頻率
注:基因型表示為AA/Aa/aa(其中A代表高頻率等位基因;a代表低頻率等位基因); AH代表高頻率基因名稱;AL代表低頻率基因名稱。

由于一個群體中單倍型的頻率不能<0.03,因此單倍型頻率未達到0.03的未參與統計,對應的單倍型組合也應忽略,SHEsis在線軟件在單倍型分析過程中,系統會自動刪除不符合條件的單倍型組合,因而哈薩克羊MHC-DQB2 exon3最終分析了21種單倍型組合(表3、表4),經分析發現在病例組中Hap4、Hap6頻率都顯著低于正常組(P<0.05),這表明這2個單倍型可能與布魯氏菌病抗性有關,或者說具有這2個單倍型的個體對布魯氏菌病不易感,其中,Hap6頻率在病例和正常組中存在極顯著差異(P<0.01);病例組Hap9頻率顯著高于正常組(P<0.05),表明具有這一單倍型的個體可能更易感布魯氏菌病。綜合推測,這2種單倍型可能與布魯氏菌病易感性具有相關性。
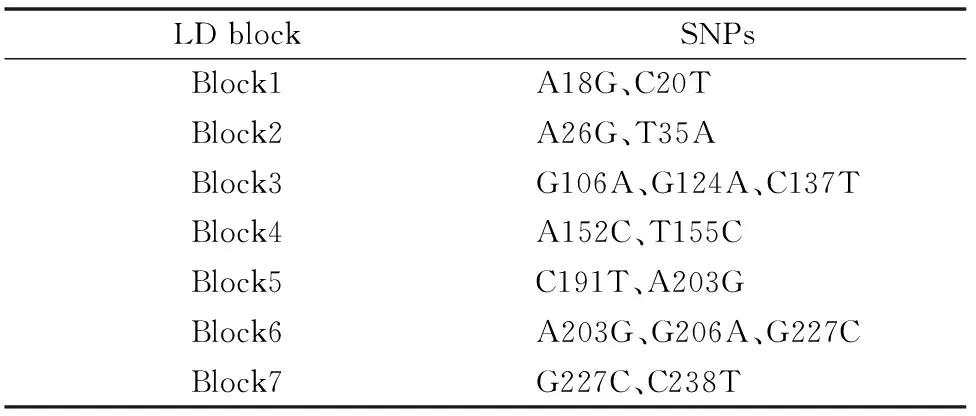
表4 每個連鎖不平衡 block 中 SNPs 組成
3 討論
MHC ClassⅡ區段是一段具有高度多態性的基因,相對于人而言,綿羊的基因多態性在DQ區較之DR區更為豐富。因此,本研究選取了DQB2 exon3作為研究對象,通過PCR-SSCP技術結合測序技術, 對哈薩克羊DQB2 exon3進行了多態性分析,最終發現了22個SNPs位點,而其中只有2個位點在病例和正常組間存在顯著差異;在22個SNPs位點中只有6個位點為單氨基酸多態位點(SAPs),其他位點均為無義突變,且都是由密碼子的簡并性引起的。對比發現,本研究有義突變率較之其他研究成果[14-15]低,這可能是由exon3與exon2所編碼的氨基酸鏈在MHC蛋白上所處的位置及其所發揮的功能不同引起的,也有可能是因為綿羊品種的不同造成的。MHC蛋白的主要功能是抗原遞呈,在機體的免疫系統中發揮著非常重要的作用,與家畜的抗病性和易感性有著密切的關系。本研究通過對MHC-DQB2 exon3多態性與綿羊布魯氏菌病易感性相關性研究,初步篩選了與布魯氏菌病易感性相關的SNPs位點,但由于MHC是一組緊密連鎖的基因群,在一條染色體上的等位基因很少發生同源染色體間的交換,構成了一個單體型(單倍型),在遺傳過程中MHC單體型是以一個完整的遺傳單位由親代傳給子代[16]。因此,本研究對符合要求的SNPs位點構建了單倍型,得到了3種與布魯氏菌病易感性相關的單倍型,其中有2個單倍型在病例和正常組間達到了顯著差異水平(P<0.05),1個達到了極顯著差異水平(P<0.01)。這為今后開展布魯氏菌抗性分子標記輔助選擇研究提供了一定依據,為進一步尋找具有易感性遺傳位點而最終進行抗病育種奠定了基礎,但MHC-DQB2 exon3 的2個SNP位點及3個單倍型能否作為抗布魯氏菌病的遺傳標記,還需要對這些哈薩克羊個體進行攻毒試驗以進一步驗證以及其如何參與綿羊布魯氏菌病的發生和發展的機制仍需進一步研究探討。

表5 病例和正常組MHC-DQB2 exon3單倍型分析
參考文獻:
[1]李振憲. 家畜布魯氏菌病防控現狀及應對策略[J]. 中國畜牧獸醫文摘,2016,32(9):108.
[2]馬曉菁,庫爾班·居麥,舒 展,等. 2012—2014年新疆動物布魯氏菌病流行病學調查與分析[J]. 畜牧與獸醫,2016,48(5):111-114.
[3]陳芳芳,潘 玲,耿照玉,等. MHC分子多態性的起源、演變與抗病機理[J]. 畜牧獸醫學報,2010,41(9):1061-1067.
[4]Hickford J,Zhou H,Fang Q. Haplotype analysis of the DQA genes in sheep:Evidence supporting recombination between the loci[J]. Journal of Animal Science,2007,85(3):577-582.
[5]Herrmann-Hoesing L M,White S N,Kappmeyer L S,et al. Genomic analysis ofOvisaries(Ovar) MHC class IIa loci[J]. Immunogenetics,2008,60(3/4):167-176.
[6]Kenter M,Otting N,Anholts J,et al. Evolutionary relationships among the Primate Mhc-DQA1 and DQA2 alleles[J]. Immunogenetics,1992,36(2):71-78.
[7]Oorschot R V,Maddox J F,Adams L A,et al. Characterization and evolution of ovine MHC classⅡ DQB sequence polymorphism[J]. Animal Genetics,1994,25(6):417-424.
[8]唐一通,肖 娜,李智山,等. 單核苷酸多態性檢測方法研究進展[J]. 現代生物醫學進展,2013,13(27):5393-5397.
[9]馬 超. 單核苷酸多態性與廣州漢族人群前列腺癌的相關性研究[D]. 廣州:南方醫科大學,2014.
[10]楊 俊. 上位作用特征基因的選擇與分類方法研究[D]. 長沙:國防科學技術大學,2011.
[11]Jakobsdottir J,Gorin M B,Conley Y P,et al. Interpretation of genetic association studies:markers with replicated highly significant odds ratios may be poor classifiers[J]. PLoS Genetics,2009,5(2):e1000337.
[12]尚軍亮. 全基因組單核苷酸多態性交互作用研究[D]. 西安:西安電子科技大學,2013.
[13]Shi Y Y,He L. SHEsis,a powerful software platform for analyses of linkage disequilibrium,haplotype construction,and genetic association at polymorphism loci[J]. Cell Research,2005,15(2):97-98.
[14]陳月娥,李建華,茍亞峰,等. 中國美利奴羊MHC-DRB1 exon2單倍型構建及與布魯氏菌病易感性關聯分析[J]. 中國科學(生命科學),2014,44(5):495-501.
[15]王文文,許萬云,胡夢薇,等. 中國美利奴羊OLA-DQB1基因exon2遺傳多態性與布魯氏菌病易感性的關聯分析[J]. 中國畜牧獸醫,2015,42(6):1495-1503.
[16]亢孝珍,額爾敦木圖,姜建強,等. 主要組織相容性復合體(MHC)基因研究進展[J]. 中國畜牧獸醫,2014,41(5):28-33.
猜你喜歡
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
海峽科技與產業(2016年3期)2016-05-17 04:32:12
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
