瀕危植物海南龍血樹的種群結構與動態
2018-06-07 03:08:18楊立榮張治禮嚴武平鄭道君陳加利
生態學報 2018年8期
楊立榮,張治禮,云 勇,嚴武平,陳 宣,張 浪,鄭道君,陳加利
1 海南省農業科學院熱帶園藝研究所,海口 571100 2 海南省農業科學院農業環境與土壤研究所,海口 571100 3 海南大學,海口 570228 4 海南省熱帶特種經濟植物種質資源創新利用重點實驗室,海口 571100
種群通常指某一特定時間和區域內同一物種構成的群體,它們共享同一基因庫或存在潛在隨機交配能力[1]。種群結構是種群的最基本特征,一直以來都是種群生態學研究的核心內容之一,通常通過分析種群的年齡結構、生命表和存活曲線等方法來獲得[2- 3]。種群結構既能反映現在的種群狀態,即現在種群內不同大小個體數量的分布和配置狀況,也能反映植物與環境之間的適合度,分析過去種群結構與受干擾情況,預測未來的種群動態和演變趨勢[4- 6]。因此,研究種群的結構及其動態,有助于人們更好地認識種群的生態特性、穩定性及發展動態等,對瀕危種群的更新復壯及其所在群落和生態系統的恢復具有重要意義。
海南龍血樹(Dracaenacambodiana)(小花龍血樹、柬埔寨龍血樹),屬百合科龍血樹屬的單子葉植物[7- 8],國家二級重點保護植物和稀有瀕危保護植物[9- 10]。海南龍血樹是一種生長緩慢,耐干旱的喜陽植物,目前分布于海南島西南部的內陸山區及南部沿海地區,生長于裸露花崗巖和石灰巖的石縫殘積土中或緊貼石壁生長于砂壤土中,屬于典型的巖石伴生植物[11- 12]。由于海南龍血樹具有較高的藥用價值和觀賞價值[13- 15],野生資源受到掠奪性采挖,野生資源已十分匱乏,種群處于瀕危狀態。目前關于海南龍血樹的研究主要集中在組織快繁、栽培技術、以血竭為基礎的基礎性研究和開發利用以及分子生物學等方面[16- 19]。此外,鄭道君等人對海南龍血樹野生資源分布、種群更新情況及種子萌發進行了研究[12, 20- 21]。近年來,海南龍血樹的種群結構和分布格局也有報道,但周文嵩等人僅對昌江地區的海南龍血樹種群結構進行了研究[22],李偉杰等人涉及的種群雖多,但僅對海南龍血樹種群的年齡結構和分布格局進行了研究,并未對海南龍血樹種群的靜態生命表、數量動態特征和生存狀況進行分析[23]。本文擬以處于不同代表性生境的海南龍血樹種群為對象,對它們的年齡結構、數量動態特征與生存狀況進行定量研究,預測種群未來的動態趨勢,以期從種群生態學方面為海南龍血樹種群的保護和恢復提供依據。
1 研究方法
1.1 野外調查
在查閱文獻資料的基礎上,采用踏查法對海南龍血樹種群進行實地調查。海南龍血樹主要分布在石灰巖和花崗巖兩種生境中,因此,本文對兩種生境的11個種群進行了調查。其中,石灰巖生境調查了東方市俄賢嶺的2個種群;花崗巖生境調查了分布于海邊的三亞南山和亞龍灣種群、萬寧樂南山種群、昌江昌化嶺種群,以及分布于非海邊的三架嶺種群、東方黑嶺種群和樂東尖峰嶺的3個種群(表1)。各種群的地理位置如圖1所示。采用路線調查法調查種群內的海南龍血樹個體,記錄個體的株型、基徑、樹高及冠幅。
1.2 種群結構分析
1.2.1 種群徑級劃分
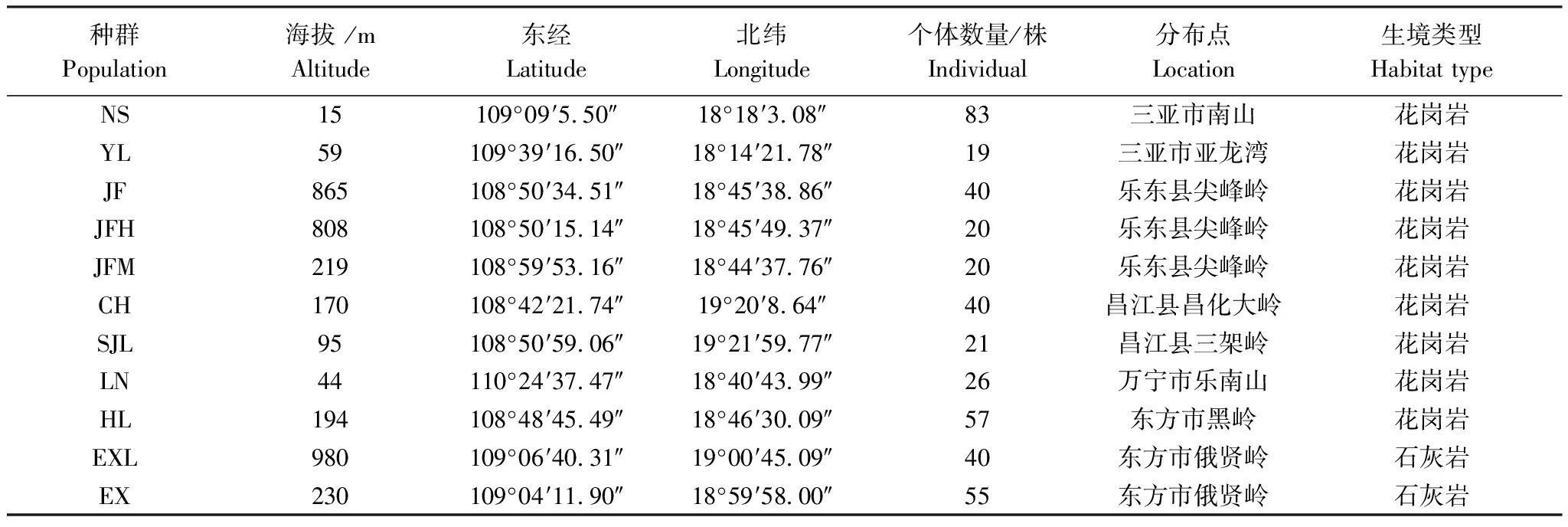
表1 海南龍血樹種群概況
NS: 三亞市南山居群, Nanshan population in Sanya City; YL: 三亞市亞龍灣居群, Yalongwan population in Sanya City; JF: 樂東縣尖峰嶺引水頭居群, Yinshuitou population on Jianfengling in Ledong County; JFH: 樂東縣尖峰嶺回頭灣居群, Huitouwan population on Jianfengling in Ledong County; JFM: 樂東縣尖峰嶺叉河口居群, Chahekou population on Jianfengling in Ledong County; CH: 昌江縣昌化大嶺居群, Changhuadaling population in Changjiang County; SJL: 昌江縣三架嶺居群, Sanjialing population in Changjiang County; LN: 萬寧市樂南山居群, Lenanshan population in Wanning City; HL: 東方市黑嶺居群, Heiling population in Dongfang City; EXL: 東方市俄賢嶺居群, Exianling population in Dongfang City; EX: 東方市俄賢嶺南浪村居群, Nanlang population on Exianling in Dongfang City
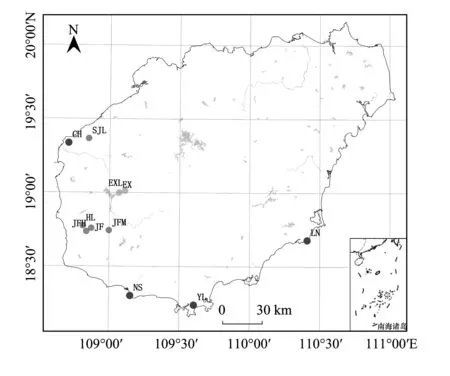
圖1 海南龍血樹種群的地理位置Fig.1 Populations′ location of Dracaena cambodiana NS: 三亞市南山居群, Nanshan population in Sanya City; YL: 三亞市亞龍灣居群, Yalongwan population in Sanya City; JF: 樂東縣尖峰嶺引水頭居群, Yinshuitou population on Jianfengling in Ledong County; JFH: 樂東縣尖峰嶺回頭灣居群, Huitouwan population on Jianfengling in Ledong County; JFM: 樂東縣尖峰嶺叉河口居群, Chahekou population on Jianfengling in Ledong County; CH: 昌江縣昌化大嶺居群, Changhuadaling population in Changjiang County; SJL: 昌江縣三架嶺居群, Sanjialing population in Changjiang County; LN: 萬寧市樂南山居群, Lenanshan population in Wanning City; HL: 東方市黑嶺居群, Heiling population in Dongfang City; EXL: 東方市俄賢嶺居群, Exianling population in Dongfang City; EX: 東方市俄賢嶺南浪村居群, Nanlang population on Exianling in Dongfang City
在同一生境下同一樹種的齡級和徑級對環境的反應規律具有一致性[24]。因此,采用空間推時間的方法,用徑級代替齡級分析海南龍血樹種群的結構[2, 25]。根據生活史特征和野外調查數據的特點,將海南龍血樹種群劃分為13個徑級,每個徑級對應相應的齡級。基徑(以下用D表示)≤3cm為第Ⅰ齡級,3cm
1.2.2 種群年齡結構及動態
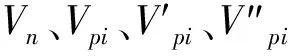
種群齡級間動態指數(Vn)為:
式中:Sn與Sn+1分別為第n和第n+1年齡級的個體數,-1≤Vn≤1。
種群年齡結構的數量變化動態指數(Vpi)為:
式中:K為種群年齡級數量,此值越大,說明種群的增長趨勢越大。

此值越大,說明種群抗干擾能力越大,種群穩定性越好。
1.2.3 靜態生命表和存活曲線
根據特定時間生命表的編制方法[27],編制海南龍血樹種群的靜態生命表。靜態生命表包含:x為年齡級;Ax為x齡級內現有個體數;由于靜態生命表是用同一時期收集到的種群中的所有個體的徑階編制而成,它只反映了多個世代重疊的年齡動態歷程中的一個特定時間,而不是對種群全部生活史的追蹤[28- 29],因此,生命表的編制中會出現死亡率為負值的情況,為此,本文采取文獻中的勻滑技術對數據Ax進行處理[27, 30- 31],ax即為Ax勻滑修正后的數據;lx為x齡級開始時標準化存活個體數;dx為從x到x+1齡級間隔期內標準化死亡個體數;qx為從x到x+1齡級間隔期間的死亡率;Lx為從從x到x+1齡級間隔期間還存活的個體數;Tx為從x齡級到超過x齡級的個體總數;ex為進入x齡級個體的生命期望或平均期望壽命;Kx為消失率。通過實測Ax值,計算以上各項的值,它們的計算公式分別為:
1.2.4 種群生存分析
種群生存分析采用種群生存率函數(S(i));累計死亡率函數(F(i));死亡率密度函數(f(ti))和危險率函數(λ(ti))[30- 31]。計算公式如下:

2 結果與分析
2.1 種群年齡結構
分析結果表明,海南龍血樹種群的年齡結構與其所處的生境關系密切。NS、YL、CH和LN種群分布在海邊的花崗巖生境,它們的特點是:種群中均存在一定比例的中齡、成齡和老齡個體。三亞NS種群和YL種群的年齡結構中均缺少幼齡個體,幼齡個體占種群個體總數的比例分別為0和5%;中、成齡個體所占的比例較高,分別為61%和90%;老齡個體占有一定的比例,分別為39%和5%。由于幼齡個體的極度缺乏,NS和YL種群結構均屬于衰退型。其中,NS種群幼齡個體缺失,且擁有更高比例的老齡個體。因此,NS種群的衰退程度比YL種群更大。位于萬寧樂南山的LN種群幼齡、中齡、成齡和老齡個體所占的比例分別為27%、46%、19%和8%,屬于穩定型種群。同樣,位于昌化大嶺的CH種群幼、中、成、老齡的個體數量占種群個體總數量的比例分別為30%、25%、25%和20%,其種群年齡結構也屬于穩定型種群。相比之下,CH種群幼、中、成、老齡的個體數量分布較LN種群均勻,種群的穩定性更好些,而LN種群雖有老齡個體,但其老齡個體比例僅為8%,偏向于增長型(圖2)。

圖2 海南龍血樹種群的年齡結構Fig.2 Age structure of Dracaena cambodiana populationsⅠ: D(個體基徑, basal diameter of individuals)≤3cm; Ⅱ: 3cm
分布于花崗巖山間斜坡上的種群包括尖峰嶺地區的JF、JFH、JFM和HL種群,以及昌江三架嶺的SJL種群,它們的年齡結構無老齡個體,成齡的個體極少,種群中個體集中分布在幼、中齡,種群為增長型。其中,JF種群的幼、中齡比例為53%和42%,成齡的個體僅為5%;HL種群幼、中齡比例為33%和60%,成齡的個體僅為7%;JFH和JFM幼、中齡比例均為100%。
位于石灰巖俄賢嶺地區的EXL和EX種群與花崗巖非海邊種群的年齡結構類似,無老齡個體,種群中個體集中分布在幼、中齡,種群也屬于增長型。但它們沒有成齡個體,中齡個體也較少,種群個體主要集中在幼齡階段,如,EXL的幼齡個體占95%,中齡僅有5%;EX幼齡個體占67%。
2.2 種群年齡結構的動態變化分析
基于種群動態量化分析方法,對海南龍血樹11個種群相鄰齡級間個體數量變化動態及各種群整體數量變化動態進行分析(表2,表3)。結果顯示,海南龍血樹各種群齡級間的個體數量均有明顯的波動過程,各種群的結構并不穩定,11個種群在齡級間多出現了負增長動態,出現齡級間衰退現象的次數在1/4以上,最高達到3/5(JFH)。Ⅰ-Ⅳ齡級在大部分種群中存在,但是10個種群(NS不存在Ⅰ和Ⅱ齡級)中,有6個種群在Ⅰ和Ⅱ齡級之間出現衰退現象,表明Ⅰ向Ⅱ齡級時有嚴重的環境篩,導致Ⅰ向Ⅱ齡級過渡時極易出現高死亡率;Ⅲ和Ⅳ齡級之間的衰退現象較少,9個種群(EX和EXL沒有Ⅲ齡級)中僅2個種群出現負增長,表明Ⅲ向Ⅳ齡級過渡時環境條件較為合適,種群穩定。此外,有6個種群存在Ⅵ向Ⅶ齡級過渡,其中有5個種群的年齡結構動態指數等于或大于零,僅NS的為負數,且僅為11.11%,表明這個階段也是較為穩定的。
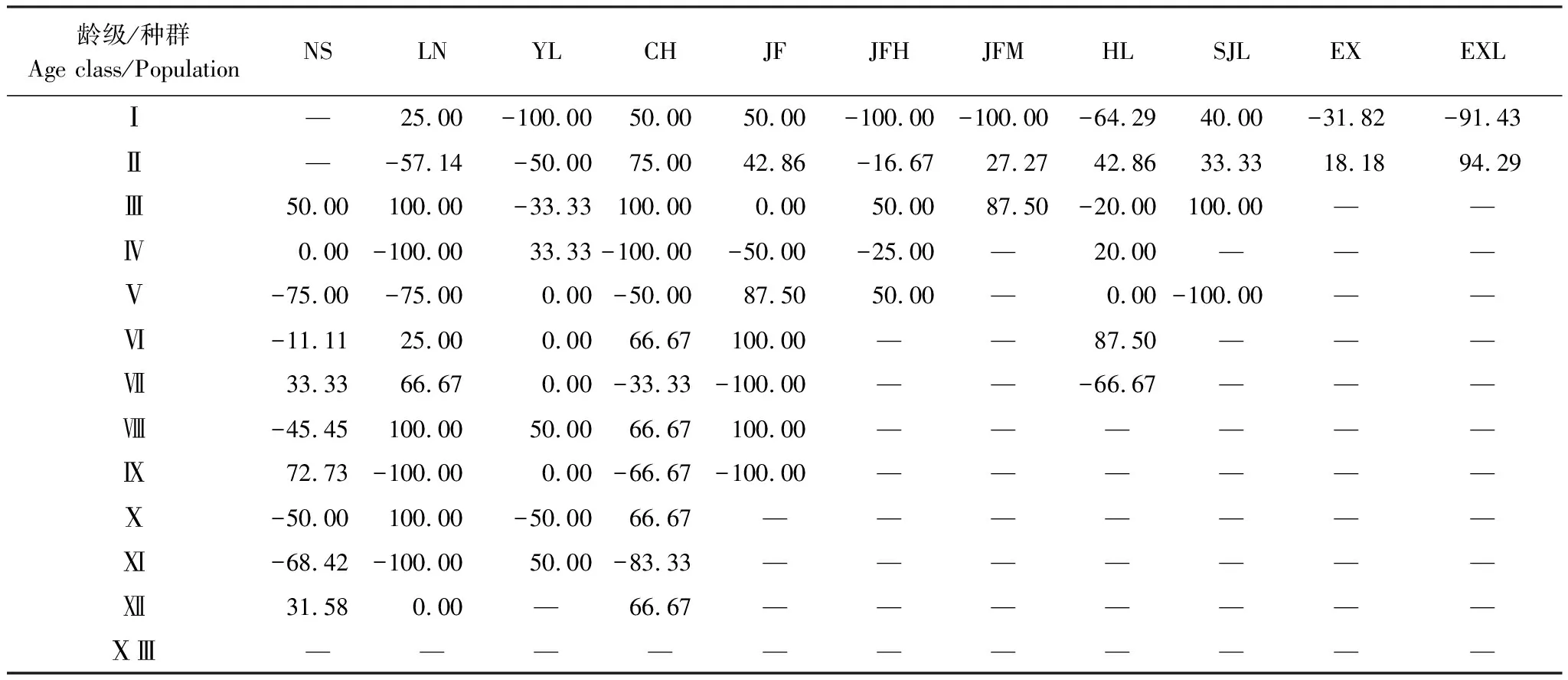
表2 海南龍血樹種群的年齡結構動態指數(Vn/%)
Ⅰ:D(個體基徑, basal diameter of individuals)≤3cm; Ⅱ: 3cm
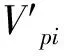
2.3 靜態生命表及存活曲線
海南龍血樹各種群的靜態生命表如表4所示,以齡級為x軸,以lnlx、qx、kx為y軸繪制各種群的存活曲線、死亡率曲線和消失率曲線如圖3和圖4所示。種群存活曲線是對種群的死亡過程的直觀描述,由于勻滑處理會掩蓋種群數量波動中的某些生態現象[32],因此,本文將勻滑處理前后的種群存活曲線進行對比。
表3 海南龍血樹種群的動態指數
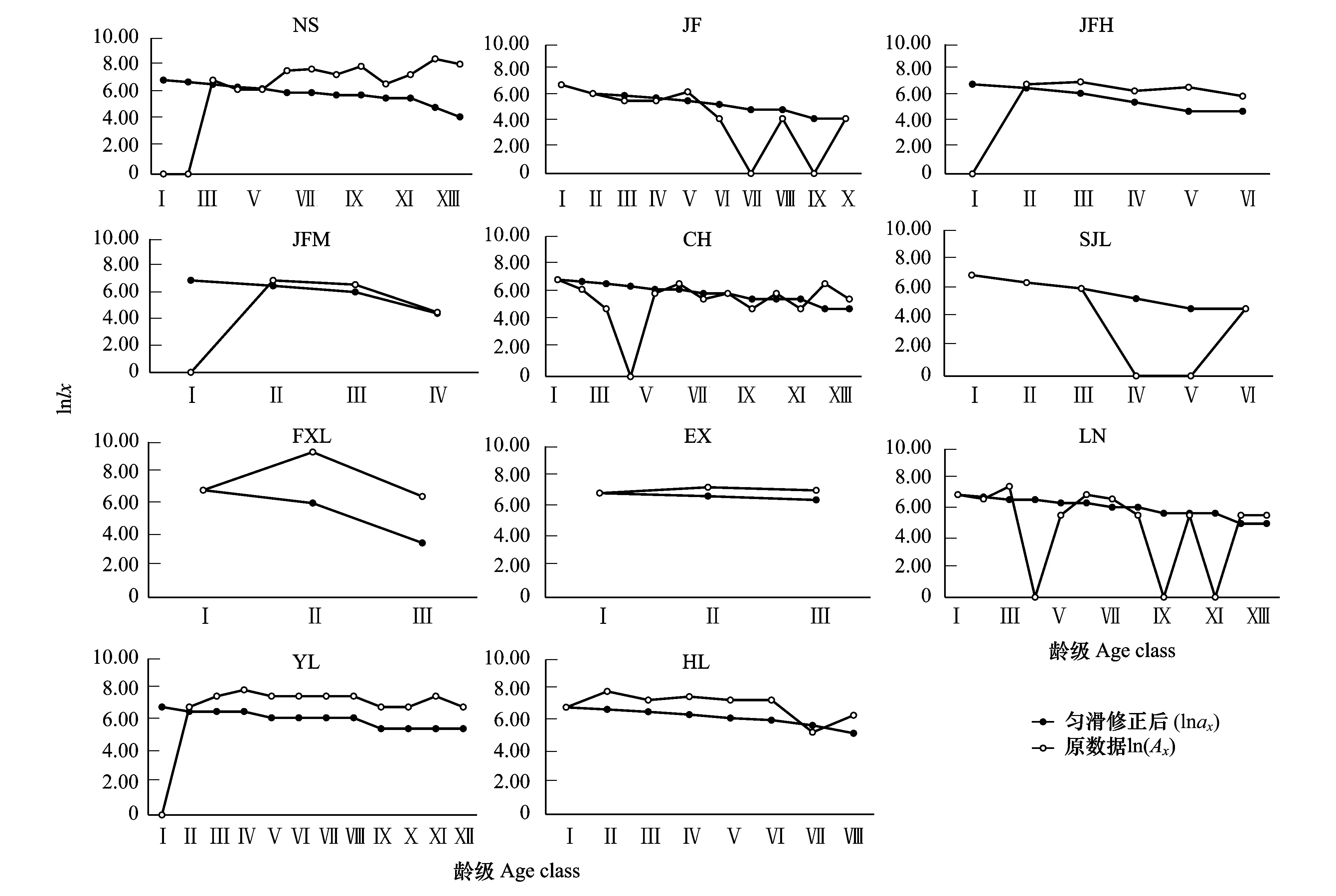
圖3 海南龍血樹種群的存活曲線Fig.3 Survival curve of Dracaena cambodiana populations
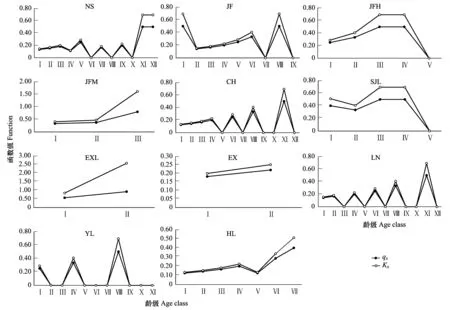
圖4 海南龍血樹種群的死亡率(qx)和消失率(Kx)Fig.4 Mortality and killing power of Dracaena cambodiana populations
從存活曲線看,11個海南龍血樹種群均屬于Deevey-Ⅰ型,呈現凸型,生命期望值ex隨著齡級的增長而呈現不斷下降的趨勢,表明海南龍血樹的生存能力隨著年齡的增加而下降,而且其生命期望值通常在老齡階段的下降趨勢更顯著,種群幼齡和中幼齡個體的生命期望通常相差不大,說明海南龍血樹幼齡及中幼齡個體的生存能力較好,接近老齡時個體生存能力較差。圖4也顯示海南龍血樹種群幼齡個體的存活率高于中齡個體和老齡個體,幼齡個體的死亡率和消失率小于中、老齡個體(圖3)。
2.4 種群生存分析
NS、YL、CH、EX、LN和HL種群累計死亡率的起始值較低,函數值≤0.2;JF、JFH、JFM和SJL種群累計死亡率的起始值較高,函數值在0.2—0.4之間;EXL種群累計死亡率的起始值最高,函數值為0.67,表明EXL種群所處的生境對齡級Ⅰ個體的環境篩作用較強。NS、JF、JFH、JFM、SJL、EXL種群的累計死亡曲線呈現出前期變化幅度大,中后期變化幅度變小的特點;YL的累計死亡曲線呈現出波動性變化,Ⅰ到Ⅱ齡級、Ⅳ到Ⅵ齡級、Ⅷ到Ⅹ齡級的死亡累計率無變化,其余各齡級間的變化幅度基本一致;CH、EX、LN、HL種群的累計死亡曲線呈現出早中晚齡期變化幅度基本一致的特點(圖5)。
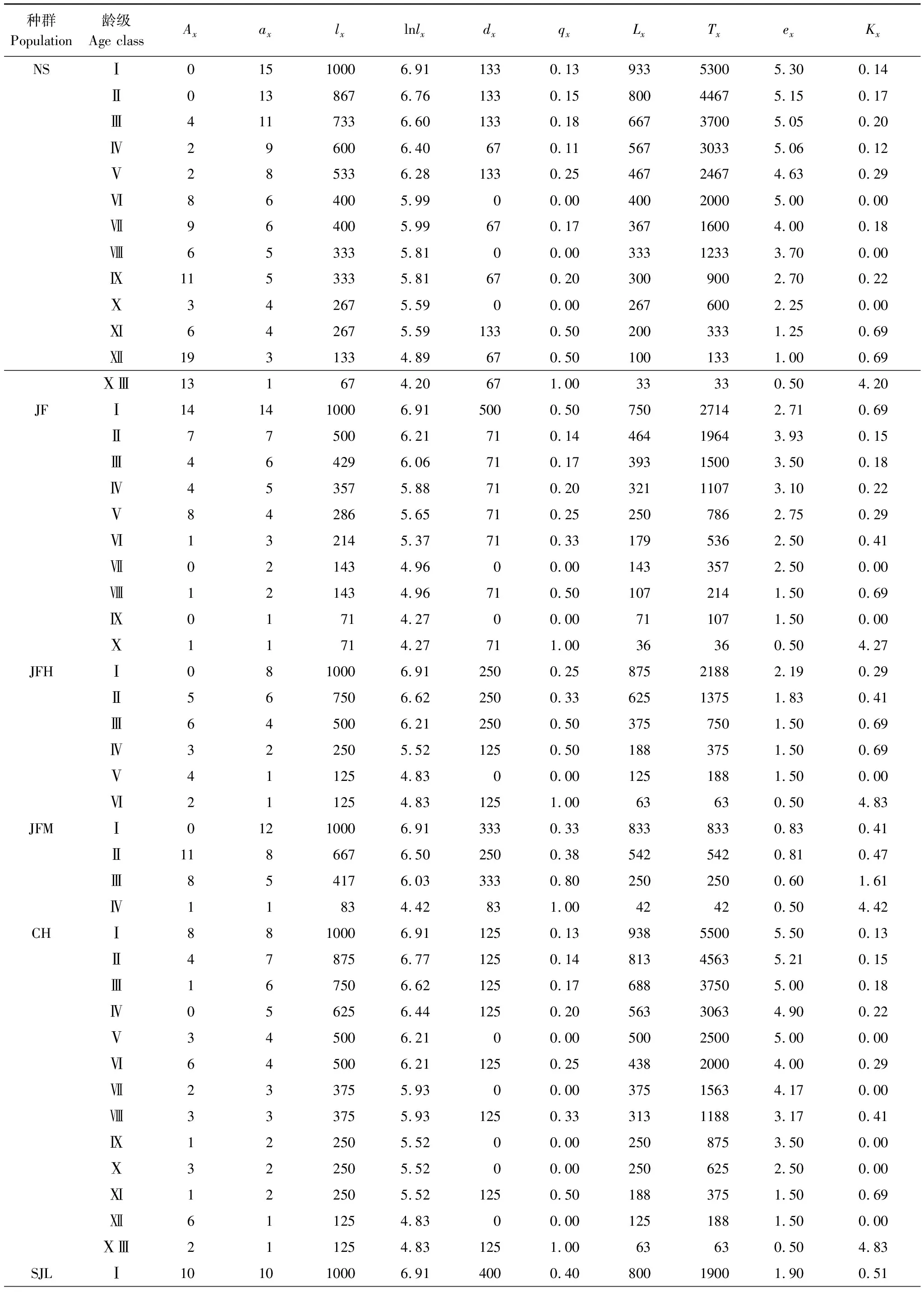
表4 海南龍血樹種群的靜態生命表
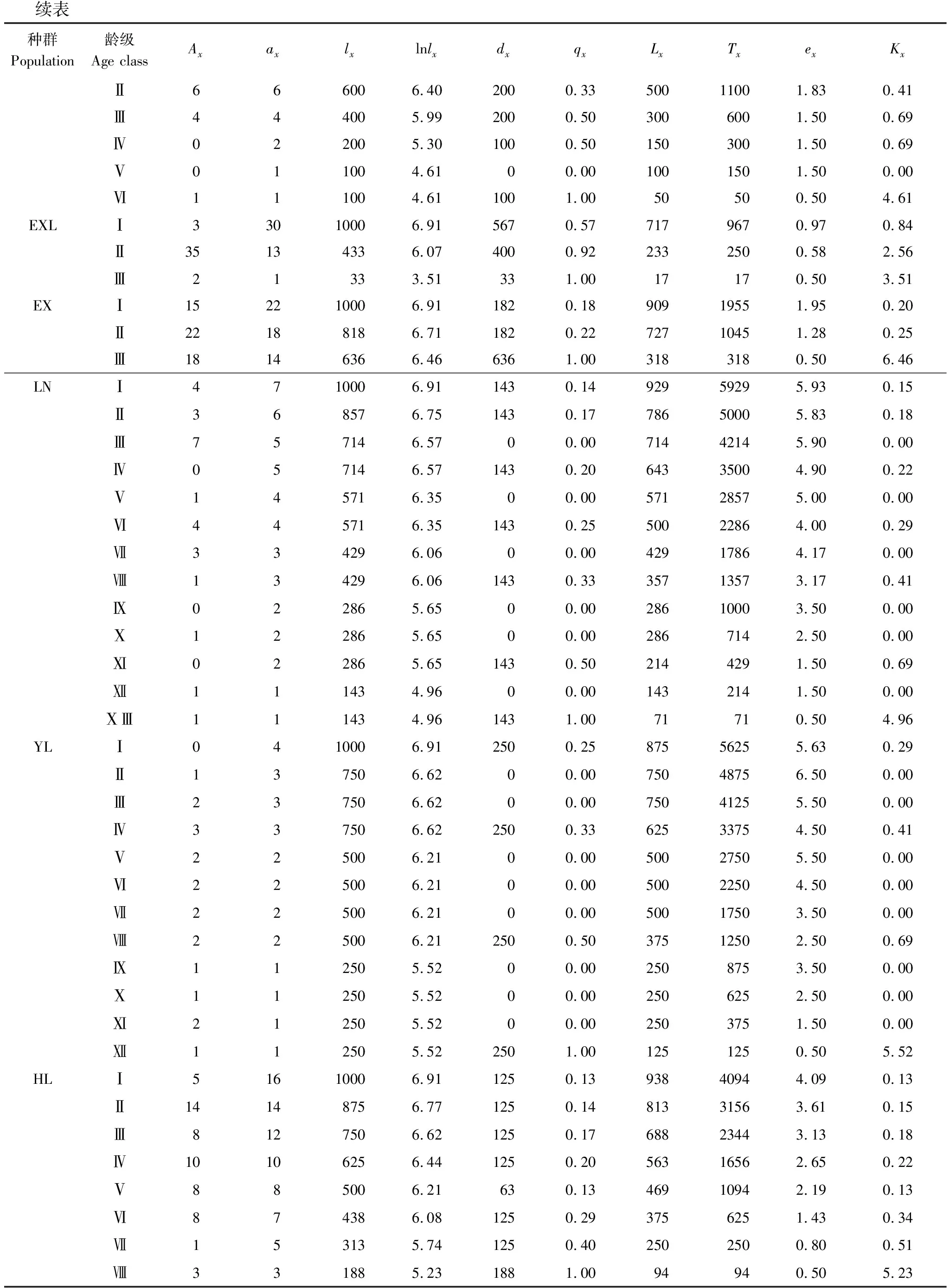
續表種群Population齡級Age classAxaxlxlnlxdxqxLxTxexKxⅡ66600 6.40 200 0.33 500 1100 1.83 0.41 Ⅲ44400 5.99 200 0.50 300 600 1.50 0.69 Ⅳ02200 5.30 100 0.50 150 300 1.50 0.69 Ⅴ01100 4.61 0 0.00 100 150 1.50 0.00 Ⅵ11100 4.61 100 1.00 50 50 0.50 4.61 EXLⅠ3 30 10006.91 5670.577179670.970.84 Ⅱ3513433 6.07 400 0.92 233 250 0.58 2.56 Ⅲ2133 3.51 33 1.00 17 17 0.50 3.51 EXⅠ15 22 10006.91 1820.1890919551.950.20 Ⅱ2218818 6.71 182 0.22 727 1045 1.28 0.25 Ⅲ1814636 6.46 636 1.00 318 318 0.50 6.46 LNⅠ4 7 10006.91 1430.1492959295.930.15 Ⅱ36857 6.75 143 0.17 786 5000 5.83 0.18 Ⅲ75714 6.57 0 0.00 714 4214 5.90 0.00 Ⅳ05714 6.57 143 0.20 643 3500 4.90 0.22 Ⅴ14571 6.35 0 0.00 571 2857 5.00 0.00 Ⅵ44571 6.35 143 0.25 500 2286 4.00 0.29 Ⅶ33429 6.06 0 0.00 429 1786 4.17 0.00 Ⅷ13429 6.06 143 0.33 357 1357 3.17 0.41 Ⅸ02286 5.65 0 0.00 286 1000 3.50 0.00 Ⅹ12286 5.65 0 0.00 286 714 2.50 0.00 Ⅺ02286 5.65 143 0.50 214 429 1.50 0.69 Ⅻ11143 4.96 0 0.00 143 214 1.50 0.00 ⅩⅢ11143 4.96 143 1.00 71 71 0.50 4.96 YLⅠ0 4 10006.91 2500.2587556255.630.29 Ⅱ13750 6.62 0 0.00 750 4875 6.50 0.00 Ⅲ23750 6.62 0 0.00 750 4125 5.50 0.00 Ⅳ33750 6.62 250 0.33 625 3375 4.50 0.41 Ⅴ22500 6.21 0 0.00 500 2750 5.50 0.00 Ⅵ22500 6.21 0 0.00 500 2250 4.50 0.00 Ⅶ22500 6.21 0 0.00 500 1750 3.50 0.00 Ⅷ22500 6.21 250 0.50 375 1250 2.50 0.69 Ⅸ11250 5.52 0 0.00 250 875 3.50 0.00 Ⅹ11250 5.52 0 0.00 250 625 2.50 0.00 Ⅺ21250 5.52 0 0.00 250 375 1.50 0.00 Ⅻ11250 5.52 250 1.00 125 125 0.50 5.52 HLⅠ5 16 10006.91 1250.1393840944.090.13 Ⅱ1414875 6.77 125 0.14 813 3156 3.61 0.15 Ⅲ812750 6.62 125 0.17 688 2344 3.13 0.18 Ⅳ1010625 6.44 125 0.20 563 1656 2.65 0.22 Ⅴ88500 6.21 63 0.13 469 1094 2.19 0.13 Ⅵ87438 6.08 125 0.29 375 625 1.43 0.34 Ⅶ15313 5.74 125 0.40 250 250 0.80 0.51 Ⅷ33188 5.23 188 1.00 94 94 0.50 5.23
Ax:x齡級內現有的個體數, individual number of age classx;ax:Ax勻滑修正后的數據, the revised data ofAx;lx:x齡級開始時標準化存活個體數, the standardized number of surviving individuals of age classx;lnlx:lx的自然對數,the natural logarithm oflx;dx:從x到x+1齡級間隔期內標準化死亡個體數, the standardized number of death individuals from age classxto age classx+1;qx:從x到x+1齡級間隔期間的死亡率, mortality from age classxto age classx+1;Lx:從x到x+1齡級間隔期間還存活的個體數, the number of surviving individuals from age classxto age classx+1;Tx:從x齡級到超過x齡級的個體總數, the total individual number of age classxand age classes older thanx;ex:進入x齡級個體的生命期望, life expectancy of individuals in the age classx;Kx:消失率, disappearance rate
各種群的死亡密度函數曲線和危險率動態基本相似,在Ⅰ齡級向Ⅱ齡級過渡過程中變化幅度較大,后面各齡級間的變化趨勢趨于平穩,除EXL種群外,其他種群的危險率函數呈現先減小后增大的趨勢,說明海南龍血樹種群在整個生活史中中期穩定,早期和晚期較為脆弱(圖6)。
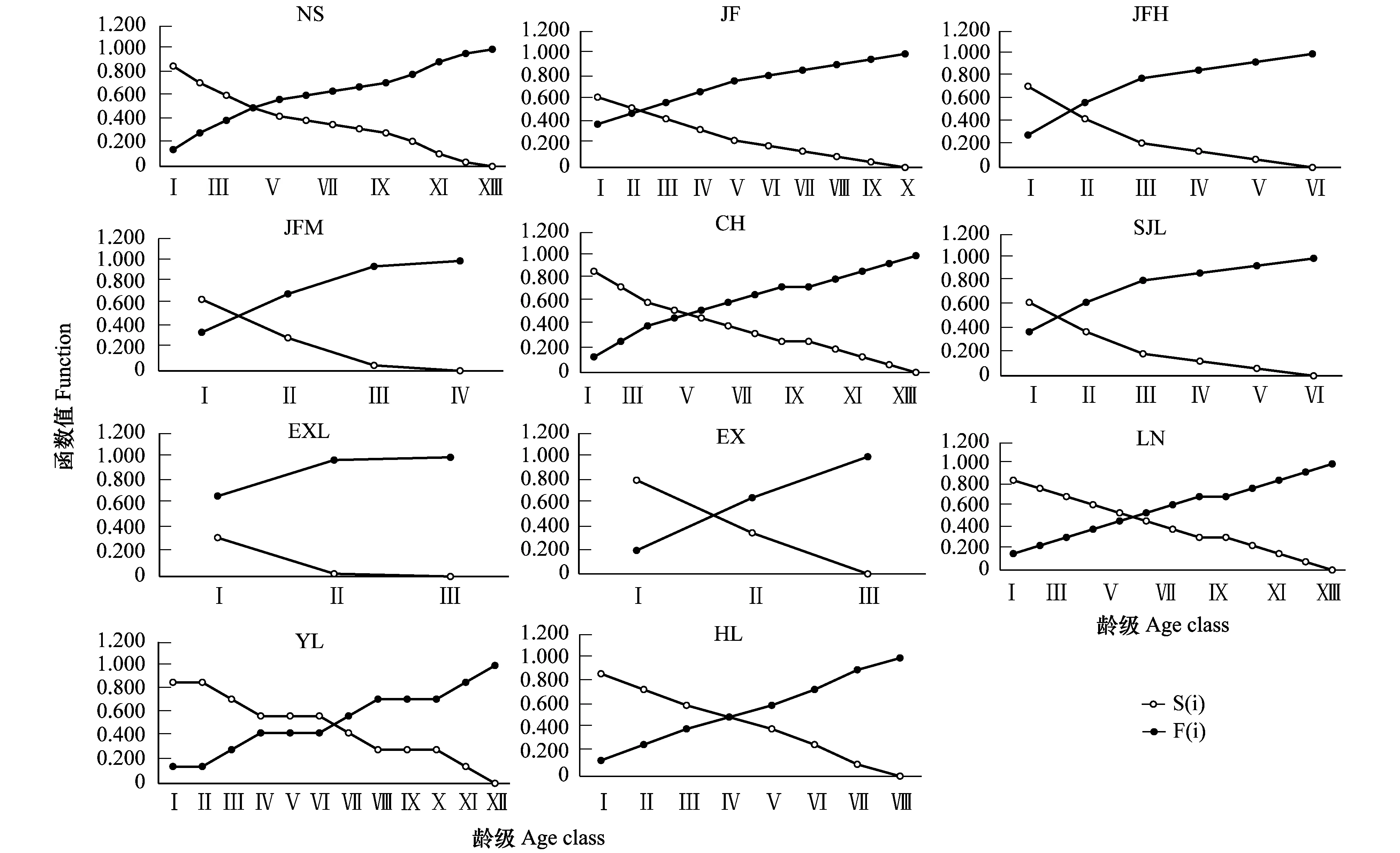
圖5 海南龍血樹種群的生存率(S(i))和累計死亡率(F(i))Fig.5 Survival rate and accumulated mortality rate curve of Dracaena cambodiana populations
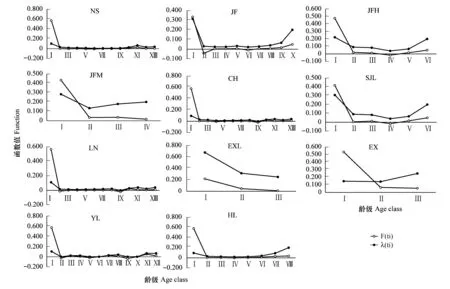
圖6 海南龍血樹種群的死亡密度函數(F(ti))和危險率(λ(ti))Fig.6 Morality density and hazard rate curve of Dracaena cambodiana populations
3 討論
3.1 海南龍血樹種群結構及生存現狀
植物種群齡級結構是其生物學特征與環境因素共同作用的結果[33-34]。李偉杰等人[23]對昌江昌化嶺、三亞大小洞天、萬寧東澳鎮和東方江邊鄉及天安鄉海南龍血樹種群的年齡結構進行分析后認為這些種群的結構均為衰退型;周文嵩等人[22]對昌江地區的海南龍血樹種群年齡結構進行分析后認為昌江地區的海南龍血樹種群也為衰退型。本文對海南龍血樹不同生境下11個種群的年齡結構及種群結構動態指數分析的綜合結果顯示,海南龍血樹種群結構與種群所處的生境關系密切,位于海邊的NS和YL種群結構為衰退型,CH、LN種群為穩定型,而位于非海邊生境的其他7個種群為增長型。這一結果與以上兩人的研究結果有一定差異。造成這種差異的原因可能是齡級劃分標準、調查研究區域和研究方法的不同。由于海南龍血樹種群中的很多個體都沒有明顯的主干,無法獲得個體胸徑,因此本研究中未用胸徑,而是采用基徑劃分海南龍血樹的個體年齡級別,而且將D≤10cm的個體劃分為幼齡個體,這一劃分標準可能將更多的海南龍血樹個體劃分成了幼齡個體。本研究調查的區域更廣,處于尖峰嶺保護區內的種群有4個,在海拔800m以上的種群有2個,它們不易受到人為干擾和破壞,跟其他種群相比,種群中的幼齡個體更多。本研究在尖峰嶺JF種群中發現了較多數量的實生小苗,這是在其他種群中沒有的現象。此外,本研究用年齡結構和種群結構動態指數兩種方法來綜合分析海南龍血樹種群的結構,比僅用單一的年齡結構分析可能更準確。
缺乏可更新的幼齡個體將導致種群趨于衰退[35]。海南龍血樹的自然更新方式有種子更新、根蘗更新和樁蘗更新3種方式,其中后兩種屬于無性繁殖;目前的研究均顯示海南龍血樹種群中極少見到實生苗,種群更新不良,其更新方式以無性繁殖為主[20,23]。本研究結果與前人的基本一致,在調查到的11個海南龍血樹種群中,僅在JF、LN和SJL種群記錄到2年生左右的實生苗,其他種群中極少見到實生苗;在CH種群中發現的幼苗均為根蘗苗或樁蘗苗,是被砍伐后殘留的海南龍血樹根或樁上萌生的幼苗。海南龍血樹種子萌發和幼苗生長對生境的依賴性較高,需要一定的遮蔭環境,但遮蔭度較好的NS和YL種群中沒有發現幼苗的存在,且幼齡個體都極度缺乏,導致NS和YL種群成為衰退種群。在調查NS和YL種群時正值海南龍血樹的結果期,但調查過程中僅發現已枯萎的花序,并未見到其果實,因此推測是否是因為種群個體無法結實才導致NS和YL種群缺少幼苗和幼齡個體而成為衰退種群。這一推測還有待于繁殖生態學方面的研究來驗證。JF、JFM、JFH、HL、EXL、EX和SJL種群僅有前3齡級的個體或缺少Ⅶ齡級以后的中老齡個體,說明這7個種群的形成時間較短,是正在發展的種群。
3.2 海南龍血樹種群的動態趨勢
生命表是判斷植物種群發展趨勢的重要指標,通過建立種群靜態生命表、繪制存活曲線及生存函數曲線可以對海南龍血樹種群動態進行分析。種群靜態生命表和繪制存活曲線分析結果顯示11個海南龍血樹種群的存活曲線均屬于Deevey-Ⅰ型,海南龍血樹幼齡個體的生存能力較好,存活率高于中齡個體和老齡個體,老齡個體生存能力較差;但生存函數分析的結果顯示海南龍血樹種群生活史早期和晚期脆弱,中期穩定,這說明海南龍血樹種群所處的生境更適合成年海南龍血樹的生存,雖然幼齡樹自身的生存能力較強,但現有的生境對幼齡個體的環境篩作用較強,對幼齡樹的生存不利,這一點與鄭道君等人[21]的研究結果一致。
植物種群的發展是一個漫長而復雜的過程,涉及到物種內和物種間的競爭,還涉及到物種與無機環境之間的適應過程。海南龍血樹在種群發展的漫長過程中,適應了嚴酷的巖石生境,對種群分布及種群結構與動態的綜合分析后可以看出,位于海邊生境的海南龍血樹種群是古老種群,處于穩定發展中或正趨于衰退;而位于非海邊的種群則是年輕種群,是增長型種群。由此推斷海南龍血樹種群正在由海邊的巖石生境向非海邊的巖石生境轉移,這一推測還需要分子生物學和生物地理學方面的研究結果來驗證。
3.3 海南龍血樹種群的保護與恢復措施
根據種群聚集程度的不同,物種可分為補償物種、中性物種和退償物種三個類別,補償物種不易滅絕,退償物種容易滅絕,當退償物種的種群數量小于該物種的最小存活種群K0時,該物種趨于滅絕[36]。海南龍血樹在石灰巖生境中為集群分布,在花崗巖生境中為均勻分布,已被列入中國珍稀瀕危保護植物名錄,應屬于退償物種。人類對海南龍血樹野生資源的掠奪性采挖和對其生境的破壞造成海南龍血樹野生資源的分布范圍縮減,分布點數量變少。從目前的文獻和本研究的調查結果來看,野生海南龍血樹種群的分布點不會超過20個,而且多數種群內的個體數量很少,在100株以下;目前種群估測數量能在500株以上的僅有三亞的大小洞天、昌江的昌化大嶺和萬寧的大洲島,其中大洲島的海南龍血樹種群是目前發現的保存最為完好的種群,具有重要的科學研究價值。
綜上所述,為了防止海南龍血樹這一物種的滅絕,海南龍血樹種群的保護工作刻不容緩。對瀕危植物的保護目前主要有三種方法:就地保護、遷地保護和利用現代生物技術的保護[37]。近年來,隨著對極小種群野生植物拯救保護工程的進行,一種新的野生植物保護方法“近地保護”被提了出來,即在物種現有分布區(點) 范圍內的附近,選擇與其相同氣候和相似的生境建立人工保護點[38]。此方法經中科院西雙版納熱帶植物園的比較論證,認為它對稀有、瀕危植物的保護,大大優于通常的遷地保護,是一種行之有效的新保護方法[39]。在就地保護中,對于生活史中存在薄弱環節的瀕危物種種群,可以采用人工輔助措施,如采用實驗室輔助種子萌發,待幼苗長至一定大小時,再移植回原種群[40]。
根據本文的分析結果,對海南龍血樹種群的保護應采取多種保護方法相結合的方式。對于缺少幼齡個體的種群可以采用人工輔助措施獲得該種群的實生幼苗,然后將幼苗移植回該種群,如NS、YL和CH種群可以采取此方式。對于處于易受人為干擾生境下的種群,可以根據種群的情況選擇遷地保護或近原地保護方式保護這類種群的遺傳多樣性,如LN、EX、SJL種群可以采取此方式。而對于已在保護區內海南龍血樹種群,如EXL、JF、JFH、JFM、HL種群,可以根據種群的實際情況,利用人工輔助措施促進種群的天然更新能力,如擾動土壤以促進種子落入土壤、伐除母樹和幼苗周圍的喬灌木、增加土壤養分等,可以促進母樹的結實,提高幼苗的生長量,增加幼苗的密度。加強對海南龍血樹遺傳多樣性的研究,明確其瀕危的內在原因對海南龍血樹種群的保護極其重要。建立異地種質資源圃來保存現存海南龍血樹的核心種質,以防現存種群因人為或自然原因而突然消失從而造成海南龍血樹遺傳多樣性進一步丟失。
參考文獻(References):
[1] 姜漢僑, 段昌群, 楊樹華, 王崇云, 蘇文華. 植物生態學(第二版). 北京: 高等教育出版社, 2010.
[2] 韓路, 王家強, 王海珍, 宇振榮. 塔里木河上游胡楊種群結構與動態. 生態學報, 2014, 34(16): 4640- 4651.
[3] 汪洋, 冷艷芝, 蘇長江, 宋叢文, 程德華, 操英南, 張敏, 付翠林. 恩施天然紅椿種群結構及空間分布格局. 浙江農林大學學報, 2016, 33(1): 17- 25.
[4] 劉普幸. 疏勒河中下游綠洲胡楊種群結構與動態研究. 自然資源學報, 2011, 26(3): 429- 439.
[5] Armesto J J, Casassa I, Dollenz O. Age structure and dynamics of Patagonian beech forests in Torres del Paine National Park, Chile. Vegetatio, 1992, 98(1): 13- 22.
[6] Fuchs M A, Krannitz P G, Harestad A S. Factors affecting emergence and first-year survival of seedlings of Garry oaks (Quercusgarryana) in British Columbia, Canada. Forest Ecology and Management, 2000, 137(1/3): 209- 219.
[7] 廣東省植物研究所. 海南植物志(第四卷). 北京: 科學出版社, 1977: 155.
[8] Chen X Q, Turland N J.DracaenaVandelli ex Linnaeus // Wu Z Y, Raven P H. Flora of China, Vol. 24. Beijing: Science Press and St. Louis: Missouri Botanical Garden Press, 2000: 215- 217.
[9] 中國科學院植物研究所. 中國數字植物標本館. http://www.nhpe.org/cvh/.[2017-08- 12]
[10] 國家環境保護局, 中國科學院植物研究所. 中國珍稀瀕危保護植物名錄(第一冊). 北京: 科學出版社, 1987: 1- 38.
[11] 羅赫, 黃世滿. 海南龍血古樹優勢群叢的調查. 熱帶林業, 2007, 35(4): 50- 51, 41- 41.
[12] 鄭道君, 云勇, 吳宇佳, 李海文, 張治禮. 海南龍血樹野生資源分布及其與水熱關系的分析. 熱帶亞熱帶植物學報, 2012, 20(4): 326- 332.
[13] 鄭道君, 謝良商, 王盈, 張治禮, 張文. 中國血竭基源植物的研究與利用. 中國野生植物資源, 2009, 28(6): 15- 20.
[14] 梅文莉, 戴好富, 吳嬌, 莊令, 洪葵. 海南龍血樹抗腫瘤新用途研究. 中藥材, 2005, 28(10): 871- 873.
[15] 楊先會, 鄧世明, 范麗霞. 海南龍血樹植物資源的開發利用. 海南大學學報: 自然科學版, 2004, 22(3): 270- 272.
[16] 楊本鵬, 張樹珍, 蔡文偉, 宋啟示, 王興紅, 王東, 楊學, 顧麗紅, 馮翠蓮, 王俊剛, 羅遵喜. 海南龍血樹組織培養過程中血竭形成的誘導. 熱帶作物學報, 2009, 30(2): 181- 185.
[17] Luo Y, Dai H F, Wang H, Mei W L. Chemical constituents from dragon′s blood ofDracaenacambodiana. Chinese Journal of Natural Medicines, 2011, 9(2): 112- 114.
[18] 羅冠勇, 鐘云芳, 宋希強, 戴好富, 楊冬華. 海南龍血樹基于愈傷組織誘導途徑的組培快繁技術體系優化. 熱帶作物學報, 2012, 33(10): 1824- 1828.
[19] Zheng D J, Xie L S, Zhu J H, Zhang Z L. Low genetic diversity and local adaptive divergence ofDracaenacambodiana(Liliaceae) populations associated with historical population bottlenecks and natural selection: an endangered long-lived tree endemic to Hainan Island, China. Plant Biology, 2012, 14(5): 828- 838.
[20] 鄭道君, 李海文, 云勇, 張浪, 張治禮. 海南龍血樹種群生境及自然更新能力調查. 熱帶亞熱帶植物學報, 2010, 18(6): 627- 632.
[21] 鄭道君, 吳宇佳, 云勇, 姜殿強, 陳宣, 張治禮. 瀕危植物海南龍血樹種子萌發及其環境適應性分析. 熱帶亞熱帶植物學報, 2016, 24(1): 71- 79.
[22] 周文嵩, 馮丹丹, 李東海, 陶楚, 萬春紅, 邢莎莎, 楊小波. 海南昌江縣瀕危藥用植物的種群動態. 熱帶生物學報, 2014, 5(4): 392- 399.
[23] 李偉杰, 肖艷, 楊海建, 陳沂章, 曾琳, 何明軍, 鄭希龍. 海南龍血樹種群的年齡結構及分布格局. 氨基酸和生物資源, 2017, 39(1): 17- 24.
[24] 張維, 李海燕, 賴曉輝, 楊允菲. 新疆天山峽谷不同坡向野核桃種群分布格局. 應用生態學報, 2016, 27(10): 3105- 3113.
[25] 李肇晨, 陳永富, 洪小江, 韓文濤, 李小成. 海南霸王嶺陸均松種群年齡結構與點格局分布特征. 生態學雜志, 2015, 34(6): 1507- 1515.
[26] 陳曉德. 植物種群與群落結構動態量化分析方法研究. 生態學報, 1998, 18(2): 214- 217.
[27] 江洪. 云杉種群生態學. 北京: 中國林業出版社, 1992.
[28] Silvertown J W. 植物種群生態學導論. 祝寧, 王義弘, 陳文斌, 譯. 哈爾濱: 東北林業大學出版社, 1987.
[29] 吳承禎, 洪偉. 杉木數量經營學引論. 北京: 中國林業出版社, 2000.
[30] 畢曉麗, 洪偉, 吳承禎, 閆淑君, 藍斌. 黃山松種群統計分析. 林業科學, 2002, 38(1): 61- 67.
[31] 蘇建榮, 劉萬德, 繆迎春, 李帥鋒, 郎學東. 云南紅豆杉種群生物學(第三十一版). 北京: 科學出版社, 2016.
[32] 李曉笑, 王清春, 崔國發, 楊傳東. 瀕危植物梵凈山冷杉野生種群結構及動態特征. 西北植物學報, 2011, 31(7): 1479- 1486.
[33] Svensson J S, Jeglum J K. Structure and dynamics of an undisturbed old-growth Norway spruce forest on the rising Bothnian coastline. Forest Ecology and Management, 2001, 151(1/3): 67- 79.
[34] 王金山, 哈力克·玉米提, Cyffka B, 吾斯曼·庫爾班, 彭樹宏. 塔里木河下游胡楊林胸徑結構及林木分布特征. 植物學報, 2008, 25(6): 728- 733.
[35] 解婷婷, 蘇培璽, 周紫鵑, 張海娜, 李善家. 荒漠綠洲過渡帶沙拐棗種群結構及動態特征. 生態學報, 2014, 34(15): 4272- 4279.
[36] 李曉波, 王克, 范猛, 李憲高. 最小存活種群的確定與生物多樣性保護. 東北師大學報: 自然科學版, 2001, 33(3): 86- 90.
[37] 吳成貢, 蔣昌順. 瀕危植物保護生物學技術研究進展. 華南熱帶農業大學學報, 2006, 12(3): 49- 51.
[38] 國家林業局野生動植物保護與自然保護區管理司, 云南省林業廳, 中國科學院昆明植物研究所. 云南省極小種群野生植物保護實踐與探索. 昆明: 云南科技出版社, 2013: 1- 100.
[39] 許再富, 高江云, 李保貴, 周惠芳. 國家重點保護植物“遷地”與“近地”保護有效性的比較研究//中國植物園(第十五期). 西安: 中國植物學會, 2011: 6- 15.
[40] 胡世俊. 生境片斷化與隔離對瀕危植物縉云衛矛(EuonymuschloranthoidesYang)種群特征的影響[D]. 重慶: 西南大學, 2007
