新疆石河子地區棉花黃萎病菌分離鑒定及其致病力分析
2018-06-29 06:47:02張夢恬裴娟李國趙輝陳建權祝建波王愛英
生物技術通報 2018年6期
張夢恬 裴娟 李國 趙輝 陳建權 祝建波 王愛英
(石河子大學生命科學學院 農業生物技術重點實驗室 石河子 832003)
棉花是世界上重要的纖維作物,由于新疆獨特的自然氣候使其成為國內面積最大的產棉區。棉花黃萎病是全球范圍內的棉花病害之一,極大影響棉花質量及產量。棉花黃萎病是由輪枝菌屬(Verticillium)的病原菌引起,目前發現有6個種是植物病原菌,分別為大麗輪枝菌(V . dahliae Klebahn)、變黑輪枝菌(V.nigrescens Pethybridge)、三體輪枝菌(V.tricorpus Isaac)、黑白輪枝菌(Verticillium albo-atrum Reinke and Berthold)、 云狀輪枝菌(V.nubilum Pethybridge)及V.theobromae(Turconi)[1-4]。其中造成棉花黃萎病的主要病原菌為大麗輪枝菌和黑白輪枝菌。由于植棉地區生態環境發生變化,造成病菌表現出不同的致病類型,從而產生許多致病力強的菌株類型,但是變異形成的菌株類型仍然不是太清楚。姚耀文等[5]通過致病力測定將我國棉花黃萎病分為3個生理型,分別為:生理型Ⅰ致病力極強,生理型Ⅱ致病力極弱,以及致病力中等介于前兩種生理型之間的生理型Ⅲ。段維軍等[6]證實新疆存在少量落葉型黃萎菌。韓宏偉等[7]通過分子手段發現新疆北部(北疆)落葉型黃萎菌比例占到39%。李彩紅等[8]發現氣候條件的改變會影響黃萎病菌產生遺傳變異及致病類型差異。劉燕霞等[9]利用ISSR標記技術對不同地區大麗輪枝菌遺傳分化及致病力進行分析。黃萎病菌菌株間的親緣關系可以利用分子進化樹及營養親和性技術來分析。Puhalla等[10]首次利用硝酸鹽營養缺陷性突變體(nit)技術研究黃萎病菌營養親和性。本研究通過現代分子技術檢測和菌株培養表型觀察相結合的方法研究新疆石河子地區棉花黃萎病菌的致病類型及菌株的親緣關系,從而明確石河子地區棉花黃萎病菌的變異和致病力分化情況,以期為該地區篩選黃萎病的耐病或抗病棉花品種提供科學依據。
1 材料與方法
1.1 材料
1.1.1 菌株與試劑 棉花病株采自新疆石河子棉花種植區,其中SHZ-1至SHZ-9采自143團石南農場,SHZ-10至SHZ-16采自145團清泉集七連,SHZ-17至SHZ-24采自大學試驗場二連。pMD19-T克隆載體、PCR反應液購自TaKaRa公司,瓊脂糖購自Biowest公司,瓊脂糖凝膠回收試劑盒購自天根生物公司,引物合成由北京華大基因有限公司完成。
1.1.2 培養基 PDA培養基Czapek液體培養基及LB固體培養基[11]用于真菌及細菌培養;BM培養基、MM培養基、KPS培養基、MO2培養基及MH培養基[12-13]均用于營養親和性研究。
1.2 方法
1.2.1 菌株分離純化 通過岳永亮等[14]的方法對本研究中感病棉株進行組織分離。分離純化后菌株低溫保存。
1.2.2 分子生物學檢測 通過優化的CTAB法[15]提取棉花黃萎菌DNA。以其為模板,通過真菌ITS通用引物[16]ITS1/ITS4、黃萎菌特異引物 DB19/DB22、黃萎菌致病類型特異引物[17]D-1/D-2、ND-1/ND-2分別對分離菌株進行PCR擴增(表1)。PCR擴增反應體系為20 μL,Taq聚合酶0.5 μL、Taq酶緩沖液 2.5 μL、dNTP 2.0 μL、引物 1.0 μL、模板 1.0 μL。PCR 反應程序:95℃,10 min;95℃ 45 s、56℃ 30 s,72℃ 30 s,30個循環;72℃,10 min。真菌ITS通用引物擴增后獲得的陽性克隆,連接至pMD19-T載體,測序連接成功后的質粒,比對測序結果,下載比對后同源性較高的ITS序列,利用MEGA 5.0采用Neighbour-Joining Method構建病原菌株分子進化樹。

表1 黃萎病菌PCR引物
1.2.3 菌株營養親和性鑒定 通過景嵐等[18]的方法對本研究中分離得到的24株棉花黃萎病菌進行營養親和性鑒定分析
1.2.4 菌株溫室致病力鑒定 采用無底營養缽定量接菌液法[19-20],接菌孢子濃度為 1.0×107cef/mL。供試棉株為新陸早 33號、新陸早7號、中棉35號。將單孢純化后的菌株利用Czapek液體培養基,26℃、180 r/min避光振蕩培養一周,四層紗布過濾菌絲,接種于長至3片真葉的棉苗,接種15 d后,根據感病棉苗的病情指數進行黃萎菌致病力分析。
黃萎病病級調查[19-20]:0級:健株,無病狀;1級:子葉出現病狀;2級:1片真葉嚴重發病;3級:2片真葉表現病狀;4級:植株生長點死亡或全株枯死;病情指數計算公式如下:

致病力類型的劃分:強致病力為I型:平均病指30.0以上;中致病型III型:平均病指20.1-30.0;弱致病型II型:平均病指20.0以下。
2 結果
2.1 菌落形態及顯微形態鑒定
本研究分離的24株菌株菌落形態,與孔德真[21]等描述相似,菌落表面為白色絨毛狀,周圍有黑色微菌核產生,菌落形態以圓形居多,少部分為橢圓形,進行顯微觀察,發現其能夠產生與大麗輪枝菌相似的含有2-4個分枝的輪狀分生孢子梗(圖1-A);還能夠產生與大麗輪枝菌相似的黑色微菌核結構(圖1-B),在微菌核旁側有少量無色透明孢子(圖1-C)。大部分分離菌株菌落都有同心輪紋(圖2-A),部分菌落微菌核呈現出由內向外放射狀(圖2-B),綜上所述,初步判定分離菌株為棉花黃萎菌。依據菌落不同形態將供試菌株分為3種。菌核型(圖2-C):培養基中央菌核較多,邊緣菌核較少,菌絲菌核周圍形成白色圓圈,氣生菌絲不發達,無凸起。絲核型(圖2-D):產生較多菌核,呈現發散狀態,邊緣整齊,培養基中央有較多氣生菌絲,有灰白色菌絲凸起。菌絲型(圖2-E):不產生菌核,菌絲密集且邊緣整齊,無凸起,氣生菌絲不發達。
2.2 供試菌株分子檢測
供試菌株ITS區段通過真菌特異引物 ITS1 和ITS4 進行聚合酶鏈式反應擴增得到約 500 bp的目的條帶(圖3-A)。測序后,在GenBank 數據庫中通過BLAST比對,發現這些序列和已知ITS 序列具有高度同源性,從分子水平鑒定證明供試菌株為大麗輪枝菌。利用PCR手段通過黃萎菌特異性引物DB19/DB22對純化的菌株進行擴增得到 539 bp 的特異性條帶(圖3-B),從而進一步確定分離得到的菌株為大麗輪枝菌。對供試的棉花黃萎病菌采用落葉型(D-1/D-2)和非落葉型特異引物(ND-1/ND-2)進行聚合酶鏈式反應,落葉性引物擴增可得到約500 bp的目的條帶,非落葉型引物擴增可得到約1 500 bp的目的條帶(圖3-C)。確定分離得到的黃萎菌11株為落葉型,13株為非落葉型。

圖1 分離菌株的分生孢子、孢子梗及微菌核的顯微形態
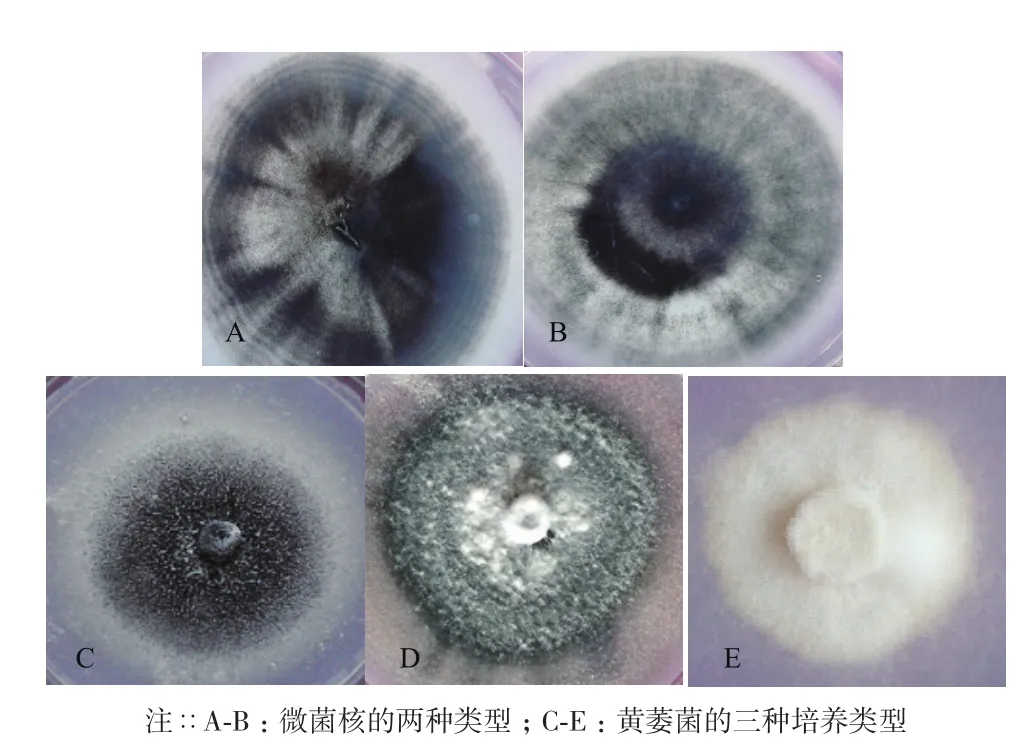
圖2 分離菌株的微菌核形態及菌落培養類型
2.3 序列比對和系統進化樹構建
將分離的24株菌株ITS序列與GenBank中同源性較高的輪枝菌 ITS 序列進行多序列比對并構建系統進化樹(圖4)。這些ITS序列信息如下:KC156638.1、U82452.1、KC156640.1、KJ696553.1、KC156635.1、KM013462.1 分別來自于中國北京、美國麥迪遜、中國河南、北京、河南及重慶;分離的24株黃萎菌中SHZ-4、SHZ-5、SHZ-6、SHZ-8、SHZ-9、SHZ-11在同一進化支,與來自北京的KJ696553.1相近;SHZ-24與來自河南KCl156635.1在同一進化支比較相近;SHZ-17、SHZ-23在同一進化支上,與進化上來自美國的U82452.1和來自中國河南的KCl156640.1比較相近。
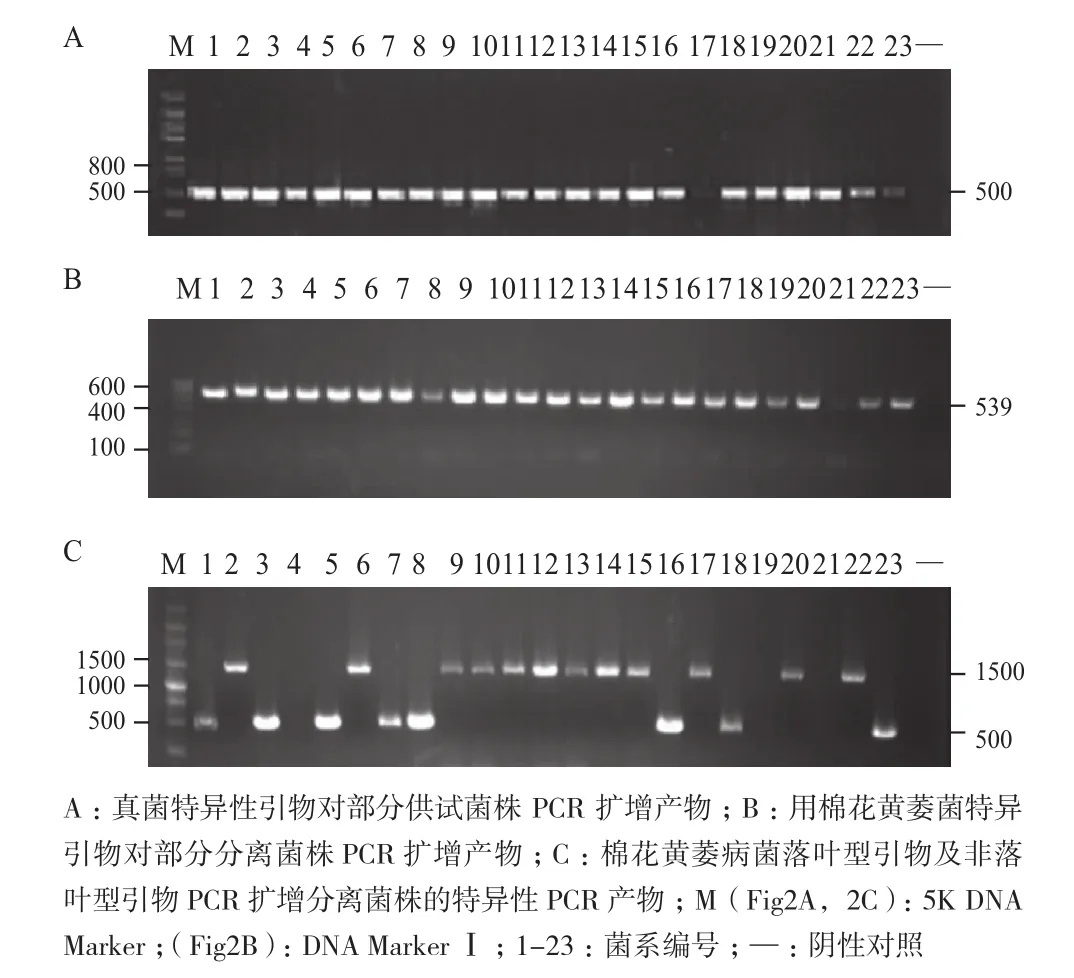
圖3 分離菌株ITS及特異性的PCR擴增產物電泳圖
2.4 分離菌株的營養親和鑒定
通過誘導培養基誘導培養15 d后,分離菌株中有10株菌株可產生突變體菌落,菌落邊緣呈現擴散性生長扇形突變體菌落。而其中14株不能產生突變體。24株菌株共獲得53個突變體,nit1、nitM、nit3的數值分別為39、10、4,分別占總突變體的73.58%、18.87%及7.54%。nit1突變體數量最多,最易獲得;nitM次之;nit3突變體最難獲得。對分離得到的24株黃萎菌的nit1和nitM突變體進行全組合配對,研究菌株之間的親和性。結果(表2)表明:供試菌株可分為3個不同的營養親和群,其中SHZ-4、SHZ-5、SHZ-6、SHZ-8、SHZ-9、SHZ-11 屬于一個親和群,命名為VCG1。SHZ-2、SHZ-18屬于一個親和群,命名為VCG2。SHZ-13、SHZ-21屬于一個親和群,命名為VCG3。
2.5 黃萎病菌致病力測定
利用無底營養缽定量接菌液法測定24株黃萎菌在3個不同的鑒別寄主上的致病力,結果顯示(表3)24株菌株表現出明顯的致病力差異。根據致病力不同將24株菌株劃分為強、中、弱3種不同類型。強致病力占75%,中等致病力占8.3%,弱致病力占16.7%。其中SHZ-9菌株的致病力較強,SHZ-4號菌株的致病力較弱。

圖4 分離的24株石河子棉花棉花黃萎病菌ITS序列構建的N-J進化樹
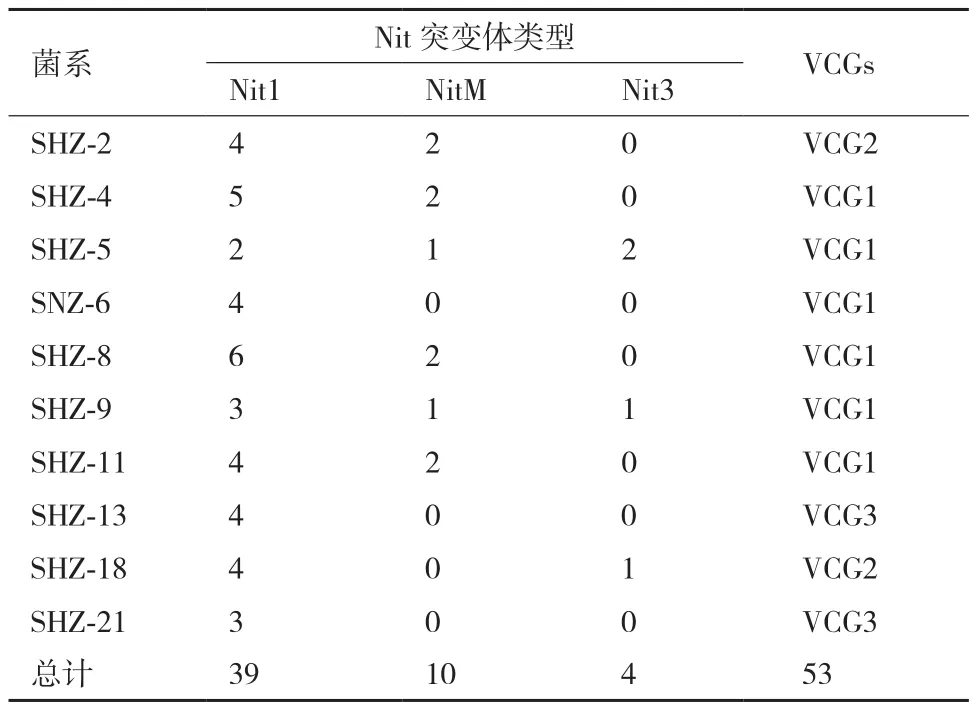
表2 分離菌株得到Nit突變體的數目及其營養親和群

表3 24個不同供試病株對3個棉花品種的致病性差異鑒定
3 討論
棉花黃萎病菌本身變異情況及致病力分化研究可為培育優良抗病品種及有效的病害預防提供理論基礎。棉花黃萎病多是由大麗輪枝菌造成的土傳病害,特點是分布廣、變異性強、造成危害大。李國英等[22]研究表明石河子地區于2001-2008年間,黃萎病發病面積成倍增加,發病率高于30%的發病面積更是增加8倍以上。韓宏偉[7]等研究表明2005-2010年間新疆北部棉花黃萎菌由以中、弱致病力為主轉變為以強致病力為主。本研究從石河子不同地區自然病圃采集感病植株,經組織分離、分子鑒定后獲得24株大麗輪枝菌,其中強、中致病力類型菌株有20株,占到83.3%,說明石河子棉區黃萎菌以強、中致病力為主,同時本研究中強致病力菌株占到75%,這與韓宏偉等研究結果中石河子地區強致病力菌系占51.7%相比高出了23.3%,由此說明,石河子棉區黃萎病情明顯加重、枯死型癥狀增多的主要原因是石河子地區黃萎菌群體致病力類型發生較大變化,致病力強菌系明顯增加。
2004年,研究表明新疆僅有少量落葉型黃萎菌存在;2008年,新疆北部落葉型黃萎菌達到39%[7];2009年新疆石河子地區落葉型黃萎菌已達到37.9%[23]。本研究分離所獲24株菌株中,落葉型黃萎菌11株,占到45.8%,這與2009年相比高出了6.8%。同時11株落葉型黃萎菌中強致病力類型9株,占81.8%,由此說明,石河子棉區在生長的中后期出現大面積落葉致光桿的主要原因是石河子地區黃萎菌群體出現較多致病性強的落葉型菌系。同時SHZ-9是24株菌株中致病力最強的菌株,但是它是非落葉型菌系,這與王國寧等[24]發現有些非落葉型黃萎菌致病力強于落葉型黃萎菌相一致。
本研究發現SHZ-9與SHZ-4在分子進化樹上親緣關系較近,且同屬于VCG1,分子檢測后發現兩者均為非落葉型大麗輪枝菌,但侵染相同品種棉花后表現出的致病力有顯著差異,SHZ-9是24株菌株中致病力最強的菌株,平均病情指數為50,而SHZ-4是致病力較弱的菌株,平均病情指數僅為17.6,這表明同為非落葉型菌系接種相同品種能表現出完全不同的致病力,與朱荷琴等[25]研究結果相似,兩者致病力分化明顯的原因還有待深入研究。
4 結論
本研究分離獲得24株石河子地區棉花黃萎病致病菌株均為大麗輪枝菌,11株落葉型黃萎菌,13株非落葉型黃萎菌。分子進化樹顯示24株菌在系統進化方式上屬于2個不同的分支。致病力結果顯示24株菌株強致病力占到75%,中等致病力占到8.3%,弱致病力占到16.7%,SHZ-9致病力最強,SHZ-4致病力較弱,菌株存在明顯致病力分化現象。
[1]Barbara DJ, Clewes E. Plant pathogenic Verticillium species:how many of them are there?[J]. Mol Plant Pathol, 2003, 4:297-305.
[2]Qin QM, et al. Phylogenetic analyses of phytopathogenic isolates of Verticillium spp.[J]. Phytopathol, 2006, 6 :582-592.
[3]Klosterman SJ, Atallah ZK, Vallad GE, et al. Diversity,pathogenicity, and management of verticillium species[J]. Annual Review of Phytopathol, 2009, 47(1):39-62.
[4]White TJ, Bruns T, Lee S, et al. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics[M]// Innis MA,Gelfand DH, Sninsky JJ, et al. PCR-Protocols:A guide to methods and applications. New York:Academic Press, 199:315-322.
[5]姚耀文, 傅翠真, 王文錄, 等. 棉花黃萎病菌生理型鑒定的初步研究[J]. 植物保護學報, 1982, 9(3):145-148.
[6]段維軍, 李國英, 張莉, 等. 新疆棉花黃萎病菌致病性分化監測研究[J]. 新疆農業科學, 2004, 41(5):324-328.
[7]韓宏偉, 劉培源, 高峰, 等. 新疆北部棉區黃萎病菌種群致病性分化及變異[J]. 植物保護學報, 2011, 38(2):121-126.
[8]李彩紅, 王永波, 趙瑞元, 等. 湖南常德地區棉花黃萎病菌培養特性及致病力分化研究[J]. 中國棉花, 2015(7):18-21.
[9]劉燕霞, 侯麗娟, 等. 棉花黃萎病菌ISSR反應體系優化及其遺傳多樣性分析[J]. 植物保護學報, 2010(5):425-430.
[10]Puhalla JE, Hummel M. Vegetative compatibility groups within Verticillium dahliae[J]. Phytopathol, 1983, 9 :1305-1308.
[11]沈萍, 陳向東. 微生物學實驗[M]. 第4版. 北京:高等教育出版社, 2007.
[12]Brooker NL, Leslie JF, Dickman MB. Nitrate non-utilizing mutants of Colletotrichum and their use in studies of vegetative compatibility and genetic relatedness[J]. Phytopathol, 1991, 6 :672-677.
[13]Emi K, Shuji T, Jun-Ichi S, et al. Agrobacterium tumefaciensmediated transformation of Aspergillus aculeatus for insertional mutagenesis[J]. Amb Express, 2011, 1 :46.
[14]岳永亮, 任毓忠, 張莉, 等. 新疆榆樹黃萎病病原菌鑒定[J].植物保護, 2016, 42(2):251-253.
[15]伏榮桃, 王劍, 陳誠, 等. 稻曲病菌基因組DNA提取方法比較與小文庫構建[J]. 生物技術通報, 2018, 34(3):102-106.
[16]姜雨萌, 牛永春, 鄧暉. rDNA ITS序列在ACCC真菌鑒定中的應用[J]. 微生物學通報, 2016, 43(5):942-947.
[17]Pérezartés E, et al. Differentiation of cotton-defoliating and nondefoliating pathotypes of Verticillium dahliae by RAPD and specific PCR analyses[J]. Eur J Plant Pathol, 2000, 6 :507-517.
[18]景嵐, 裴旭, 楊柳青, 等. 向日葵黃萎病菌營養親和性及致病力分化[J]. 中國油料作物學報, 2014, 36(1):106-112.
[19]任杰, 張貴, 曹雄, 等. 向日葵黃萎病菌不同培養型產毒能力和致病力分化[J]. 中國油料作物學報, 2014(3):393-397.
[20]石磊巖, 王波, 文學. 我國棉花黃萎病菌類型分化及培養特性研究[J]. 植物保護學報, 1993(3):247-252.
[21]孔德真, 黎玉順, 裴娟, 等. 新疆烏蘇地區棉花黃萎病菌分離鑒定和致病力分析[J]. 中國農學通報, 2015(20):61-66.
[22]李國英, 張新全, 宋玉萍, 等. 北疆棉區棉花黃萎病發生趨勢、抗性研究[J]. 新疆農業科學, 2015, 52(1):185-190.
[23]韓宏偉, 等. 石河子地區棉花黃萎病菌培養特性和致病力分化研究[J]. 新疆農業科學, 2011, 48(3):522-527.
[24]王國寧, 趙貴元, 岳曉偉, 等. 河北省棉花黃萎病菌致病性與ISSR遺傳分化[J]. 棉花學報, 2012, 24(4):348-357.
[25]朱荷琴, 李志芳, 馮自力, 等. 我國棉花黃萎病研究十年回顧及展望[J]. 棉花學報, 2017, 29(S1):37-50.
