Numb基因不同亞型對乳腺癌細胞MCF-7增殖和遷移能力的影響
2018-07-05 09:29:26王保壘李華順2朱晨光
上海大學學報(自然科學版) 2018年3期
蘇 杭, 王保壘, 李華順2,, 朱晨光
(1.上海大學生命科學學院,上海200444;2.中國科學院上海高等研究院干細胞與納米醫學研究中心,上海201210;3.同濟大學附屬東方醫院轉化醫學高等研究院,上海200085)
Numb作為細胞不對稱性分裂的決定分子,最早被發現參與神經系統的不對稱分裂,近年來被發現與腫瘤的惡性程度密切相關[1].Numb的缺陷會導致腫瘤的惡性程度增大,Numb在抑制上皮間質轉化(epithelial-mesenchymal transitions,EMT)中的作用也較受關注[2].然而Numb表達缺陷導致腫瘤惡性程度增大的機制尚不清楚.研究表明,Numb能夠阻滯抑癌蛋白p53被泛素化和降解的過程,并且與Notch,Hedgehog等重要信號通路密切相關;同時,Numb還是一種調節細胞內吞的因子[3-8].據報道Numb有6種剪切形式,但是最為廣泛關注的剪切體有4種,分別為膜聯形式(p66和p72)和胞質形式(p65和p71),其分子及細胞內定位的多樣性使研究Numb的功能機制變得更為復雜[9-12].過去的一項研究對321例乳腺癌樣本Numb進行免疫組化分析,發現Numb陽性率越低,乳腺癌的惡性程度就會越大[5].但是,很多研究僅僅通過Numb表達的總量來對腫瘤惡性程度進行分類,很少有研究涉及到Numb蛋白不同亞型在腫瘤細胞中的作用,而Numb的不同蛋白亞型之間,不論是在發育的時空表達方面,還是在功能方面,都有明顯差異.研究Numb的不同亞型在腫瘤中的表達和功能,是一項亟待解決的問題.
區分Numb不同形式的拼接分子的不同功能,對進一步了解Numb的功能及其調控機制極為必要.基于前期的研究結果,本工作研究Numb在乳腺癌細胞發生過程中的作用,并著重探討不同拼接形式的Numb亞型對乳腺癌細胞的作用及機制,嘗試以Numb的不同亞型表達高低作為腫瘤惡性程度分型的標志,為癌癥個性化治療提供指導.
1 材料與方法
1.1 材料
DMEM(dulbeceo’s modif i ed eagle medium)培養液(高糖)、胎牛血清、磷酸鹽緩沖鹽溶液(phosphate buあer saline,PBS)(Hyclone公司);胰蛋白酶-乙二胺四乙醇(ethylendiaminetetraacetic acid)(Life Technologies公司);轉染試劑Megatran1.0(Origene公司);細胞系篩選抗生素嘌呤霉素(Puromycin)(Sigma公司);放射免疫沉淀實驗(radio immunoprecipitation assay,RIPA)裂解液(強)(碧云天公司);蛋白酶抑制劑coctail(Roche公司);質粒小量提取試劑(Omega公司);質粒中量提取試劑(MN公司);基因組提取試劑(天根生物公司);轉錄激活樣效應因子核酸酶(transcription activator-like eあector nuclease,TALEN)試劑(斯丹賽公司);各種限制性內切酶BamHⅠ和PstⅠ(NEB公司);細胞增殖與活性檢測試劑CCK-8(同仁化學公司);人乳腺細胞系MCF-7細胞和人胚腎上皮細胞系293T細胞(中國科學院上海生命科學院細胞庫);兔抗人Numb多克隆抗體(Cell Signaling Technology公司);大鼠抗小鼠E-cadherin抗體(Takara公司);小鼠來源β-actin內參抗體(金斯瑞公司);山羊抗兔IgG(H&L)Antibody IRDye800CW抗體和山羊抗小鼠IgG(H&L)Antibody IRDye800CW抗體(Rockland Immunochemicals公司);Numb剪切體表達載體p65,p66,p71和p72(同濟大學附屬東方醫院王保壘老師構建).
1.2 細胞培養
乳腺癌細胞MCF-7和人胚腎上皮細胞系293T細胞均采用含10%胎牛血清高糖DMEM培養液,置于37°C、CO2體積分數為5%的培養箱中培養.
1.3 TALEN打靶載體構建及驗證
人體Numb基因有23個轉錄本,選取23個轉錄本中共同的編程序列(coding sequencce,CDS)區TCCAGAGGCCAGTCGTCCACATCAGTGGCAGACAGATGAAGAAGGCGTTCGC設計打靶位點組合.TALEN打靶載體的構建及驗證參照斯丹賽公司提供的TALEN試劑盒使用說明書.
1.4 Numb敲除的MCF-7細胞系構建
在成功構建Numb基因TALEN打靶載體后,利用細胞轉染試劑Megatran 1.0按標準方法將載體轉入MCF-7細胞,Megatran 1.0與載體比例為3μL:1μg.24 h后換成含2μg/mL Puromycin的篩選培養基,篩選培養3 d,3 d后去上清,用PBS洗3次,消化細胞并計數,利用有限稀釋法接種在96孔板中,待單克隆細胞團長起,消化細胞,提取基因組,聚合酶鏈式反應(polymerase chain reaction,PCR)擴增Numb基因片段,送測序.Numb基因PCR引物序列 (5’-3’):AGAGGTAGATGGGTAGGCAGA,CACTCCCTCAGCTTCACCAG.Numb 基因測序引物序列(5’-3’):GTTAGTATGGTAAT-CCAAAG.選取Numb基因發生突變的細胞系,經Western blot檢測Numb蛋白表達情況,選取不表達蛋白的細胞系,作為Numb-KO穩定細胞系.
1.5 Western blot實驗
Numb基因敲除和正常MCF-7細胞用胰酶消化,收集細胞,用冷的PBS洗3次,然后用蛋白裂解液RIPA(強)冰上裂解30 min,4°C 13 000 g離心15 min,取上清即為提取的總蛋白懸液;二喹啉甲酸(bicinchoninic acid,BCA)法測量蛋白質量百分比,加入SDS上樣緩沖液100°C煮5 min,冰上放置5 min,即可進行SDS-聚丙烯酰氨凝膠電泳(polyacrylamide gel electroph-oresis,PAGE)電泳;完成電泳后轉印到聚偏二氟乙烯(polyvinylidene f l uoride,PVDF)膜上,294 mA,1 h;用含5%脫脂奶粉的PBS封閉1 h;4°C一抗(1:1 000)過夜,然后用PBS洗3次,每次5 min;二抗(1:10 000)室溫孵育1 h,然后用PBS洗3次,每次5 min,利用Odyssey雙色紅外熒光成像系統成像.
1.6 Numb各亞型瞬時表達細胞系構建
將p65,p66,p71,p72和EGPF這5種質粒表達載體轉染進Numb-KO MCF-7細胞中,分別命名為p65組,p66組,p71組,p72組和增強綠色熒光蛋白(enhanced green f l uorescent protein,EGFP)組(陰性對照).使用的轉染試劑為Megaran 1.0,于轉染后24 h更換培養基,各瞬時表達細胞系用于以后的實驗.
1.7 細胞增殖曲線測定
收集細胞,加細胞懸液100μL(5 000個細胞)到96孔板(邊緣孔用無菌水或PBS填充).設置空白孔(有培養基,無細胞)和對照孔(正常MCF-7細胞),每組設定5個復孔.置37°C,5%CO2孵育過夜,倒置于顯微鏡下觀察.每孔加入10μL CCK-8溶液,37°C孵育1 h.測定450 nm各孔的吸光值.將各測試孔的光密度值(optical density,OD)值減去空白孔OD值,即為相對吸光度.
1.8 細胞劃痕實驗測定遷移能力
將Numb-KO MCF-7組、MCF-7組細胞接種于6孔板中(5×104個細胞/孔),待細胞呈現單層緊密貼壁生長時用黃色移液器吸頭劃痕,用PBS洗去懸浮細胞,用含2%胎牛血清的高糖DMEM培養基繼續培養細胞.分別于劃痕后0,24,48 h在光學顯微鏡下觀察并拍照,并應用Image-Pro Plus 6.0軟件測量劃痕間的距離,計算細胞遷移率.
1.9 PI染色檢測細胞周期
計數細胞,在流式管中加入1×106個細胞,用PBS洗滌1次(300 g離心5 min).用多聚甲醛固定細胞:去上清,加入500μL冷PBS,輕柔混勻.加入500μL的4°C的2%多聚甲醛,再次混勻.4°C孵育1 h.乙醇破膜:300 g離心5 min,去上清,再次用3 mL冷PBS洗滌1次.逐滴加入1 mL 70%的乙醇(?20°C),邊加邊振蕩混勻,4°C孵育過夜.PI染色:300 g離心5 min,去上清,加入RNAse工作液1 mL,室溫放置30 min,300 g離心,去上清加入1 mL碘化丙啶(propidium iodide,PI)工作液,37°C避光孵育30 min,用40或70μm的濾網過濾掉團塊.上機分析.
1.10 流式細胞術檢測細胞表面蛋白
將單細胞懸液加入離心管中,離心,300 g,5 min,棄上清液.以冷PBS 1 mL離心洗滌,棄上清液.用PBS稀釋的第一抗體200μL,對照管中加入對應于一抗的動物IgG,輕輕吹打混勻,4°C或置冰上孵育1.5~2.0 h.離心,棄上清.PBS 1 mL離心洗滌2次,以去除多余的未結合的特異性抗體.PBS適當稀釋的熒光素標記的第二抗體200μL,吹打混勻,4°C或置冰上孵育30 min,避光.PBS 1 mL離心洗滌2次.將細胞重新懸浮于0.5 mL的PBS中,混勻,置流式管中,避光,4°C冰箱保存,4 h內上機檢測.
1.11 統計學方法
本工作中的各項實驗均重復3次.采用SPSS 18.0軟件進行數據分析,用Student’s t-test分析2組間的差異,以p<0.05表示差異具有統計學意義.
2 結果
2.1 利用TALEN技術建立Numb基因敲除體外模型
構建的TALEN敲除載體轉染MCF-7細胞后,經Puromycin抗性篩選,單克隆挑取,PCR擴增Numb靶點DNA序列,測序驗證,選取Numb基因發生突變的單克隆細胞擴大培養.測序結果顯示,Numb-KO MCF-7細胞Numb基因發生缺失7個堿基的突變(見圖1).經Western blot檢測,檢測不到Numb蛋白的表達,故Numb敲除細胞模型成功建立(見圖2).
2.2 Numb基因敲除促進MCF-7細胞增殖
CCK-8檢測細胞增殖結果顯示,從培養第3天起,Numb-KO MCF-7的細胞增殖速度明顯高于對照組正常MCF-7細胞,說明Numb基因敲除能夠明顯促進MCF-7的增殖,提示Numb對乳腺癌發生發展具有抑制作用(見圖3).PI染色檢測細胞周期結果顯示,在敲除Numb基因后,G0G1期細胞比例降低了9.28%,G2M期細胞比例增加了4.36%,S期細胞比例增加了4.92%,說明Numb可以抑制MCF7細胞由G0G1期往G2M期和S期轉化,對MCF-7細胞的增殖起到抑制作用(見表1).
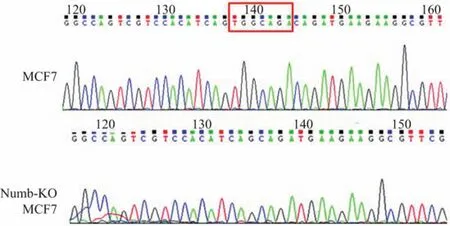
圖1 Numb-KO MCF-7細胞Numb基因測序結果(紅框內:缺失的堿基)Fig.1 DNA sequencing of Numb in Numb-KO MCF-7 cells(Red box:deleted bases)
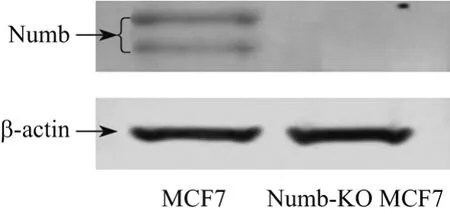
圖2 MCF-7細胞和Numb-KO MCF-7細胞Numb表達Fig.2 Expression of Numb protein in MCF-7 and Numb-KO MCF-7 cells
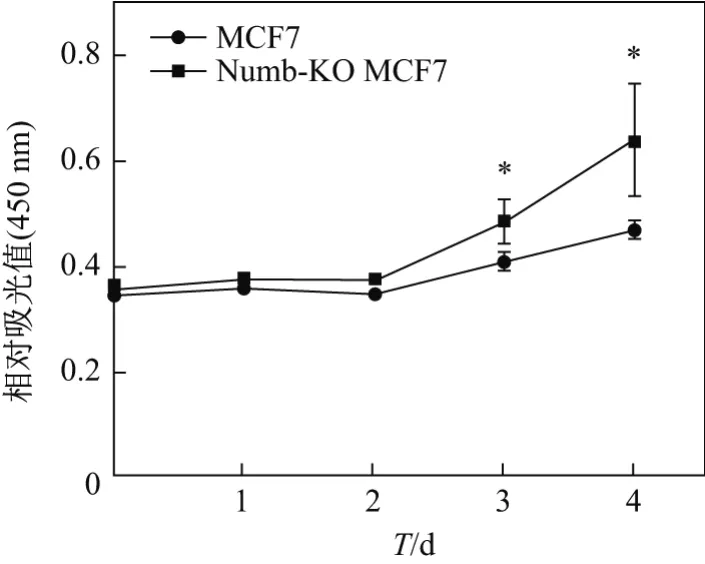
圖3 CCK-8檢測MCF-7細胞增殖Fig.3 MCF-7 cell proliferation assessed by CCK-8

表1 PI染色及流式細胞術檢測MCF-7和Numb-KO MCF-7的細胞周期Table 1 Detection for cell cycle of MCF-7 and Numb-KO MCF-7 cells by staining with PI and f l ow cytometry %
2.3 Numb基因敲除促進MCF-7細胞遷移
采用細胞劃痕實驗檢測正常MCF-7細胞和Numb敲除后MCF-7細胞遷移能力的結果顯示,在細胞劃痕48 h后,Numb-KO MCF-7細胞遷移率比陰性對照組正常MCF-7細胞遷移率高出22%,提示Numb對MCF-7細胞的遷移能力有抑制作用(見圖4).流式細胞術檢測上皮細胞表面鈣粘蛋白(E-cadherin)結果顯示,Numb敲除后的MCF-7細胞細胞表面E-cadherin的表達量(平均熒光強度(median f l uoresence intensity,MFI))降低了20.28%,說明Numb敲除后MCF-7細胞間黏附能力減弱,細胞遷移能力增強(見圖5).
2.4 Numb基因各亞型轉染效率觀察
Numb基因各亞型轉染Numb-KO MCF-7細胞24 h后,在熒光顯微鏡下觀察可見載體質粒上的綠色熒光蛋白表達,細胞發出綠色熒光,轉染效率大于90%,說明載體基因得到充分表達(見圖6).圖6中,第1行圖為在相差顯微鏡明場下觀察,第2行圖為暗場觀察綠色熒光.
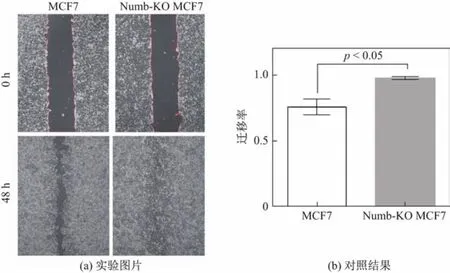
圖4 劃痕實驗檢測細胞遷移能力Fig.4 Detection for the abilities of migration of MCF-7 cells by wound scratch assay
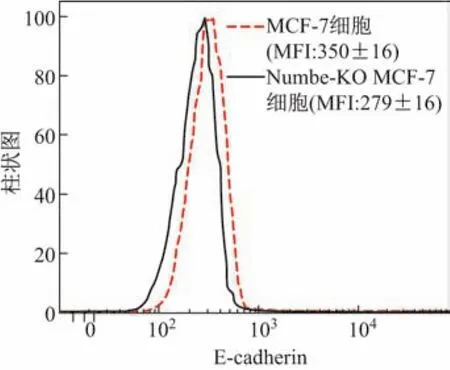
圖5 流式細胞術檢測E-cadherin表達水平Fig.5 E-cadherin expression level detected by f l ow cytometry
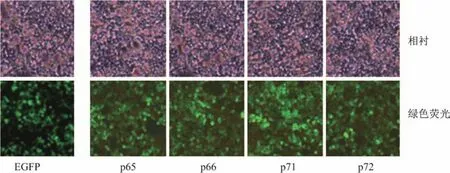
圖6 Numb-KO MCF-7轉染后熒光顯微鏡觀察Fig.6 Fluorescent microscope observation of Numb-KO MCF-7 cells
2.5 Numb基因的4個亞型對MCF-7細胞增殖的影響
采用CCK-8細胞增殖實驗結果繪制細胞增殖曲線(見圖7(a)),發現在培養的第3天時,與陰性對照組(EGFP)相比,Numb基因的亞型P72轉染組細胞增殖能力明顯減弱.當培養第3天時各組細胞增殖情況可由圖7(b)直觀地看出.相對吸光值p72組比EGFP組降低16%,提示p72可以抑制Numb-KO MCF-7細胞的增殖.其他3種亞型p65,p66,p71對Numb-KO MCF-7細胞增殖無明顯抑制作用(見圖7).通過PI檢測細胞周期實驗結果發現,對Numb-KO MCF-7細胞增殖抑制最明顯的仍為p72,p72轉染后可將G0G1期細胞比例提高17.75%,S期細胞比例降低18.2%,對G2M期細胞比例沒有影響,說明p72可以抑制細胞由G0G1往S期的轉變,從而抑制細胞增殖(見表2).
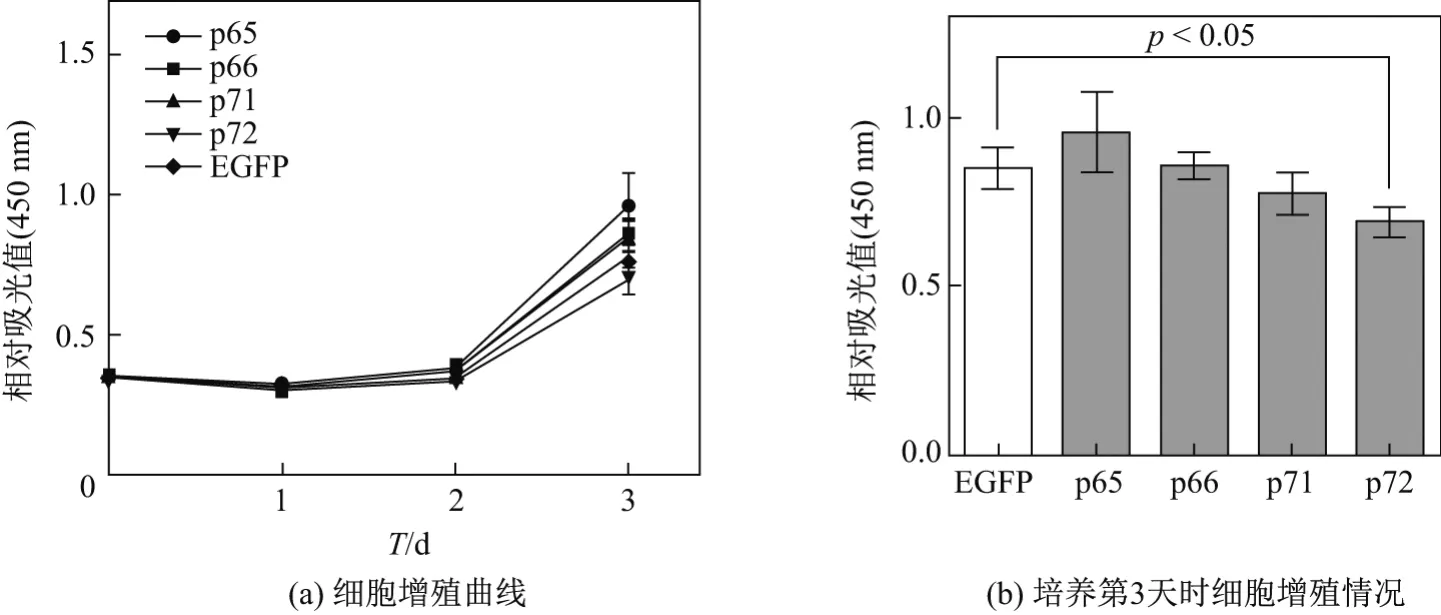
圖7 CCK-8檢測個亞型轉染后Numb-KO MCF-7細胞增殖能力Fig.7 Numb-KO MCF-7 cell proliferation assessed by CCK-8 after transfection with Numb isoforms
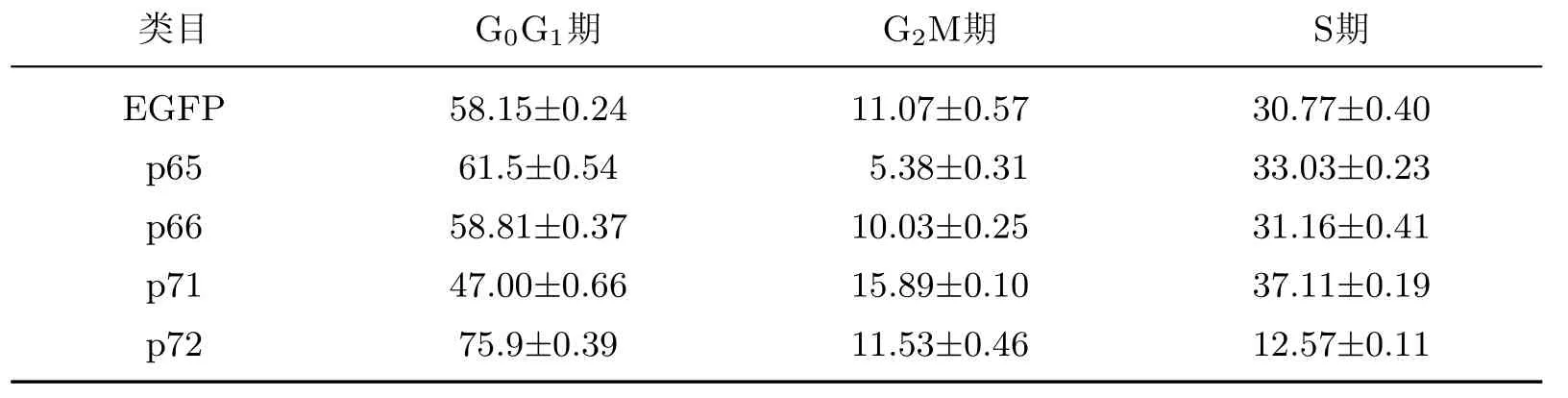
表2 PI染色及流式細胞術檢測各亞型轉染后Numb-KO MCF-7的細胞周期Table 2 Detection for cell cycle of Numb-KO MCF-7 cells after p65,p66,p71 and p72 expression vector transfecting by staining with PI and f l ow cytometry %
2.6 Numb基因的4個亞型對MCF-7細胞侵襲能力的影響
細胞劃痕后于含2%FBS的高糖DMEM培養基中培養24 h,結果顯示各亞型轉染組遷移率與陰性對照EGFP組相比,遷移比率降低程度分別為p65組8.29%,p66組7.95%,p71組8.95%和p72組8.49%,這說明4種亞型均對Numb-KO MCF-7細胞遷移能力有抑制作用,但是抑制能力沒有差異(見圖8).用流式細胞術檢測各亞型轉染后Numb-KO MCF-7細胞表面E-cadherin的表達情況后發現,4種亞型轉染組與EGFP(陰性對照)組相比,E-cadherin表達量(MFI)均有所提高,提高程度分別為p65組23.65%,p66組39.78%,p71組24.73%和p72組44.80%,說明4種亞型均對Numb-KO MCF7細胞的遷移能力起到抑制作用(見表3).
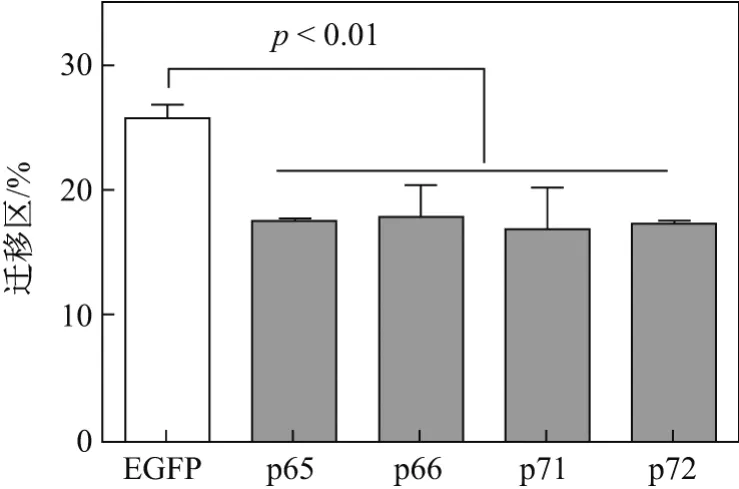
圖8 劃痕實驗檢測Numb各亞型對Numb-KO MCF-7細胞遷移能力的影響Fig.8 Dection for the abilities of migration of Numb-KO MCF-7 cells by wound scratch assay

表3 流式細胞術檢測各亞型轉染后Numb-KO MCF-7細胞表面E-cadherin表達水平Table 3 E-cadherin expression levels of Numb-KO MCF-7 cells after p65,p66,p71,p72 expression vector transfecting by f l ow cytometry
3 討論
為探討Numb不同亞型在乳腺癌發生中的作用,本工作利用TALEN技術構建了Numb敲除的MCF-7穩定細胞系Numb-KO MCF-7.首先,在Numb基因敲除后,MCF-7細胞增殖和遷移能力明顯增強,進一步研究發現,Numb基因敲除后的MCF-7細胞S期和G2M期的比例增大,說明Numb可以阻滯MCF-7細胞由G0G1期往S期和G2M的轉變,從而抑制MCF-7細胞的分裂.E-cadherin是一類介導同源細胞間粘附的鈣離子依賴性跨膜糖蛋白,主要存在于人和動物上皮細胞中,對于維持上皮細胞形態和組織完整性發揮重要作用,同時抑制腫瘤的侵襲轉移,影響原發腫瘤的早期生長和繼發腫瘤的增殖生長.Numb敲除后的MCF-7細胞表面E-cadherin表達量降低,可能是腫瘤遷移能力增強的一個標志[13].Numb基因對MCF-7細胞的增殖和遷移的抑制,原因可能是由于Notch、Hedgehog等信號通路失去Numb蛋白的抑制從而被增強,p53穩定性和細胞內吞穩態平衡也可能受到影響,這與以往Numb基因在其他腫瘤細胞中的研究結果相吻合[14].
Numb基因的4種亞型的產生,是由于哺乳動物Numb基因存在轉錄后的選擇性剪切,在磷酸酪氨酸結合(phosphotyrosine-binding,PTB)和富含脯氨酸域(proline-rich region,PRR)結構域分別插入11個氨基酸(ERKFFKGFFGK)或48個氨基酸(ANGTDSAFHVLAKPAHTALAPVA MPVRETNPWAHAPDAANKEIAATCS),由此產生PTBSPRRS(p65),PTBLPRRS(p66),PTBSPRRL(p71)和PTBLPRRL(p72)[9-10,15]4種蛋白亞型.本工作通過4種Numb表達質粒載體p65,p66,p71和p72分別轉染Numb-KO MCF-7細胞,發現PTBLPRRL(p72)能夠將Numb-KO MCF-7細胞的細胞周期阻滯在G0G1期,從而對細胞的增殖有明顯的抑制作用,4種Numb基因亞型均對Numb-KO MCF-7細胞遷移能力有抑制作用,而且這4種亞型轉染后,均能提高Numb-KO MCF-7細胞表面E-cadherin的表達量.
過去曾有學者在不同的系統中對Numb各亞型進行研究,發現在原代培養的小鼠神經前體細胞中,過表達PRRL將促進增殖,同時將不影響或抑制分化,而過表達PRRS則促進分化,同時抑制或不影響增殖[16];在P19細胞系中,PRRS能促進分化,而PRRL則不能;在視黃酸的誘導下,PRRL將促進P19細胞的增殖,而PRRS則不能[9];在果蠅外視原基中,h-PTBSPRRL(h表示來自人)能促進神經上皮細胞增殖,同時不影響分化,h-PTBSPRRS則能促進神經上皮分化而抑制其增殖[17].本研究結果與之前的一些報道有些不同,究其原因可能是研究系統有差異,因為在不同的組織細胞中,不同的Numb蛋白亞型具有不同功能,這是Numb的一個主要特點.Numb蛋白發揮作用主要是通過PTB和PRR 2個結構域,這2個結構域使得Numb能夠介導其他蛋白的連接,從而發揮多種生理功能[18-20].Numb蛋白亞型p72與其他亞型相比,最主要的特點是在PTB和PRR 2個結構域中都有插入序列.Numb蛋白亞型p72對乳腺癌細胞增殖能力的抑制,可能是發揮維持Notch,Hedgehog和TP53等信號通路平衡的作用,而這一作用可能需要PTB和PRR結構域中的2個插入序列.而p65,p66,p71和p72這4種亞型均對MCF-7細胞遷移起到抑制作用,與Numb參與的細胞黏附、細胞遷移的功能有關[21],這一功能的發揮可能不需要PTB和PRR結構域中的2個插入序列.
Numb基因的不同蛋白亞型之間,不論是在發育時空的表達上,還是在功能方面,都有明顯的差異,這使得Numb蛋白的亞型研究非常復雜.在過去10多年里得到了一些有價值的結果,但由于各種研究采用的系統不同,無法得出一個統一的結論[22].這是由Numb蛋白本身的特點決定的:Numb作為一個銜接蛋白,能夠介導不同蛋白之間的連接,這使得Numb細胞選擇命運的一個工具在不同的發育時空背景下被反復使用.因此,對Numb蛋白不同亞型的研究,要精確到特定的發育階段和細胞類型.本工作對乳腺癌細胞系MCF-7中的Numb亞型作用進行了初步研究,但Numb亞型在腫瘤中的表達情況及生物學功能、各個亞型與Notch,Hedgehog等信號通路是如何作用的,都還值得在今后作進一步研究.
[1]PECE S,CONFALONIERI S,RR P,et al.Numb-ing down cancer by more than just a Notch[J].Biochimica et Biophysica Acta,2011,1815(1):26-43.
[2]WANG Z,LI S S C.Numb:a new player in EMT[J].Cell Adhesion and Migration,2010,4(2):176-179.
[3]DING X,MA M,TENG J,et al.Numb induces E-cadherin adhesion dissolution,cytoskeleton reorganization,and migration in tubular epithelial cells contributing to renal f i brosis[J].Current Molecular Medicine,2015,15(4):368-379.
[4]TOSONI D,ZECCHINI S,COAZZOLI M,et al.The Numb/p53 circuitry couples replicative selfrenewal and tumor suppression in mammary epithelial cells[J].Journal of Cell Biology,2015,211(4):845-862.
[5]PECE S,SERRESI M,SANTOLINI E,et al.Loss of negative regulation by Numb over Notch is relevant to human breast carcinogenesis[J].Journal of Cell Biology,2004,167(2):215-221.
[6]DI M L,FERRETTI E,GRECO A,et al.Numb is a suppressor of Hedgehog signalling and targets Gli1 for itch-dependent ubiquitination[J].Nature Cell Biology,2006,8(12):1415-1423.
[7]SHAO X,LIU Y,YU Q,et al.Numb regulates vesicular docking for homotypic fusion of early endosomes via membrane recruitment of Mon1b[J].Cell Research,2016,26(5):593-612.
[8]SUN M,ASGHAR S Z,ZHANG H.The polarity protein Par3 regulates APP traきcking and processing through the endocytic adaptor protein Numb[J].Neurobiology of disease,2016,93(9):1-11.
[9]DHO S E,FRENCH M B,WOODS S A,et al.Characterization of four mammalian Numb protein isoforms.Identif i cation of cytoplasmic and membrane-associated variants of the phosphotyrosine binding domain[J].Journal of Biological Chemistry,1999,274(46):33097-33104.
[10]錢文雨,李華順.Numb與神經發育[J].中國細胞生物學學報,2010(1):1-16.
[11]BANI-YAGHOUB M,KUBU C J,COWLING R,et al.A switch in Numb isoforms is a critical step in cortical development[J].Developmental Dynamics,2007,236(3):696-705.
[12]VERDI J M,BASHIRULLAH A,GOLDHAWK D E,et al.Distinct human Numb isoforms regulate diあerentiation vs.proliferation in the neuronal lineage[J].Proceedings of the National Academy of Sciences of USA,1999,96(18):10472-10476.
[13]SLEEMAN J P,THIERY J P.SNAPSHOT:the epithelial-mesenchymal transition[J].Cell,2011,145(1):162-162.
[14]GULINO A,DI M L,SCREPANTI I.The multiple functions of Numb[J].Experimental Cell Research,2009,316(6):900-906.
[15]KARACZYN A,BANIYAGHOUB M,TREMBLAY R,et al.Two novel human Numb isoforms provide a potential link between development and cancer[J].Neural Development,2010,5(12):1040-1048.
[16]VERDI J M,BASHIRULLAH A,GOLDHAWK D E.Distinct human Numb isoforms regulate diあerentiation vs.proliferation in the neuronal lineage[J].Proceedings of the National Academy of Sciences,1999,96(18):10472-10476.
[17]TORIYA M,TOKUNAGA A,SAWAMOTO K,et al.Distinct functions of human Numb isoforms revealed by misexpression in the neural stem cell lineage in the Drosophila larval brain[J].Developmental Neuroscience,2005,28(1/2):142-155.
[18]GUO M,JAN L Y,JAN Y N.Control of daughter cell fates during asymmetric division:interaction of Numb and Notch[J].Neuron,1996,17(1):27-41.
[19]DHO S E,JACOB S,WOLTING C D,et al.The mammalian Numb phosphotyrosine-binding domain.Characterization of binding specif i city and identif i cation of a novel PDZ domain-containing Numb binding protein,LNX[J].Journal of Biological Chemistry,1998,273(15):9179-9187.
[20]WILLIAMSON M P.The structure and function of proline-rich regions in proteins[J].Biochemical Journal,1994,297(2):249-260.
[21]BERDNIK D,TOROK T G M,KNOBLICH J A.The endocytic protein alpha-adaptin is required for Numb-mediated asymmetric cell division in Drosophila[J].Developmental Cell,2002,3(2):221-231.
[22]RASIN M R,GAZULA V R,BREUNIG J J,et al.Numb and Numbl are required for maintenance of cadherin-based adhesion and polarity of neural progenitors[J].Nature Neuroscience,2007,10(7):819-827.
猜你喜歡
中老年保健(2022年6期)2022-08-19 01:41:48
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中國生殖健康(2019年2期)2019-08-23 08:11:42
中國生殖健康(2019年6期)2019-01-06 09:20:12
中國生殖健康(2019年5期)2019-01-06 09:16:40
祝您健康(2018年5期)2018-05-16 17:10:16
