CCK8和PGE2對CVB體外攻擊人外周血pDC的調控①
2018-07-06 12:23:08賈苗苗齊莉莉劉文宣賈嫻嫻
中國免疫學雜志 2018年6期
關鍵詞:檢測
賈苗苗 齊莉莉 李 慧 高 翔 劉文宣 賈嫻嫻
(河北醫科大學基礎醫學院病原生物學教研室,石家莊 050017)
樹突狀細胞(Dendritic cells,DC)是機體功能最強的專職抗原遞呈細胞,能通過模式識別受體高效地攝取、加工處理和遞呈抗原,繼而激活na?ve T 細胞[1]。DC處于啟動、調控、并維持免疫應答的中心環節,DC失控將會導致嚴重的疾病狀態[2-4]。Toll樣受體(Toll-like receptors,TLR)是一類重要模式識別受體,人類CD1c-CD123highCD303+CD304+漿細胞樣DC(plasmacytoid dendritic cells,pDC)選擇性表達TLR7和TLR9[5]。CD1c+CD123low傳統的DC(conventional DC,cDC)包括循環中的髓樣DC(myeloid dendritic cells,mDC)表達TLR1、TLR2、 TLR3、TLR4、TLR5、TLR6、TLR7[6],和在體外產生的單核細胞來源的DC(moDC)表達TLR1、TLR2、 TLR3、TLR4、TLR5、TLR8[7]。在人類,TLR9只在pDC上表達,一旦暴露到病毒觸發器,pDC快速生產大量的Ⅰ型IFN[8],不僅可以直接抑制病毒的復制,而且還誘導NK細胞、B細胞、T細胞、cDC的激活,誘導和放大抗病毒免疫應答。恰當的固有免疫應答可以防止病原體的感染,維持機體的健康,但是TLR信號的無限制活化將會引起機體功能的嚴重紊亂,導致嚴重的免疫病理損傷。因此,TLR信號系統必須得以嚴格的控制,以達到免疫穩態和避免免疫相關的病理損傷。大量研究表明柯薩奇病毒B(Coxsackie virus B,CVB)觸發的pDC活化在導致病毒性心肌炎患者嚴重組織病理損傷中發揮了關鍵作用[9,10],但目前關于pDC功能調控機制的研究卻并不深入。
膽囊收縮素(Cholecystokinin,CCK)是一種典型的腦腸肽和免疫調節肽。在體內以多種分子形式存在,八肽膽囊收縮素(Cholecystokinin octapeptide,CCK8)是體內含量最高且具有CCK全部生物活性的最小分子形式。它通過內分泌、旁分泌以及自分泌等多種方式作用于靶細胞,與CCK1受體(CCK1R)或CCK2受體(CCK2R)結合,介導相應的適應性細胞保護作用。近幾年來,本課題組做了許多有關CCK8抗內毒素休克和抗炎維持機體內穩態的研究工作[11-13]。在炎癥刺激因素作用下,單核細胞、巨噬細胞、成纖維細胞等多種細胞可以分泌PGE2,這表明體內炎癥反應局部的pDC是暴露于PGE2的環境中的。PGE2可以調節DC的多種生物學功能,比如成熟細胞因子的生產、T細胞的活化和凋亡。所以,本研究旨在體外模擬體內,探討PGE2存在時免疫調節肽CCK8能否參與調控柯薩奇病毒B誘導的人外周血漿細胞樣樹突狀細胞(pDC)的活化,以期對CVB導致的病毒性心肌炎的治療提供新的思路。
1 材料與方法
1.1實驗材料、試劑 CCK8、PGE2購自Sigma公司,CVB為本實驗室保存,Human CD304 MicroBeads、LS Columns購自Miltenyi Biotec公司,重組人白細胞介素3(Recombinant human interleukin 3,IL-3)購自Pepro Tech公司,Rabbit anti-human CCK1R及Mouse anti-human CCK2R購自Santa Cruz公司,FITC anti-human CD303、PE anti-human CD1c、PE/Cy5 anti-human CD123、FITC anti-human CD80、FITC anti-human CD86和FITC anti-human MHC class Ⅱ以及其所需相應的同型對照抗體均購自BD公司,PrimescriptTMRT reagent kit、SYRP Premix Ex TagTMKit購自大連寶生物公司,TRIzol Reagent購自Invitrogen公司,Human IFN-α ELISA Kit購自eBioscience公司。
1.2方法
1.2.1細胞來源及培養 新鮮分離的健康志愿者抗凝外周血白細胞由河北省血液中心提供,通過淋巴細胞分離液密度梯度離心,獲得外周血單個核細胞(PBMC),加入 Human CD304 MicroBeads,采用 mini MACS 免疫磁珠分離系統分離純化CD304+pDC。分離純化的pDC在含IL-3(10 ng/ml)的RPMI1640完全培養基中,37℃ 5%CO2,培養4 d,分化為靜息狀態的pDC,然后離心棄上清,加入CVB(100 TCID50)攻擊pDC 30 min,離心棄上清后加入含有CCK8或PGE2的完全培養基培養24 h。具體分為五組:control組;CVB組;CVB+CCK8組;CVB+PGE2組;CVB+PGE2+CCK8組。
1.2.2流式細胞技術檢測pDC純度 收集Human CD304 MicroBeads 分選的pDC,用PBS洗1次,分別加入FITC anti-human CD303、PE anti-human CD1c、PE/Cy5 anti-human CD123抗體,4℃避光孵育30 min,PBS洗2遍,流式細胞儀檢測鑒定CD1c-CD123highCD303+pDC細胞純度。
1.2.3RT-PCR檢測膽囊收縮素受體mRNA的表達 收集免疫磁珠分選純化的pDC和經CVB 體外攻擊的pDC,根據Trizol說明書提取細胞總RNA,檢測RNA純度和完整性后反轉錄成cDNA,以cDNA為模板,在ABI 7500 Real-time PCR儀上進行實時熒光定量PCR反應,檢測CCK1R和CCK2R mRNA的表達;瓊脂糖凝膠電泳及測序驗證PCR產物的特異性。引物由上海生物技術工程有限公司合成如表1。
1.2.4免疫熒光技術檢測膽囊收縮素受體的表達 收集免疫磁珠分選純化的pDC和經CVB體外感染的pDC,4%多聚甲醛固定,分別加入一抗(兔抗人CCK1R抗體和鼠抗人CCK2R抗體),二抗(FITC標記的山羊抗兔和texas red標記的山羊抗小鼠IgG),另外采用DAPI復染細胞核,PBS代替一抗作陰性對照,PC12細胞做陽性對照細胞。甘油封片后,于激光共聚焦顯微鏡下采集圖像,觀察CCK1R和CCK2R 蛋白的表達。
1.2.5流式細胞術檢測細胞表型 分組培養pDC,control組;CVB組;CVB+PGE2(10-7mol/L)組;CVB+CCK8(10-8mol/L)組;CVB+PGE2(10-7mol/L)+ CCK8(10-8mol/L)組,收集不同處理組的pDC,用PBS洗1次,分別加入 FITC anti-human CD80、FITC anti-human CD86、FITC anti-human MHC class Ⅱ和FITC anti-human CCR7抗體,4℃避光孵育30 min,PBS洗2遍,流式細胞儀檢測。每次實驗用同型抗體作為對照。用平均熒光強度(mean)表示CD80、CD86、MHC Ⅱ和CCR7蛋白表達量。
1.2.6ELISA技術檢測細胞因子的含量 純化pDC按上述不同因素處理培養24 h后,收集培養上清,按ELISA試劑盒說明書操作流程檢測人IFN-α的含量。
1.2.7RT-PCR檢測IFN-α mRNA的表達 收集不同處理組的pDC,根據Trizol說明書提取細胞總RNA,檢測RNA純度和完整性后反轉錄成cDNA,以cDNA為模板,在ABI 7500 Real-time PCR儀上進行實時熒光定量PCR反應,檢測IFN-α mRNA的表達。引物由上海生物技術工程有限公司合成,如表2。
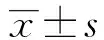
2 結果
2.1CCK受體在pDC上的表達 免疫磁珠分選純化的pDC,流式細胞儀檢測結果顯示CD303陽性細胞大于99%,同時高表達CD123,提示CD1c-CD123highCD303+pDC純度大于99%,結果如圖1所示。分選獲得的pDC在含IL-3(10 ng/ml)的RPMI1640完全培養基中培養4 d,分化為靜息狀態的pDC,然后給予CVB(100 TCID50)體外攻擊30 min。收集細胞,檢測不同狀態的pDC CCK1R和CCK2R表達。經統計學分析發現,CCK1R、CCK2R蛋白表達及mRNA表達在CVB體外攻擊后均增加(P<0.05 vs control),結果如圖2、3所示,提示CCK8系統可能參與了CVB體外攻擊人外周血pDC的調控。然而,我們在人外周血pDC中并沒有檢測到CCK mRNA,也未發現CCK蛋白的表達。提示,CCK8并非通過自分泌而是旁分泌或內分泌的方式作用于pDC,參與免疫調節作用。
2.2CCK8和PGE2對CVB誘導的協同刺激分子表達的影響 未經CVB誘導的pDC表達較低水平的CD80、CD86及HLA-DR;經CVB誘導的pDC協同刺激分子CD80、CD86和HLA-DR的表達顯著增高(P<0.05);CCK8(10-8mol/L)可以抑制CVB誘導的CD80、CD86和HLA-DR的表達,提示CCK8可抑制CVB誘導的pDC表型成熟(P<0.05 vs CVB組);然而PGE2(10-7mol/L)對CVB誘導的協同刺激分子CD80、 CD86和HLA-DR的表達增高有促進作用(P<0.05 vs CVB組)。提示CCK8和PGE2雙向調控CVB誘導的pDC協同刺激分子的表達,結果如圖4所示。

圖1 流式細胞技術檢測pDC純度Fig.1 Detect purity of pDC by flow cytometryNote:The purity of CD1c-CD123highCD303+pDC was 99%.
表1CCK1R和CCK22R引物序列
Tab.1PrimersequencesofCCK1RandCCK2R

GenesAccession No.PrimersCCK1RNM_000730.2Forward primer:5'-GACGCTTCGGTCATTAGA-3'Reverse primer:5'-AGGGAGGAGTGAAGTTGC-3'CCK2RNM_176875.2Forward primer:5'-CCCACTCCCTCCATTGCT-3'Reverse primer:5'-CTGCTCCATTCTTATTCCTCTT-3'β-actinNM_031144Forward primer:5'-GGACTTCGAGCAAGAGATGG-3'Reverse primer:5'-TGTGTTGGCGTACAGGTCTTTG-3'
表2IFN-α引物序列
Tab.2PrimersequencesofIFN-α

GenesAccession No.PrimersIFN-αNM_002171Forward primer:5'-GGAGGAAGGATTGAAAACTGG-3'Reverse primer:5'-GATGAAGTGAGACATCAGAATGG-3'
2.3CCK8和PGE2對CVB誘導趨化因子受體表達的影響 未經CVB誘導的pDC表達較低水平的CCR7;經CVB誘導的pDC協同刺激分子CCR7的表達顯著增高(P<0.05);CCK8(10-8mol/L)可以促進CVB誘導的CCR7的表達,提示CCK8可促進CVB誘導的pDC的趨化(P<0.05 vs CVB組);PGE2(10-7mol/L)對CVB誘導的趨化因子受體CCR7的表達增高也有促進作用(P<0.05 vs CVB組)。提示CCK8和PGE2均可促進CVB誘導的pDC趨化,結果如圖5所示。
2.4CCK8和PGE2對CVB誘導pDC產生IFN-α的調控 收集不同因素作用后的各組細胞的培養上清,用ELISA法檢測IFN-α的含量,RT-PCR檢測IFN-α mRNA的表達。未經CVB誘導的pDC,表達較低水平的IFN-α mRNA;經CVB誘導的pDC,IFN-α mRNA的表達顯著增高(P<0.05);CCK8可以劑量依賴性抑制IFN-α mRNA表達的增高(P<0.05 vs CVB組),且對pDC存活無明顯影響;PGE2(10-7mol/L) 對CVB誘導IFN-α mRNA的表達有促進作用(P<0.05 vs CVB組)。同時,培養上清中IFN-α含量檢測結果也證明體外給予CCK8(10-8mol/L),可以抑制CVB誘導的pDC分泌IFN-α(P<0.05 vs CVB組),相反PGE2可以促進pDC分泌IFN-α(P<0.05 vs CVB組)。提示CCK8和PGE2參與了CVB誘導pDC產生Ⅰ型干擾素的雙向調控作用,結果如圖6所示。

圖2 RT-PCR檢測CCK1R和CCK2R mRNA的表達Fig.2 Detect CCK1R and CCK2R mRNA by RT-PCRNote:The mRNA expression of CCK1R and CCK2R was up-regulated by CVB attack.*.P<0.05.


圖3免疫熒光技術檢測CCK1R和CCK2R蛋白的表達
Fig.3ExpressionofCCK1RandCCK2Rproteindetectedbyimmunofluorescencetechnique
Note: Green fluorescence stands for CCK1R protein;Red fluorescence stands for CCK2R protein.A.Control group;B.CVB group;C.Control group;D.CVB group.The expression of CCK1R and CCK2R was up-regulated by CVB attack.*.P<0.05 vs Control,t-test.

圖4 流式細胞技術檢測協同刺激分子的表達Fig.4 Expression of costimulatory molecules detect by flow cytometryNote:CCK8 repressed the expression of costimulatory molecules on CVB-attacked pDC.P<0.05,CVB+CCK8 vs CVB,t-test,while PGE2 promoted it(*.P<0.05).

圖5 流式細胞技術檢測CCR7的表達Fig.5 Expression of CCR7 detected by flow cytometryNote:CCK8 and PGE2 promoted the expression of CCR7 on CVB-attacked pDC.*.P<0.05.

圖6 RT-PCR和ELISA 法檢測IFN-α的合成和分泌Fig.6 Synthesis and secretion of IFN-α detected by RT-PCR and ELISANote:A.Exogenous CCK8 inhibited the IFN-α synthesis,dose-dependently,in vitro;B.CCK8(10-8mol/L)inhibited CVB induced IFN-α mRNA expression,while PGE2 promoted;C.CCK8(10-8mol/L)inhibited CVB induced IFN-α secretion,while PGE2 promoted.*.P<0.05.
3 討論
CVB是一種常見的腸道病毒,感染后大部分患者表現為隱性感染或只表現為一般感冒癥狀,少數患者則表現為嚴重的心肌炎,甚至導致猝死,也可發展為擴張性心肌病[14]。有研究證實,CVB 感染導致的心肌炎并非由CVB 直接破壞心肌細胞引起,很大程度上是由于過強的免疫應答導致的心肌損傷[15]。因此,不同的臨床表現與不同的免疫反應有關。病毒感染,可以激活漿細胞樣樹突狀細胞(pDCs),并迅速產生大量Ⅰ型IFN,這不僅可以直接抑制病毒復制而且還可誘導NK細胞、B細胞、T細胞、cDCs發生免疫應答。但Ⅰ型IFN的大量產生和免疫系統的過度活化也將對機體組織造成免疫病理損傷[16]。CVB觸發的pDCs細胞活化在導致病毒性心肌炎患者嚴重組織病理損傷中發揮了關鍵作用,但目前關于pDCs細胞功能負向調控機制的研究卻并不深入。有研究提示餐后釋放的膽囊收縮素(Cholecystokinin,CCK)類物質,除了干預消化系統功能外,還能干預神經信號的轉導以及免疫應答反應,之間存在異常復雜的相互作用。CCK是一個激素家族,CCK8是擁有完整生物活性的最小片段。人外周血pDC是病毒感染的主要防御者。我們的研究證明體外給予CVB,刺激活化人外周血pDC,CCK1R和CCK2R的表達顯著升高,pDCs的活化對它們的表達有明顯的正向調節作用。CCK受體表達的增加可能是人體應激狀態下神經、內分泌、免疫系統的一種保護性機制或者是它們之間進行的信息傳遞。我們的研究結果表明作為一種免疫調節因子的CCK8,體外可以抑制CVB刺激誘導的pDCs表型的成熟,而且,CCK8會抑制CVB誘導的IFN-α的合成與分泌。體外由CCK8所誘導的對人pDC活化的調節,也許將為CVB感染后引起的病毒性心肌炎提供以調控免疫為基礎的新療法。機體存在精密的負向免疫調控機制以保證 TLR 信號的適度活化,CCK8因此將會被用于免疫應答調節,以防止TLR 通路過度活化引發的免疫應答紊亂。
在炎癥刺激因素作用下,單核細胞、巨噬細胞、成纖維細胞等多種細胞可以分泌PGE2,這表明體內炎癥反應局部的pDC是暴露于PGE2的環境中的。PGE2可以調節DC的多種生物學功能[17],比如成熟細胞因子的生產、T細胞的活化和凋亡。同時也觀察了在PGE2存在時CCK8對pDCs表型成熟的影響。結果表明,體外PGE2單獨作用可以促進抑制CVB刺激誘導的pDCs表型的成熟和IFN-α的合成與分泌。而CCK8(10-8mol/L)能抑制PGE2(10-7mol/L)對CVB誘導的pDC協同刺激分子CD80、CD86和HLA-DR的表達以及IFN-α的合成與分泌的促進作用。提示CCK8和PGE2可以雙向調控CVB誘導pDC的免疫應答水平。
CCK8作為一種免疫抑制劑或細胞激素參與調控CVB誘導的人類pDCs活化,從而防止發生免疫過度引發的免疫病理損傷,這已非常清楚。我們前期研究結果證明CCK8可以通過CCK2R調節重要的信號分子TRAF6的表達,進而抑制TLR9誘導的pDC的活化。TRAF6是NF-κB、MAPK、IRF7等激活過程中重要的銜接蛋白,CCK受體屬于G蛋白偶聯受體(GPCR),TLR-MyD88-TRAF6和GPCR-PI3K/PKA/PKC/CAMK信號通路之間的相互聯系機制尚未明了,這需要更深入的研究來解決。但所有這些結果提醒我們,CCK8對于pDCs免疫功能的調節不容忽視,這將為CVB 感染導致的心肌炎提供新的治療手段。
參考文獻:
[1] Benvenuti F.The dendritic cell synapse:a life dedicated to T cell activation[J].Front Immunol,2016,7:70.
[2] 葉麗萍,胡靜濤,王春鳳.樹突狀細胞與輪狀病毒相互作用機制研究進展[J].中國免疫學雜志,2017,33(6):947-950.
Ye LP,Hu JT,Wang CF.Recent progress in the mechanism of interaction between dendritic cells and rotavirus[J].Chin J Immunol,2017,33(6):947-950.
[3] McGovern N,Shin A,Low G,etal.Human fetal dendritic cells promote prenatal T-cell immune suppression through arginase-2[J].Nature,2017,546(7660):662-666.
[4] Anguille S,Van de Velde AL,Smits EL,etal.Dendritic cell vaccination as postremission treatment to prevent or delay relapse in acute myeloid leukemia[J].Blood,2017,130(15):1713-1721.
[5] Macri C,Pang ES,Patton T,etal.Dendritic cell subsets[J].Semin Cell Dev Biol,2017,doi:10.1016/j.semcdb.2017.12.009.
[6] Patel VI,Metcalf JP.Identification and characterization of human dendritic cell subsets in the steady state:a review of our current knowledge[J].J Investig Med,2016,64(4):833-847.
[7] Alculumbre S,Pattarini L.Purification of human dendritic cell subsets from peripheral blood[J].Methods Mol Biol,2016,1423:153-167.
[8] Stead SO,McInnes SJP,Kireta S,etal.Manipulating human dendritic cell phenotype and function with targeted porous silicon nanoparticles[J].Biomaterials,2018,155:92-102.
[9] Hamalainen S,Nurminen N,Ahlfors H,etal.Coxsackievirus B1 reveals strain specific differences in plasmacytoid dendritic cell mediated immunogenicity[J].J Med Virol,2014,86(8):1412-1420.
[10] Wu F,Fan X,Yue Y,etal.A vesicular stomatitis virus-based mucosal vaccine promotes dendritic cell maturation and elicits preferable immune response against coxsackievirus B3 induced viral myocarditis[J].Vaccine,2014,32(31):3917-3926.
[11] Jia X,Cong B,Zhang J,etal.CCK8 negatively regulates the TLR9-induced activation of human peripheral blood pDCs by targeting TRAF6 signaling[J].Eur J Immunol,2014,44(2):489-499.
[12] Zhang JG,Liu JX,Jia XX,etal.Cholecystokinin octapeptide regulates the differentiation and effector cytokine production of CD4(+)T cells in vitro[J].Int Immunopharmacol,2014,20(2):307-315.
[13] Zhang D,Li H,Geng J,etal.The therapeutic effects of cholecystokinin octapeptide on rat liver and kidney microcirculation disorder in endotoxic shock[J].Immunopharmacol Immunotoxicol,2017,39(1):2-10.
[14] Liao Y,Chen KH,Dong XM,etal.A role of pre-mir-10a coding region variant in host susceptibility to coxsackie virus-induced myocarditis[J].Eur Rev Med Pharmacol Sci,2015,19(18):3500-3507.
[15] Cen Z,Guo Y,Kong Q,etal.IL-10-producing B cells involved in the pathogenesis of Coxsackie virus B3-induced acute viral myocarditis[J].Int J Clin Exp Pathol,2015,8(1):830-835.
[16] Teijaro JR.Type I interferons in viral control and immune regulation[J].Curr Opin Virol,2016,16:31-40.
[17] Maric J,Ravindran A,Mazzurana L,etal.PGE2 suppresses human group 2 innate lymphoid cell function[J].J Allergy Clin Immunol,2018,141(5):1761-1773.
猜你喜歡
中國設備工程(2022年12期)2022-07-11 04:33:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:36
中學生數理化·七年級數學人教版(2019年9期)2019-11-25 07:34:34
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:50
中學生數理化·七年級數學人教版(2019年12期)2019-05-21 02:53:48
