miR-140表達與腫瘤發生發展關系的研究進展
2018-07-12 07:30:30李維娜何飛
解放軍醫學雜志 2018年6期
李維娜,何飛
MicroRNA是真核生物中的一類小分子非編碼RNA,能調控約1/3人類基因的表達,在細胞發育、分化、增殖、凋亡和腫瘤發生發展等生理及病理過程中都發揮著重要作用[1-7]。miR-140最早被發現表達于軟骨細胞,能夠調控軟骨的發育[8-9]。近年來越來越多的研究發現miR-140在多種腫瘤中呈低表達,可能發揮抑癌基因的作用[10]。
1 miR-140概述
MicroRNA主要通過結合mRNA的3'非翻譯區(UTR)來影響mRNA的降解或翻譯,進而在轉錄后水平調控基因的表達。microRNA的5'端6~9個核苷酸的種子序列(seed sequence)與mRNA的3'-UTR形成不完全互補,因此一個種子序列可能與數百個mRNA結合,調控數百個靶基因的表達。microRNA由Ⅱ型RNA聚合酶合成,首先在核內轉錄成初級轉錄本miRNA(pri-miRNA),后者經Drosha核糖核酸酶剪切加工后形成約70nt大小、呈莖環樣結構的前體miRNA(pre-miRNA),pre-miRNA在轉運蛋白Exportin-5和Ran-GTP的作用下轉運出細胞核。在核糖核酸內切酶Dicer的作用下,pre-miRNA被加工為大小約20~23個堿基對的雙鏈RNA,后者與RNA誘導的沉默復合物(RISC)結合之后解開雙鏈,此時將靠近5'端的一條RNA鏈稱為miR-X-5p,而靠近3'端的互補鏈稱為miR-X-3p。通常情況下,microRNA只有一條RNA鏈豐度高,而另一條則被降解[2-4]。但是,人miR-140與大多數microRNA不同,有miR-140-5p和miR-140-3p兩種形式(圖1),二者表達豐度基本相當[11],其內在的分子機制目前仍不清楚。
miR-140位于人染色體16q22.1上,該位點是染色體的脆性部位(圖2)。研究發現,miR-140位于E3泛素連接酶WWP2的內含子中,成熟miR-140與WWP2-C的剪接體基因共表達,受其啟動子調控[12]。
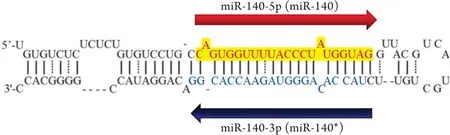
圖1 pre-miR-140的莖環結構以及兩種成熟形式Fig.1 A stem-loop structure and two mature forms of pre-miR-140

圖2 miR-140在人類染色體的位置Fig. 2 Schematic diagram of the position of miR-140 gene
2 miR-140與腫瘤
2.1 miR-140與乳腺腫瘤 miR-140與乳腺癌密切相關。在早期乳腺癌中,Li等[13]比較了正常乳腺組織和不同惡性程度乳腺導管內原位癌(DCIS)miRNA表達譜的差異,發現DCIS中miR-140-3p表達降低,且DCIS惡性程度越高,miR-140-3p表達越低;進一步研究發現,在雌激素受體陰性(ERα–)的基底樣癌中,腫瘤干細胞(CD44+CD24–)的miR-140-3p表達水平低于乳腺干細胞;在乳腺癌發生模型MCF10中,過表達miR-140-3p可抑制克隆形成和細胞增殖,抑制腫瘤干細胞的自我更新和腫瘤生長,而敲低miR-140-3p則能促進腫瘤細胞的增殖、抑制細胞凋亡。研究發現,在ERα–的乳腺癌腫瘤干細胞中,miR-140-3p低表達與DNA的異常甲基化有關。miR-140啟動子區CpG島的甲基化程度明顯增高,用DNA甲基轉移酶抑制劑處理后,miR-140表達水平升高。此外,對miR-140-3p的下游靶基因進行分析發現,miR-140-3p可能是通過調控干細胞調節因子人性別決定區Y框蛋白9(SOX9)和乙醛脫氫酶1(ALDH1)的表達來影響腫瘤干細胞生長的[13]。
同時,在雌激素受體陽性(ERα+)的侵襲性和非侵襲性乳腺癌中,miR-140-3p的表達水平亦下降[14]。miR-140-3p能調控雌激素受體(ERα)作用下乳腺癌腫瘤干細胞的自我更新,從而發揮抑癌基因作用。ERα通過雌二醇(E2)結合于miR-140-3p啟動子區的雌激素應答元件(ERE),抑制miR-140-3p的表達,進而導致miR-140-3p的下游靶基因干細胞調節因子SOX2表達升高。
與Li等[13]的研究類似,Yoshida等[15]亦發現miR-140能調控乳腺腫瘤干細胞的干性,發揮抑制腫瘤的作用。他們發現抑癌基因Rb通過正性調節miR-140的表達,抑制白細胞介素6(IL-6)的表達,而IL-6能促進腫瘤干細胞、胚胎干細胞的自我更新[16]。但是Rb如何調控miR-140的表達,其機制尚未闡明。
外泌體是調節腫瘤干細胞龕活性的一個重要途徑[17]。Gernapudi等[18]發現紫草素(SK)能使乳腺脂肪前體細胞外泌體中的miR-140含量明顯增加,從而抑制乳腺腫瘤干細胞的自我更新和腫瘤生長,發揮抑癌作用。此外,miR-140也能作為判斷乳腺癌患者預后的指標之一。Chang等[19]發現高表達miR-140-3p的患者預后好。
2.2 miR-140與肝臟腫瘤 肝癌是最常見的腫瘤之一,我國肝癌發病率居世界首位[20]。肝癌的發生與肝臟的慢性炎性反應密切相關,與核轉錄因子-kappa B(NF-κB)信號傳導失調有關[21]。Takata等[22]發現miR-140-3p能抑制NF-κB共激活因子NRIP的表達,從而抑制NF-κB的信號活性。Takata等[23]通過進一步建立二乙基亞硝胺(DEN)誘導肝細胞肝癌(HCC)模型,發現miR-140敲除小鼠更容易出現HCC。分子機制研究表明,蛋白質-RNA復合物miRNP蛋白DEAD框解旋酶20(DDX20)表達降低可導致miR-140-3p功能障礙,使其下游靶基因Dnmt1表達增高,金屬硫蛋白MTs啟動子區CpG島甲基化,MTs表達下降,NF-κB信號活性增強,從而促進腫瘤的發生[23]。
除miR-140-3p在肝癌中發揮抑癌基因作用外,miR-140-5p也可抑制肝癌細胞的生長。Yang等[24]發現在肝癌組織和多種肝癌細胞系中miR-140-5p表達降低。他們發現miR-140-5p通過調控下游靶基因轉化生長因子β受體1(TGFBR1)和成纖維細胞生長因子9(FGF9),調節轉化生長因子β(TGF-β)和絲裂原活化蛋白激酶(MAPK)信號通路,從而影響肝癌細胞的轉移和增殖。Yan等[25]發現miR-140-5p可作用于下游靶基因脯胺酰異構酶Pin1,降低蛋白激酶B(Akt)和細胞外調節蛋白激酶(ERK)的磷酸化,阻斷多種促癌信號的激活。Lv等[26]發現乙型肝炎病毒(hepatitis B virus,HBV)能誘導肝癌細胞非編碼長鏈RNA(lncRNA)Unigene56159的表達,而Unigene56159是miR-140-5p的競爭性內源RNA(ceRNA),拮抗miR-140-5p行使功能,因此,HBV可通過Unigene56159/miR-140-5p/Slug途徑促進HCC的侵襲和轉移。
miR-140-5p不僅在HCC中發揮抑癌作用,同樣也在膽管細胞癌中發揮抑制腫瘤的作用。研究發現,miR-140-5p在膽管細胞癌中呈低表達,其靶基因腫瘤促進基因(tumor promoting gene)Septin2表達上調[27]。
2.3 miR-140與肺癌 近年來,研究者對miR-140在肺癌中的作用機制展開了一系列研究。Yanaihara等[28]早在2006年就比較了肺癌組織與正常肺組織的microRNA表達譜,從中發現43種表達異常的microRNA,其中肺癌組織中miR-140-5p表達下調。Tan等[29]進一步比較了肺組織與肺鱗狀細胞癌(SCC)的microRNA表達譜,發現了5個可以區分正常組織與癌組織的具有差異表達特性的microRNA,miR-140-3p即為其中之一,它在SCC中表達降低。Kong等[30]和Dong等[31]亦發現miR-140-3p在肺癌組織中低表達,而其過表達可抑制肺癌細胞的增殖、侵襲和遷移,其靶基因為ATP6AP2和ATP8A1。Yuan等[32]發現miR-140-5p在非小細胞肺癌組織和細胞系中低表達,miR-140-5p的低表達導致其靶基因胰島素樣因子1受體(IGF1R)高表達,從而促進腫瘤細胞的增殖、侵襲和轉移。Zhang等[33]發現Smad3是miR-140-5p的靶基因,肺癌細胞中miR-140-5p的低表達導致pSmad3活性增強,后者不僅促進上皮間質轉化(EMT),促進腫瘤細胞的侵襲、轉移,同時也作用于癌基因Trib2啟動子,增加Trib2的表達,促進肺癌細胞增殖。
本課題組研究發現,單核細胞向巨噬細胞分化相關基因(MMD)在肺癌組織中高表達,且與miR-140-5p的表達呈負相關,而進一步研究表明MMD是miR-140-5p的靶基因[34]。miR-140-5p通過調控MMD的表達而影響細胞增殖相關信號途徑MAPKERK1/2的活性[34]。
Izzotti等[35]通過建立大鼠吸煙模型,發現吸煙能使大鼠肺組織中miR-140的表達下降,因此推測miR-140是吸煙導致肺癌的分子機制之一。
2.4 miR-140與消化道腫瘤 miR-140與各種消化道腫瘤的發生、發展、侵襲和轉移密切相關。在食管癌中,Li等[36]發現,與相鄰正常的食管組織相比,miR-140-5p在食管癌組織中表達水平降低。進一步研究表明,在食管癌細胞中,miR-140-5p可通過調控靶基因Slug的表達來影響腫瘤細胞的侵襲和轉移。
Liang等[37]發現,miR-140-5p在胰腺癌組織和細胞系中表達明顯下降,在人胰腺癌PANC-1細胞中,過表達的miR-140-5p能削弱腫瘤細胞的增殖和侵襲能力。同時,他們的研究還發現,miR-140-5p能作用于靶基因凋亡刺激蛋白P53的抑制因子(iASPP),過表達miR-140-5p會降低PANC-1中iASPP、腫瘤蛋白63(ΔNp63)、基質金屬蛋白酶2/9(MMP2/9)的水平,從而發揮抑癌作用。
Zou等[38]發現,在胃癌組織和多種胃癌細胞系中存在miR-140-5p表達降低的情況,在人胃癌細胞HGC-27中過表達miR-140-5p能抑制腫瘤細胞存活和克隆形成,導致G0/G1期阻滯,推測其分子機制可能是miR-140-5p作用于靶基因SOX4,從而抑制了胃癌細胞的增殖。
Piepoli等[39]通過microRNA表達譜篩選了19例結腸癌樣本中差異表達的microRNA,發現4個失調的miRNA,即miR-195、miR-1280、miR-140-3p和miR-1246,其中miR-140-3p表達下調。Zhang等[40]發現結腸癌樣本和細胞系中miR-140-5p的表達下調,通過靶基因血管內皮生長因子A(VEGFA)影響腫瘤細胞的增殖、侵襲,且miR-140-5p低表達的程度與結腸癌的惡性程度及患者生存率密切相關。與此研究相似,Yu等[41]發現,結腸癌中的miR-140-5p表達下降,但不同的是,miR-140-5p可能是通過靶基因血小板反應蛋白解整合素金屬肽酶5(ADAMTS5)和胰島素樣生長因子結合蛋白5(IGFBP5)來影響結腸癌細胞的侵襲和遷移的。趙文月等[42]認為miR-140-5p通過作用于靶基因蠕蟲果蠅母抗同源蛋白3(Smad3)來影響結腸癌細胞的侵襲和轉移。Zhai等[43]研究發現miR-140-5p還能調控結腸癌腫瘤干細胞(CSC,CD44highCD133high)的功能。體外實驗發現,過表達miR-140-5p能明顯抑制CSC的生長和干細胞球的形成,而體內研究則發現過表達miR-140-5p可抑制結腸癌CSC的成瘤和轉移[43]。他們認為miR-140-5p可通過靶基因Smad2來調節結腸癌的增殖和侵襲、轉移。此外他們還發現miR-140-5p可使自噬相關蛋白12(ATG12)的表達降低,從而干擾CSC的自噬能力。盡管miR-140-5p能抑制結腸癌CSC的增殖能力,但Song等[44]認為miR-140-5p可導致CSC對5-氟尿嘧啶等細胞周期化療藥物的敏感性降低。
2.5 miR-140與骨肉瘤 miR-140最早被發現高表達于軟骨細胞,其與軟骨的發育密切相關[8-9]。但miR-140與骨腫瘤的關系尚不清楚。Xiao等[45]發現,與正常骨組織相比,骨肉瘤中miR-140-5p的表達下調,過表達miR-140-5p能抑制骨肉瘤細胞的增殖和遷移。miR-140-5p能通過作用于靶基因組蛋白去乙酰化酶4(HDAC4)抑制骨肉瘤細胞的增殖。Song等[44]發現,miR-140-5p可誘導p53和p21蛋白的表達并作用于靶基因HDAC4,使骨肉瘤細胞在G1和G2期阻斷,抑制細胞增殖。同時,體外研究發現,由于miR-140-5p抑制細胞周期的作用,導致骨肉瘤細胞對5-氟尿嘧啶等細胞周期特異性化療藥物不敏感。但與此不同的是,Meng等[46]發現,在對化療藥物抵抗的骨肉瘤組織標本中miR-140-5p的表達明顯低于對化療藥物敏感的標本,故推測過表達miR-140-5p可增強骨肉瘤細胞對甲氨蝶呤等化療藥物的敏感性,其分子機制是miR-140-5p作用于靶基因高遷移率族核小體結合域蛋白5(HMGN5),從而抑制了細胞自噬,而自噬則是腫瘤化療藥物抵抗的重要機制之一[47]。
2.6 miR-140與其他腫瘤
2.6.1 頭頸部腫瘤 Sand等[48]用microRNA芯片技術分別篩選了皮膚基底樣細胞癌組織與正常組織、皮膚鱗狀細胞癌與正常組織中差異表達的microRNA,分別發現了10個和6個表達下調的microRNA,而miR-140-3p是其中之一。Jing等[49]發現,miR-140-5p在下咽癌腫瘤組織中低表達,并且與下咽癌患者腫瘤TN分期明顯相關,miR-140-5p能抑制下咽癌細胞的遷移和侵襲能力,這種作用可能是通過調控整合素金屬蛋白酶10(ADAM10)的表達,進而影響Notch1蛋白的激活來實現的。Kai等[50]在舌部鱗狀細胞癌中也發現miR-140-5p可通過ADAM10調控舌癌細胞的遷移和侵襲,同時還發現遷移相關基因HDAC7、層粘連蛋白Gamma 1(LAMC1)和配對盒基因6(PAX6)是miR-140-5p的靶基因。
2.6.2 神經系統腫瘤 膠質瘤是神經系統最常見的腫瘤。Liu等[51]發現,miR-140在膠質瘤樣本和細胞系中低表達,在膠質瘤細胞中過表達miR-140-5p能抑制細胞的增殖和侵襲,他們推測miR-140發揮此作用的靶基因可能是ADAM9。腫瘤相關的lncRNA H19在多種腫瘤中呈高表達[52],Zhao等[53]發現H19在膠質瘤中高表達,且能與miR-140-5p結合并抑制其功能,導致miR-140-5p下游靶基因iASPP的表達升高,而后者升高能夠促進腫瘤細胞存活。
2.6.3 血液腫瘤 Reddemann等[54]篩選了30例血管免疫母細胞性T細胞淋巴瘤(AITL)樣本的microRNA表達譜,發現miR-140-3p表達下調。Correia等[55]發現miR-140-5p在急性T淋巴細胞白血病(T-ALL)樣本和細胞系中低表達,其靶基因是與T-ALL發生密切相關的促癌基因T淋巴細胞白血病因子1(TAL1)。
2.6.4 泌尿生殖系統腫瘤 宮頸癌和卵巢癌都是女性的常見腫瘤。Su等[56]在宮頸癌標本中發現miR-140-5p和miR-140-3p都呈低表達,且miR-140-5p低表達與患者的預后密切相關,同時還發現miR-140-5p能通過靶基因IGF2BP1調控腫瘤細胞生長、侵襲和轉移。Iorio等[57]通過microRNA芯片技術比較了69例惡性卵巢癌樣本、15例正常卵巢樣本和5種卵巢癌細胞系中差異表達的microRNA,其中miR-140-5p表達下調。Miles等[58]通過microRNA芯片發現在卵巢癌樣本中miR-140-3p呈低表達。Lan等[59]進一步研究發現,部分miR-140-5p通過作用于靶基因血小板源性生長因子受體A(PDGFRA)而發揮抑制卵巢癌細胞生長的作用。Ingelmo-Torres等[60]發現,膀胱癌組織中miR-140-5p表達水平降低,且miR-140-5p的表達水平越低,膀胱癌的惡性程度越高。Wang等[61]進一步研究發現,過表達miR-140-5p可通過降低ΔNp63基因的表達來抑制膀胱癌的進展。
2.7 miR-140的促癌作用 盡管在大多數腫瘤中miR-140都能發揮抑制腫瘤生長、侵襲和轉移的作用,但也有相反的報道。研究發現miR-140-3p在脊索瘤中表達增高,增高的miR-140-3p與腫瘤復發和患者預后密切相關[62]。甲狀腺乳頭狀癌患者血清中miR-140-3p水平明顯高于甲狀腺良性結節患者[63]。
綜上所述,miR-140受多種因子調控,在多種腫瘤中呈低表達,無論是成熟的miR-140-5p還是miR-140-3p,在乳腺癌、肝癌、肺癌、消化道腫瘤、骨肉瘤等多數腫瘤中均發揮著抑癌基因作用。由表1可見,它們通過抑制多種下游靶基因的表達,調控腫瘤細胞的增殖、凋亡和侵襲轉移。
3 展 望
miR-140在大多數腫瘤中低表達,可能成為多種腫瘤的標志物之一,與其他指標相結合,可用于健康篩查、早期診斷和腫瘤復發的檢測。此外,檢測組織樣本或血液中miR-140的表達水平可能有助
于判斷腫瘤的惡性程度及預后情況。
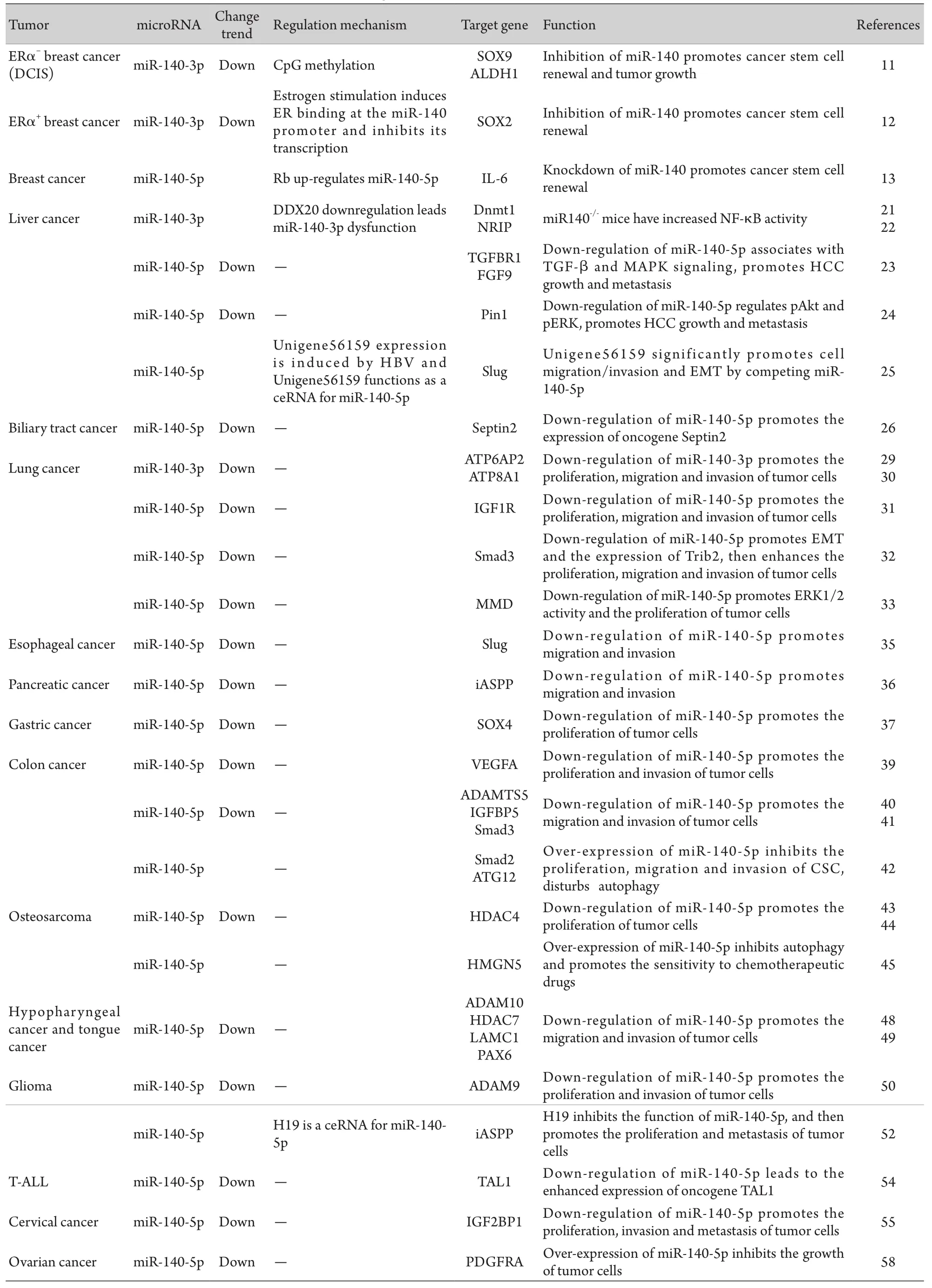
表1 miR-140在多種腫瘤中的表達及其下游靶基因和功能Tab.1 Expression, target and function of miR-140 in different tumors
miR-140在多種腫瘤中發揮抑癌基因的作用,有望成為腫瘤治療的靶標。通過調控miR-140來治療腫瘤的策略有:①外源性補充miR-140;②去除抑制miR-140表達的因素,促進其內源性表達;③抑制miR-140下游靶基因的表達或功能。總之,miR-140在多種腫瘤的發生發展過程中都發揮著重要作用,是一種重要的抑癌miRNA,但目前的研究仍處在初步探索階段,進一步深入研究的結果值得期盼。
[1] Ambros V. The functions of animal microRNAs[J]. Nature,2004, 431(7006): 350-355.
[2] Bartel DP. MicroRNAs: target recognition and regulatory functions[J]. Cell, 2009, 136(2): 215-233.
[3] Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function[J]. Cell, 2004, 116(2): 281-297.
[4] Fabian MR, Sonenberg N, Filipowicz W. Regulation of mRNA translation and stability by microRNAs[J]. Annu Rev Biochem,2010, 79: 351-379.
[5] Liu D, Gao NJ, Tian XX, et al. Effects of microRNA-124 on the proliferation and apoptosis of human umbilical vein endothelial cells[J]. Med J Chin PLA, 2016, 41(1): 7-11. [劉丹, 高乃婧, 田孝祥, 等. MicroRNA-124對人臍靜脈內皮細胞增殖和凋亡的影響[J]. 解放軍醫學雜志, 2016, 41(1): 7-11.]
[6] Cao D, Sun XF. Research progress of Micro RNA expression in renal fibrosis[J]. Chin J Pract Intern Med, 2017, 37(3): 262-266.[曹聃, 孫雪峰. MicroRNA在腎組織纖維化中的研究進展[J].中國實用內科雜志, 2017, 37(3): 262-266.]
[7] Liu D, Gao NJ, Tian XX, et al. Effects of microRNA-124 on the proliferation and apoptosis of human umbilical vein endothelial cells[J]. Med J Chin PLA, 2016, 41(1): 7-11. [劉丹, 高乃婧, 田孝祥, 等. MicroRNA-124對人臍靜脈內皮細胞增殖和凋亡的影響[J]. 解放軍醫學雜志, 2016, 41(1): 7-11.]
[8] Lagos-Quintana M, Rauhut R, Yalcin A, et al. Identification of tissue-specific microRNAs from mouse[J]. Curr Biol, 2002,12(9): 735-739.
[9] Miyaki S, Sato T, Inoue A, et al. MicroRNA-140 plays dual roles in both cartilage development and homeostasis[J]. Genes Dev,2010, 24(11): 1173-1185.
[10] Green D, Dalmay T, Fraser WD. Role of miR-140 in embryonic bone development and cancer[J]. Clin Sci (Lond), 2015,129(10): 863-873.
[11] Rakoczy J, Fernandez-Valverde SL, Glazov EA, et al. MicroRNAs-140-5p/140-3p modulate Leydig cell numbers in the developing mouse testis[J]. Biol Reprod, 2013, 88(6): 143.
[12] Yang J, Qin S, Yi C, et al. MiR-140 is co-expressed with Wwp2-C transcript and activated by Sox9 to target Sp1 in maintaining the chondrocyte proliferation[J]. FEBS Lett, 2011, 585(19): 2992-2997.
[13] Li Q, Yao Y, Eades G, et al. Downregulation of miR-140 promotes cancer stem cell formation in basal-like early stage breast cancer[J]. Oncogene, 2014, 33(20): 2589-2600.
[14] Zhang Y, Eades G, Yao Y, et al. Estrogen receptor α signaling regulates breast tumor-initiating cells by down-regulating miR-140 which targets the transcription factor SOX2[J]. J Biol Chem,2012, 287(49): 41514-41522.
[15] Yoshida A, Kitajima S, Li F, et al. MicroRNA-140 mediates RB tumor suppressor function to control stem cell-like activity through interleukin-6[J]. Oncotarget, 2017, 8(8): 13872-13885.
[16] Korkaya H, Kim GI, Davis A, et al. Activation of an IL6 inflammatory loop mediates trastuzumab resistance in HER2+breast cancer by expanding the cancer stem cell population[J].Mol Cell, 2012, 47(4): 570-584.
[17] Barcellos-Hoff MH, Lyden D, Wang TC. The evolution of the cancer niche during multistage carcinogenesis[J]. Nat Rev Cancer, 2013, 13(7): 511-518.
[18] Gernapudi R, Yao Y, Zhang Y, et al. Targeting exosomes from preadipocytes inhibits preadipocyte to cancer stem cell signaling in early-stage breast cancer[J]. Breast Cancer Res Treat, 2015,150(3): 685-695.
[19] Chang JT, Wang F, Chapin W, et al. Identification of MicroRNAs as breast cancer prognosis markers through the cancer genome atlas[J]. PLoS One, 2016, 11(12): e0168284.
[20] Block TM, Mehta AS, Fimmel CJ, et al. Molecular viral oncology of hepatocellular carcinoma[J]. Oncogene, 2003, 22(33): 5093-5107.
[21] Luedde T, Schwabe RF. NF-κB in the liver--linking injury,fibrosis and hepatocellular carcinoma[J]. Nat Rev Gastroenterol Hepatol, 2011, 8(2): 108-118.
[22] Takata A, Otsuka M, Kojima K, et al. MicroRNA-22 and microRNA-140 suppress NF-κB activity by regulating the expression of NF-κB coactivators[J]. Biochem Biophys Res Commun, 2011, 411(4): 826-831.
[23] Takata A, Otsuka M, Yoshikawa T, et al. MicroRNA-140 acts as a liver tumor suppressor by controlling NF-κB activity by directly targeting DNA methyltransferase 1 (Dnmt1) expression[J].Hepatology, 2013, 57(1): 162-170.
[24] Takata A, Otsuka M, Yoshikawa T, et al. A miRNA machinery component DDX20 controls NF-κB via microRNA-140 function[J]. Biochem Biophys Res Commun, 2012, 420(3):564-569.
[25] Yang H, Fang F, Chang R, et al. MicroRNA-140-5p suppresses tumor growth and metastasis by targeting transforming growth factor β receptor 1 and fibroblast growth factor 9 in hepatocellular carcinoma[J]. Hepatology, 2013, 58(1): 205-217.
[26] Yan X, Zhu Z, Xu S, et al. MicroRNA-140-5p inhibits hepatocellular carcinoma by directly targeting the unique isomerase Pin1 to block multiple cancer-driving pathways[J]. Sci Rep, 2017, 7: 45915.
[27] Lv J, Fan HX, Zhao XP, et al. Long non-coding RNA Unigene56159 promotes epithelial-mesenchymal transition by acting as a ceRNA of miR-140-5p in hepatocellular carcinoma cells[J]. Cancer Lett, 2016, 382(2): 166-175.
[28] Yu J, Zhang W, Tang H, et al. Septin 2 accelerates the progression of biliary tract cancer and is negatively regulated by mir-140-5p[J]. Gene, 2016, 589(1): 20-26.
[29] Yanaihara N, Caplen N, Bowman E, et al. Unique microRNA molecular profiles in lung cancer diagnosis and prognosis[J].Cancer Cell, 2006, 9(3): 189-198.
[30] Tan X, Qin W, Zhang L, et al. A 5-microRNA signature for lung squamous cell carcinoma diagnosis and hsa-miR-31 for prognosis[J]. Clin Cancer Res, 2011, 17(21): 6802-6811.
[31] Kong XM, Zhang GH, Huo YK, et al. MicroRNA-140-3p inhibits proliferation, migration and invasion of lung cancer cells by targeting ATP6AP2[J]. Int J Clin Exp Pathol, 2015, 8(10):12845-12852.
[32] Dong W, Yao C, Teng X, et al. MiR-140-3p suppressed cell growth and invasion by downregulating the expression of ATP8A1 in non-small cell lung cancer[J]. Tumour Biol, 2016,37(3): 2973-2985.
[33] Yuan Y, Shen Y, Xue L, et al. miR-140 suppresses tumor growth and metastasis of non-small cell lung cancer by targeting insulinlike growth factor 1 receptor[J]. PLoS One, 2013, 8(9): e73604.[34] Zhang YX, Yan YF, Liu YM, et al. Smad3-related miRNAs regulated oncogenic TRIB2 promoter activity to effectively suppress lung adenocarcinoma growth[J]. Cell Death Dis, 2016,7(12): e2528.
[35] Li W, He F. Monocyte to macrophage differentiation-associated(MMD) targeted by miR-140-5p regulates tumor growth in nonsmall cell lung cancer[J]. Biochem Biophys Res Commun, 2014,450(1): 844-850.
[36] Izzotti A, Calin GA, Arrigo P, et al. Downregulation of microRNA expression in the lungs of rats exposed to cigarette smoke[J]. FASEB J, 2009, 23(3): 806-812.
[37] Li W, Jiang G, Zhou J, et al. Down-regulation of miR-140 induces EMT and promotes invasion by targeting Slug in esophageal cancer[J]. Cell Physiol Biochem, 2014, 34(5): 1466-1476.
[38] Liang S, Gong X, Zhang G, et al. MicroRNA-140 regulates cell growth and invasion in pancreatic duct adenocarcinoma by targeting iASPP[J]. Acta Biochim Biophys Sin (Shanghai), 2016,48(2): 174-181.
[39] Zou J, Xu Y. MicroRNA-140 Inhibits cell proliferation in gastric cancer cell line HGC-27 by suppressing SOX4[J]. Med Sci Monit, 2016, 22: 2243-2252.
[40] Piepoli A, Tavano F, Copetti M, et al. Mirna expression profiles identify drivers in colorectal and pancreatic cancers[J]. PLoS One, 2012, 7(3): e33663.
[41] Zhang W, Zou C, Pan L, et al. MicroRNA-140-5p inhibits the progression of colorectal cancer by targeting VEGFA[J]. Cell Physiol Biochem, 2015, 37(3): 1123-1133.
[42] Yu L, Lu Y, Han X, et al. microRNA-140-5p inhibits colorectal cancer invasion and metastasis by targeting ADAMTS5 and IGFBP5[J]. Stem Cell Res Ther, 2016, 7(1): 180.
[43] Zhao WY, Zou JR, Wang B, et al. microRNA-140 suppresses the migration and invasion of colorectal cancer cells through targeting Smad3[J]. Chin J Oncol, 2014, 36(10): 739-745. [趙文月, 鄒佳芮, 王波, 等. miR140通過下調Smad3表達抑制結腸癌細胞遷移和侵襲能力[J]. 中華腫瘤雜志, 2014, 36(10):739-745.]
[44] Zhai H, Fesler A, Ba Y, et al. Inhibition of colorectal cancer stem cell survival and invasive potential by hsa-miR-140-5p mediated suppression of Smad2 and autophagy[J]. Oncotarget, 2015,6(23): 19735-19746.
[45] Song B, Wang Y, Xi Y, et al. Mechanism of chemoresistance mediated by miR-140 in human osteosarcoma and colon cancer cells[J]. Oncogene, 2009, 28(46): 4065-4074.
[46] Xiao Q, Huang L, Zhang Z, et al. Overexpression of miR-140 inhibits proliferation of osteosarcoma cells via suppression of histone deacetylase 4[J]. Oncol Res, 2017, 25(2): 267-275.
[47] Meng Y, Gao R, Ma J, et al. MicroRNA-140-5p regulates osteosarcoma chemoresistance by targeting HMGN5 and autophagy[J]. Sci Rep, 2017, 7(1): 416.
[48] Pan ST, Li ZL, He ZX, et al. Molecular mechanisms for tumour resistance to chemotherapy[J]. Clin Exp Pharmacol Physiol,2016, 43(8): 723-737.
[49] Sand M, Skrygan M, Sand D, et al. Expression of microRNAs in basal cell carcinoma[J]. Br J Dermatol, 2012, 167(4): 847-855.
[50] Jing P, Sa N, Liu X, et al. MicroR-140-5p suppresses tumor cell migration and invasion by targeting ADAM10-mediated Notch1 signaling pathway in hypopharyngeal squamous cell carcinoma[J]. Exp Mol Pathol, 2016, 100(1): 132-138.
[51] Kai Y, Peng W, Ling W, et al. Reciprocal effects between microRNA-140-5p and ADAM10 suppress migration and invasion of human tongue cancer cells[J]. Biochem Biophys Res Commun, 2014, 448(3): 308-314.
[52] Liu X, Wang S, Yuan A, et al. MicroRNA-140 represses glioma growth and metastasis by directly targeting ADAM9[J]. Oncol Rep, 2016, 36(4): 2329-2338.
[53] Matouk IJ, DeGroot N, Mezan S, et al. The H19 non-coding RNA is essential for human tumor growth[J]. PLoS One, 2007,2(9): e845.
[54] Zhao H, Peng R, Liu Q, et al. The lncRNA H19 interacts with miR-140 to modulate glioma growth by targeting iASPP[J].Arch Biochem Biophys, 2016, 610: 1-7.
[55] Reddemann K, Gola D, Schillert A, et al. Dysregulation of microRNAs in angioimmunoblastic T-cell lymphoma[J].Anticancer Res, 2015, 35(4): 2055-2061.
[56] Correia NC, Mel?o A, Póvoa V, et al. microRNAs regulate TAL1 expression in T-cell acute lymphoblastic leukemia[J].Oncotarget, 2016, 7(7): 8268-8281.
[57] Su Y, Xiong J, Hu J, et al. MicroRNA-140-5p targets insulin like growth factor 2 mRNA binding protein 1 (IGF2BP1) to suppress cervical cancer growth and metastasis[J]. Oncotarget, 2016,7(42): 68397-68411.
[58] Iorio MV, Visone R, Di Leva G, et al. MicroRNA signatures in human ovarian cancer[J]. Cancer Res, 2007, 67(18): 8699-8707.
[59] Miles GD, Seiler M, Rodriguez L, et al. Identifying microRNA/mRNA dysregulations in ovarian cancer[J]. BMC Res Notes,2012, 5: 164.
[60] Lan H, Chen W, He G, et al. miR-140-5p inhibits ovarian cancer growth partially by repression of PDGFRA[J]. Biomed Pharmacother, 2015, 75: 117-122.
[61] Ingelmo-Torres M, Lozano JJ, Izquierdo L, et al. Urinary cell microRNA-based prognostic classifier for non-muscle invasive bladder cancer[J]. Oncotarget, 2017, 8(11): 18238-18247.
[62] Wang M, Du M, Ma L, et al. A functional variant in TP63 at 3q28 associated with bladder cancer risk by creating an miR-140-5p binding site[J]. Int J Cancer, 2016, 139(1): 65-74.
[63] Zou MX, Huang W, Wang XB, et al. Identification of miR-140-3p as a marker associated with poor prognosis in spinal chordoma[J]. Int J Clin Exp Pathol, 2014, 7(8): 4877-4885.
[64] Zhang X, Cairns M, Rose B, et al. Alterations in miRNA processing and expression in pleomorphic adenomas of the salivary gland[J]. Int J Cancer, 2009, 124(12): 2855-2863.
猜你喜歡
保健醫苑(2023年2期)2023-03-15 09:03:04
中國臨床醫學影像雜志(2022年2期)2022-05-25 13:24:34
天津醫科大學學報(2019年3期)2019-08-13 06:53:08
西南國防醫藥(2016年7期)2016-12-01 06:01:15
中國衛生標準管理(2015年1期)2016-01-14 03:41:26
腫瘤預防與治療(2015年1期)2015-09-26 07:26:20
醫學研究雜志(2015年12期)2015-06-10 06:57:46
中國當代醫藥(2015年16期)2015-03-01 02:03:11
鄭州大學學報(醫學版)(2015年1期)2015-02-27 14:50:26
癌變·畸變·突變(2015年4期)2015-02-27 06:15:25
