蒙古高原種子植物屬的區系地理分析
2018-07-28 08:18:28,,,*
草地學報 2018年2期
關鍵詞:植物
, , , *
(1. 內蒙古自治區林業監測規劃院, 內蒙古 呼和浩特 010020; 2. 內蒙古農業大學草原與資源環境學院, 內蒙古 呼和浩特 010019)
蒙古高原地處亞洲大陸腹地干旱與半干旱地區(87°40′~122°15′ E,37°46′~53°08′ N)。南以陰山、賀蘭山、祁連山為界,北至唐努山、薩彥嶺、肯特山以及雅布洛諾夫山脈一帶,東起大興安嶺,西部為薩彥嶺和阿爾泰山脈所環繞[1]。蒙古高原大致包括蒙古國全部地區,俄羅斯聯邦西伯利亞部分地區,以及中國內蒙古自治區、甘肅和新疆維吾爾自治區的部分地區,廣義的蒙古高原還包括陰山以南被黃河包圍的鄂爾多斯高原。
蒙古高原宏觀地貌主要為山地和高平原,平均海拔1580m,整體地勢西高東低[1]。蒙古高原周邊為中、高山地所環繞,遠離海洋,阻擋了來自遠海和遠洋吹來的濕潤氣流,成為一個強烈的大陸性氣候區[2]。氣候特點為冬季漫長寒冷,夏季短暫炎熱,降水稀少,且分布極不均勻,除高原東部、東南部以及北部等少數地區外,絕大部分地區年降水均少于400mm[1]。在山地半濕潤氣候帶形成了森林、草甸和草原的植被組合,在廣大的半干旱氣候帶,發育形成了草原,在干旱地帶發育著荒漠草原植被,在極干旱氣候帶是荒漠植被[3]。
1 材料與方法
2008-2010年間,野外調查以蒙古高原的重要地區為原點進行多方位呈輻射狀逐段調查,根據野外考察線路設置植物樣地,對不同生境不同植被類型的區域進行植物種類調查,并采集制作標本[4]。室內數據處理結合內蒙古、甘肅和新疆地區各大學的植物標本館標本,并參考相關資料[5-17]準確鑒定出物種名稱,系統整理出研究區種子植物名錄,同時運用植物區系地理學基本原理和方法,通過統計、排序以及比較分析的方法,對研究區種子植物區系進行了系統的研究和詳細的論述。
2 結果與分析
2.1 植物種類組成
本區種子植物共有4 280種(含種下分類單位,不包括栽培種),隸屬于761屬115科。其中裸子植物3科、8屬、32種;被子植物112科、753屬、4 248種(雙子葉植物92科、610屬、3 333種;單子葉植物20科、143屬、915種),占區域種子植物總種數的99.3%,在數量上占絕對優勢。從各類群所含的科、屬、種數占世界區系相應類群的比例(表1)可見,蒙古高原種子植物區系相對貧乏,所含種數僅占世界種子植物種數的1.9%(遠低于中國所占比例12.1%),且以草原植被和荒漠植被為主,說明蒙古高原自然環境較為嚴酷,這與蒙古高原所處的半干旱和干旱地區的地理位置相適宜。
2.2 屬的組成分析
對屬的大小統計表明(表2),本區域單型屬312屬,含312種,占總屬數的41.0%,總種數的7.3%,其中,單種屬80屬,占區域總屬數的10.5%;少型屬(2~9種)共338屬,含1 317種,占總屬數的44.6%,占總種數的30.8%;中型屬(10~19種)共71屬,含911種,占總屬數的9.3%,占總種數的21.3%;較大屬(20~29種)共22屬,含519種,占總屬數的2.9%,占總種數的12.1%;大型屬(≥30種)只有18屬,共計1 221種,僅占本區域總屬數的2.4%,卻占總種數的28.5%。這說明本區域屬的分化較大,中型屬、小型屬非常豐富,大型屬較少,卻包含較多的種,比較發達,在植被構成中占據著十分重要的地位。從種屬比例看,大型屬、較大屬和中型屬的種屬比例分別是67.8、23.6和12.8,明顯高于區域植物種屬比的平均值5.6,說明大型屬、較大屬和中型屬在種水平上豐富度較高,具有一定的優勢。
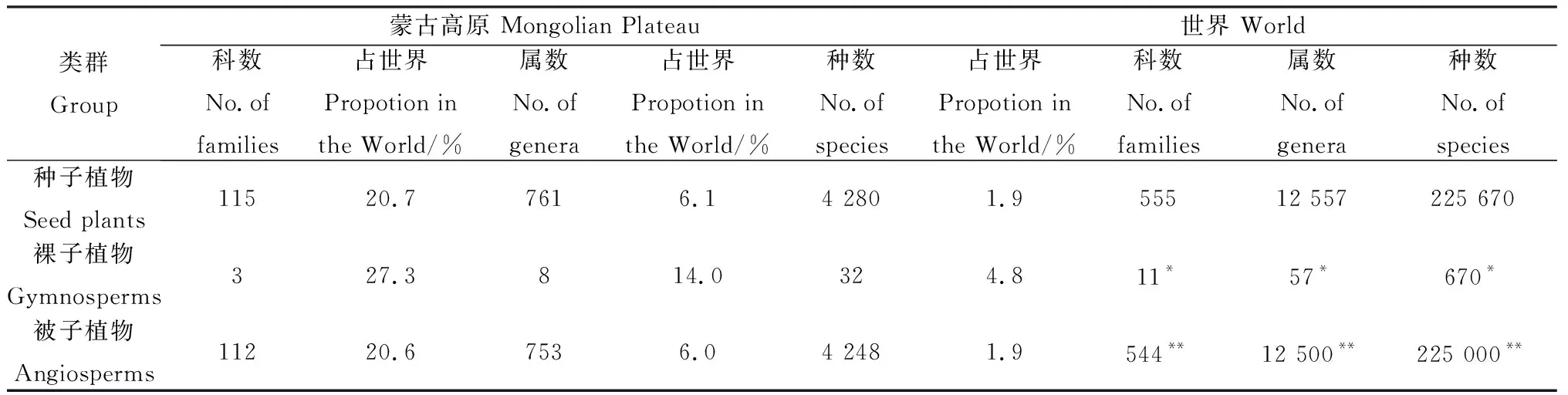
表1 蒙古高原植物大類群統計Table 1 Statistic of communities in Mongolian Plateau
注:*裸子植物的世界資料來源于J.C.Willis(1973)[18].**被子植物世界資料來源于R.Good(1974)[19]
Note:*indicates the Gymnosperms data of the World from J.C.Willis(1973)[18].**indicates the Angiosperms data of the World from R.Good(1974)[19]
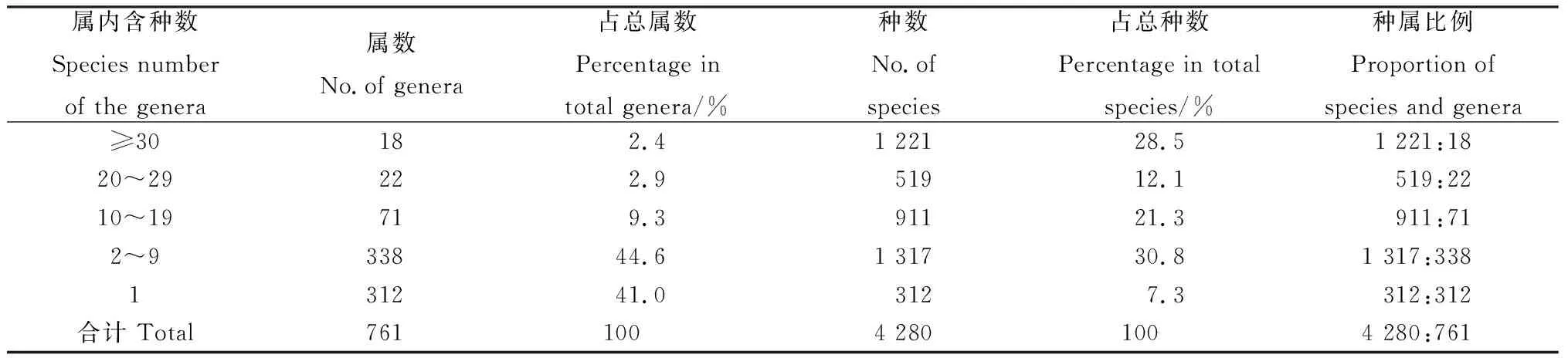
表2 蒙古高原種子植物屬的統計Table 2 Genera of spermatophyte in Mongolian Plateau
2.3 屬的地理成分分析
參考吳征鎰先生對中國種子植物屬的分布區類型的劃分,將蒙古高原種子植物761屬劃分成15個分布區類型和22個變型(表3)[20-21]。
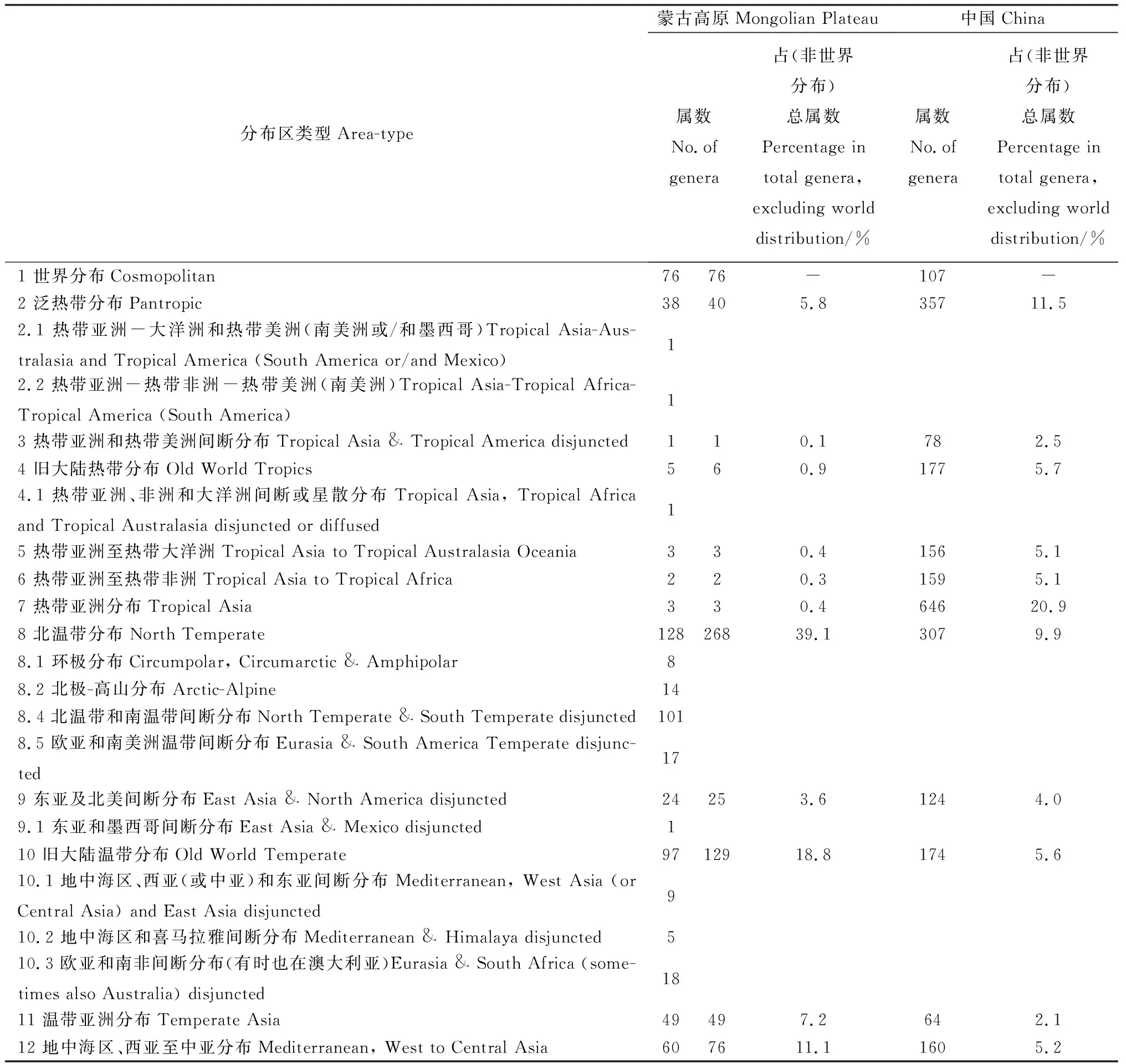
表3 蒙古高原種子植物屬的分布區類型Table 3 The distribution types of spermatophyte genera in Mongolian Plateau
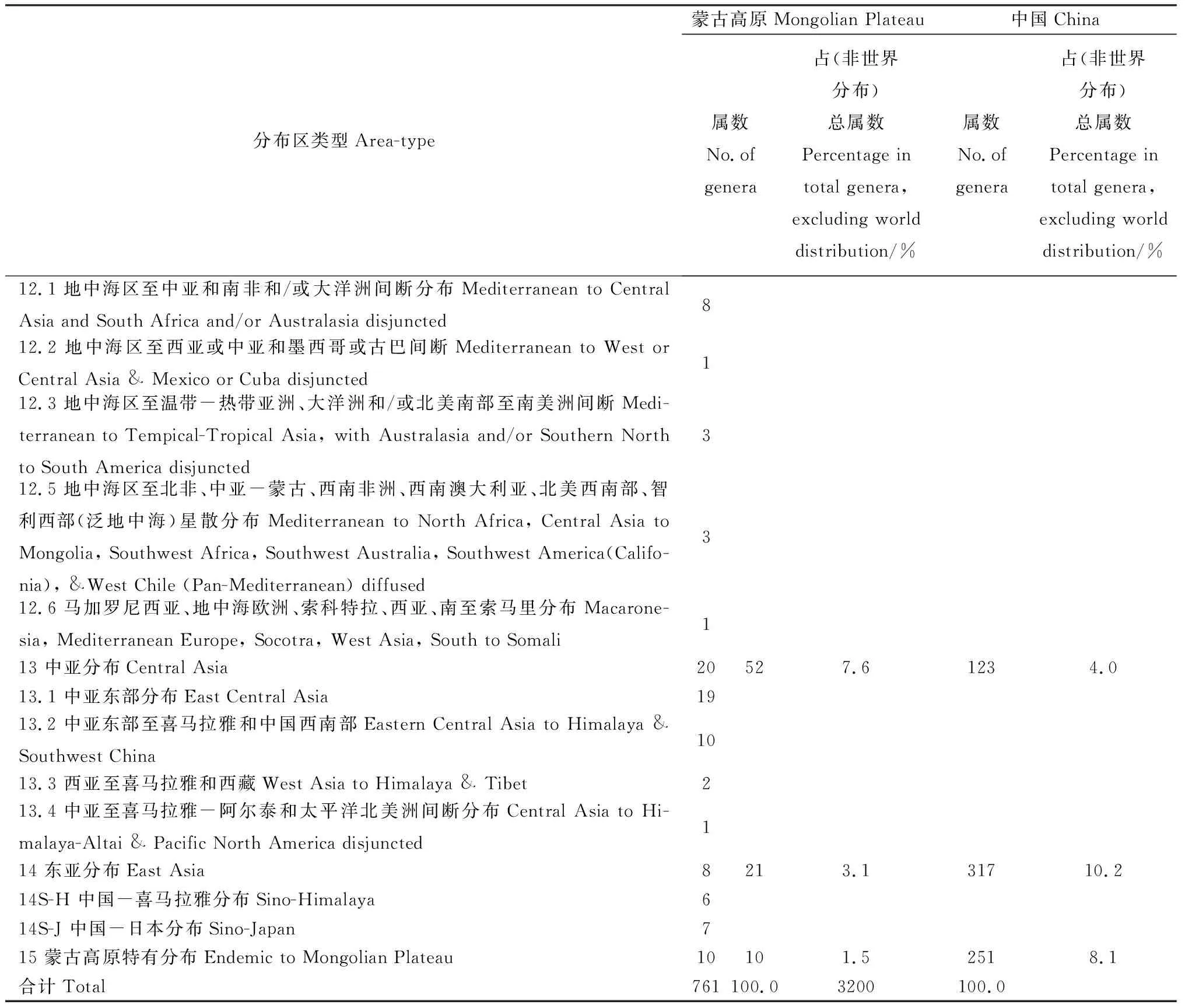
續表3
2.3.1世界分布屬 本類型蒙古高原有76屬,占總屬數的10.0%,遠高于我國該類型所占的比例(3.5%)[22]。從生態型來看,該類型的屬是以溫帶起源的喜濕或中生草本植物為主[23],且廣布于蒙古高原。如在各地草甸分布的有千里光屬(Senecio)、龍膽屬(Gentiana)、老鸛草屬(Geranium)、酸模屬(Rumex)、黃芩屬(Scutellaria)、遠志屬(Polygala)、毛茛屬(Ranunculus)和苔草屬(Carex)等;在各地湖沼及河岸淺水中分布的有莎草屬(Cyperus)、藨草屬(Scirpus)、燈心草屬(Juncus)、香蒲屬(Typha)、浮萍屬(Lemna)、眼子菜屬(Potamogeton)、貍藻屬(Utricularia)、茨藻屬(Najas)等。區域內還有少數含木本植物的屬,如槐屬(Sophora)、懸鉤子屬(Rubus)、鼠李屬(Rhamnus)、蒿屬(Artemisia)、衛矛屬(Euonymus)和豬毛菜屬(Salsola),是灌叢的主要組成成分,唯有槐屬和衛矛屬可達到喬木的高度。其中不乏有少數屬,如豬毛菜屬、旋花屬(Convolvulus)、蒿屬、堿蓬屬(Suaeda)、鹽角草屬(Salicornia)、黃芪屬(Astragalus)、和補血草屬(Limonium)等,主要產于干旱地區或鹽漬化的生境上,是荒漠植被或鹽生草甸的主要植被。
2.3.2泛熱帶分布 本類型蒙古高原有40屬,其中熱帶亞洲-大洋洲和熱帶美洲分布變型1屬,熱帶亞洲-熱帶非洲-熱帶美洲分布變型1屬,共占蒙古高原總屬數的5.8%,低于我國該類型所占的比例(11.5%)[22],這些屬中沒有僅限于熱帶地區分布的屬。該類型的木本屬只有樸屬(Celtis)、一葉萩屬(Securinega)、牡荊屬(Vitex)、棗屬(Zizyphus)和醉魚草屬(Buddleja),它們主要分布于蒙古高原溫帶地區的森林、山地灌叢及溝谷,幾乎不進入寒溫帶地區。該類型的草本植物屬多為單種屬或寡種屬,可見這些泛熱帶屬在蒙古高原這一遼闊的地區種系發育得并不好。
2.3.3熱帶亞洲和熱帶美洲間斷分布 本類型蒙古高原僅有1屬,為地榆屬(Sanguisorba),常生于草原、草甸、山坡草地、灌叢中或疏林下,占蒙古高原總屬數的0.1%,明顯低于我國該類型所占的比例(2.5%)[23]。可見,蒙古高原植物區系與熱帶亞洲和熱帶美洲植物區系曾有共同的淵源,這是由于熱帶美洲(南美洲)本來位于古南大陸的西部,最早于侏羅紀末期和非洲開始分離,至白堊紀末期與之完全分離,之后熱帶亞洲和熱帶美洲的植物又都經過漫長的演化和發展歷程,使種群逐步擴散到溫帶地區,造就了蒙古高原種子植物區系與熱帶亞洲和熱帶美洲植物區系的微弱聯系[23]。
2.3.4舊大陸熱帶分布 本類型蒙古高原有6屬,其中熱帶亞洲、非洲和大洋洲間斷或星散分布變型1屬,共占總屬數的0.9%,遠低于我國同類型屬數的比例(5.7%)[22]。其中,天門冬屬(Asparagus)、百蕊草屬(Thesium)廣泛分布于蒙古高原各類生境中。槲寄生屬(Viscum)、虎舌蘭屬(Epipogium)主要分布在蒙古高原的森林地區。牛鞭草屬(Hemarthria)、藎草屬(Arthraxon)常見于蒙古高原的各類草甸中。
2.3.5熱帶亞洲至熱帶大洋洲 本類型蒙古高原有3屬,占總屬數的0.4%,明顯低于我國同類型屬所占的比例(5.1%)[22]。其中臭椿屬(Ailanthus)是該類型中唯一一個作為優勢樹種能夠分布到蒙古高原南部地區的森林植被中[23]。大豆屬(Glycine)和犁頭尖屬(Typhonium)作為伴生種,常分布于蒙古高原地區的河岸、灌叢、山地或田野。
2.3.6熱帶亞洲至熱帶非洲 本類型蒙古高原有2屬,分別為杠柳屬(Periploca)和芒屬(Miscanthus),其中的2個變型(華南、西南到印度和熱帶非洲間斷分布和熱帶亞洲和東非間斷分布)均缺,占總屬數的0.3%,明顯低于我國同類屬所占的比例(5.1%)[22],這與蒙古高原所處的氣候帶有關。
2.3.7熱帶亞洲分布 本類型蒙古高原僅有3屬,分別為赤爮屬(Thladiantha)、苦荬菜屬(Ixeris)和小苦荬屬(Ixeridium),占總屬數的0.4%,遠低于全國同類型屬數的比例(20.9%)[22]。它們在蒙古高原常出現在林緣、山坡草地、河邊、溝谷甚至是沙地,這一類型在蒙古高原植物區系中所起的作用很小。
2.3.8北溫帶分布 本類型蒙古高原有268屬,其中環極分布變型8屬、北極-高山分布變型14屬、北溫帶和南溫帶間斷分布變型101屬、歐亞和南美洲溫帶間斷分布變型17屬,共占總屬數的39.1%,明顯高于全國同類屬所占的比例(9.9%)[22]。這一類型在屬和種的數量上均占據著最高的比例,在蒙古高原植物區系組成上占有極為重要的作用。它們的水分生態型多為中生的木本或草本植物。前者如裸子植物的冷杉屬(Abies)、云杉屬(Picea)、落葉松屬(Larix)、松屬(Pinus)、刺柏屬(Juniperus)、圓柏屬(Sabina)均為山地針葉林植被的重要組成成分,尤以落葉松屬和松屬最為突出,可作為蒙古高原山地植物群落的優勢種和建群種,也呈斑塊狀少量進入森林草原地帶;被子植物的楊屬(Populus)、樺木屬(Betula)、櫟屬(Quercus)、榆屬(Ulmus)、槭樹屬(Acer)、椴樹屬(Tilia)、接骨木屬(Sambucus)、花楸屬(Sorbus)、桑屬(Morus)是構成蒙古高原落葉闊葉林和針闊葉混交林的主要成分,其中多數種是區域森林植被的建群種或優勢種。還有一些灌木屬如山梅花屬(Philadelphus)、榛屬(Corylus)、越橘屬(Vaccinium)、薔薇屬(Rosa)、杜鵑花屬(Rhododendron)、繡線菊屬(Spiraea)、茶藨屬(Ribes)、莢蒾屬(Viburnum)、忍冬屬(Lonicera)、胡頹子屬(Elaeagnus)是構成植物群落下木層的主要成分或山地落葉灌叢的主要建群植物。溪谷、河岸兩側及河流下游的河床上常分布有柳屬(Salix)植物,部分地區還可見梾木屬(Swida)、榿木屬(Alnus)、繡線菊屬和薔薇屬植物。此外,該類型的草本屬更是發達。雙子葉植物如龍芽草屬(Agrimonia)、蚊子草屬(Filipendula)、舞鶴草屬(Maianthemum)、委陵菜屬(Potentilla)、迷果芹屬(Sphallerocarpus)、景天屬(Sedum)等及單子葉植物如鈴蘭屬(Convallaria)、舞鶴草屬(Maianthemum)、黃精屬(Polygonatum)等均為山地林下或山地草甸植物的主要代表;雙子葉植物遠志屬(Polygala)、棘豆屬(Oxytropis)和單子葉植物針茅屬(Stipa)、堿茅屬(Puccinellia)、披堿草屬(Elymus)、賴草屬(Leymus)和拂子茅屬(Calamagrostis)等旱生植被,是蒙古高原典型草原和山坡草地的優勢成分。巖高蘭屬(Empetrum)、杜香屬(Ledum)、北極花屬(Linnaea)、毛蒿豆屬(Oxycoccus)、松毛翠屬(Phyllodoce)、冰沼草屬(Scheuchzeria)、熊果屬(Arctostaphylos)、北極果屬(Arctous)、肉果薺屬(Braya)、單側花屬(Orthilia)、裂稃茅屬(Schizachne)、巖須屬(Cassiope)和山崳菜屬(Eutrema)等,常常是山地凍原、高山植被或寒溫帶針葉林下的代表植物或重要成分。
2.3.9東亞和北美間斷分布 本類型蒙古高原有25屬,包括東亞和墨西哥間斷分布變型1屬,占總屬數的3.6%,與全國同類屬所占的比例(4.0%)[22]相近。含木本的屬僅有珍珠梅屬(Sorbaria)、羅布麻屬(Apocynum)、八仙花屬(Hydrangea)、胡枝子屬(Lespedeza)和五味子屬(Schisandra)5屬。除羅布麻屬會分布到較為干旱的鹽堿荒地、沙漠邊緣、戈壁荒灘及河流兩岸、沖擊平原、河湖周圍外,其余4屬主要是蒙古高原森林中的重要成分,但胡枝子屬的草本種也常分布到草原中。這一間斷分布的成分或在聯合古陸解體之前的白堊紀就已存在于北美及東亞,或在聯合古陸分離后繼續演化發展而來,并呈現出現今的間斷分布格局[24]。
2.3.10舊大陸溫帶分布 本類型蒙古高原有129屬,包括地中海區、西亞(或中亞)和東亞間斷分布變型9屬、地中海區和喜馬拉雅間斷分布變型5屬、歐亞和南非間斷分布變型18屬,共占總屬數的18.8%,遠高于全國同類屬所占的比例(5.6%)[22],這一類型及變型在蒙古高原植物區系中占的比例僅次于北溫帶分布型及其變型。從生活型上看,木本屬僅有沙棘屬(Hippophae)、檉柳屬(Tamarix)、水柏枝屬(Myricaria)、梨屬(Pyrus)、栒子屬(Cotoneaster)、桃屬(Amygdalus)、木蓼屬(Atraphaxis)、鮮卑花屬(Sibiraea)和丁香屬(Syringa)9屬,占該類型屬數的7.0%,其中檉柳屬、水柏枝屬和木蓼屬是蒙古高原荒漠地區極為廣布的重要的優勢屬和建群屬,其它木本屬均為森林的重要組成成分。中生或旱中生的草本屬占大多數,如芨芨草屬(Achnatherum)、冰草屬(Agropyron)、隱子草屬(Cleistogenes)常成為典型草原地區的優勢或建群植物,其他草本屬如沙參屬(Adenophora)、荊芥屬(Nepeta)、聚合草屬(Symphytum)、白鮮屬(Dictamnus)、滇紫草屬(Onosma)、菊屬(Dendranthema)、藍盆花屬(Scabiosa)、乳菀屬(Galatella)、蓍屬(Achillea)、萱草屬(Hemerocallis)、沼蘭屬(Microstylis)、芯芭屬(Cymbaria)、敗醬屬(Patrinia)、石竹屬(Dianthus)、麻花頭屬(Serratula)、樟味藜屬(Camphorosma)、蠟菊屬(Helichrysum)等,為林下草本層、高山灌叢、山地草甸、草甸、典型草原、荒漠草原的重要組成成分。
2.3.11溫帶亞洲分布 本類型蒙古高原有49屬,占總屬數的7.2%,遠高于全國同類屬所占的比例(2.1%)[22]。含木本的屬僅有錦雞兒屬(Caragana)、杏屬(Armeniaca)和亞菊屬(Ajania)3屬,其余均為草本植物,主要有線葉菊屬(Filifolium)、馬蘭屬(Kalimeris)、狗娃花屬(Heteropappus)、米口袋屬(Gueldenstaedtia)、防風屬(Sapoghnikovia)、軸藜屬(Axyris)、裂葉荊芥屬(Schizonepeta)、狼毒屬(Stellera)、細柄茅屬(Ptilagrostis)等。其中,錦雞兒屬、亞菊屬、線葉菊屬、細柄茅屬等常是草原中的建群植物。
2.3.12地中海區、西亞至中亞分布 本類型蒙古高原有76屬,包括地中海區至中亞和南非和/或大洋洲間斷分布變型8屬、地中海區至西亞或中亞和墨西哥或古巴間斷分布變型1屬、地中海區至溫帶-熱帶亞洲、大洋洲和/或北美南部至南美洲間斷分布變型3屬、地中海區至北非、中亞-蒙古、西南非洲、西南澳大利亞、北美西南部、智利西部(泛地中海)星散分布變型3屬、馬加羅尼西亞、地中海歐洲、索科特拉、西亞、南至索馬里分布變型1屬,共占總屬數的11.1%,遠高于全國同類屬所占的比例(5.2%)[22],這一類型及變型在蒙古高原植物區系中所占的比例居第三位。這一成分中單型屬和少型屬很豐富。從生態類型上看,多是旱生和鹽生的種類,它們是蒙古高原荒漠草原、荒漠以及鹽沼、堿地的隱域植被的重要組成者,在蒙古高原植物區系中起著十分重要的作用,常常成為群落的優勢植物或建群植物,甚至形成單優勢群落。如紅砂屬(Reaumuria)、半日花屬(Helianthemum)、假木賊屬(Anabasis)、梭梭屬(Haloxylon)、駝絨藜屬(Ceratoides)、小蓬屬(Nanophyton)、鹽爪爪屬(Kalidium)、單刺蓬屬(Cornulaca)、裸果木屬(Gymnocarpos)、沙拐棗屬(Calligonum)、霸王屬(Zygophyllum)、白刺屬(Nitraria)、駱駝蓬屬(Peganum)、苓菊屬(Jurinea)、補血草屬(Limonium)、花花柴屬(Karelinia)及禾本科的獐茅屬(Aeluropus)等。還有一些短命、類短命植物及珍貴的肉質寄生植物,如鎖陽(Cynomorium)及肉蓯蓉屬(Cistanche)等。
2.3.13中亞分布 本類型蒙古高原有52屬,包括中亞東部分布變型19屬、中亞東部至喜馬拉雅和中國西南部變型10屬、西亞至喜馬拉雅和西藏分布變型2屬、中亞至喜馬拉雅-阿爾泰和太平洋北美洲間斷分布變型1屬,共占總屬數的7.6%,明顯高于全國同類屬數所占的比例(4.0%)[22]。這一類型單型屬和少型屬更為豐富,它們多是旱生或耐旱的草本或木本,多生于山前荒漠、礫石戈壁、丘間地。典型的中亞分布屬有矮刺蘇屬(Chamaesphacos)、翅膜菊屬(Alfredia)、短舌菊屬(Brachanthemum)、寒蓬屬(Psychrogeton)、琉苞菊屬(Hyalea)、絨藜屬(Londesia)、四齒芥屬(Tetracme)、兔唇花屬(Lagochilus)、蛛絲蓬屬(Micropeplis)、紫菀木屬(Asterothamnus)等;分布于亞洲中部的戈壁藜屬(Iljinia)、革苞菊屬(Tugarinovia)、合頭草屬(Sympegma)、沙蓬屬(Agriophyllum)、沙冬青屬(Ammopiptanthus)、櫛葉蒿屬(Neopallasia)、沙芥屬(Pugionium)等屬均是比較古老的荒漠植物。扁芒菊屬(Waldheimia)、高河菜屬(Megacarpaea)、角蒿屬(Incarvillea)、長柱琉璃草屬(Lindelfia)、擬耬斗菜屬(Paraquilegia)等,主要是蒙古高原高山上衍生的比較年輕的成分。這類成分均為地中海區、西亞至中亞分布成分在亞洲地區經過強烈旱化的衍生物,由于天山山脈和青藏高原的抬升,產生山地特有現象,從北溫帶和舊大陸溫帶廣布的大屬中進一步分化出一些新屬,如女蒿屬(Hippolytia)和沙鞭屬(Psammochloa)分別是從菊蒿屬(Tanacetum)和芨芨草屬(Achnatherum)分化衍生而來。個別的如角蒿屬(Incarvillea)則遠達四川和貴州,可以說這些高原高山成分是鄰近亞熱帶森林區系經過高山旱化和特化的結果[23]。
2.3.14東亞分布屬 本類型蒙古高原有21屬,包括中國-喜馬拉雅分布變型6屬、中國-日本分布變型7屬,共占總屬數的3.1%,明顯低于全國同類屬數的比例(10.2%)。本類型以森林成分為主,如側柏屬(Platycladus)在蒙古高原南部山地往往形成單優勢群落;五加屬(Eleutherococcus)、錦帶花屬(Weigela)、斑種草屬(Bothriospermum)、翅果菊屬(Pterocypsella)、黨參屬(Codonopsis)、裂瓜屬(Schizopepon)、兔兒傘屬(Syneilesis)、翠菊屬(Callistephus)、黃瓜菜屬(Paraixeris)、桔梗屬(Platycodon)、松蒿屬(Phtheirospermum)等成為林下或林緣草甸的常見成分。也有分布在干旱、半干旱區山地的屬,如野丁香屬(Leptodermis)、蕕屬(Caryopteris)、地黃屬(Rehmannia)。還有部分屬是組成山地草甸或河灘草甸的重要成分,如甘松屬(Nardostachys)、微孔草屬(Microula)、肉果草屬(Lancea)等。
2.3.15蒙古高原特有屬 蒙古高原僅有10個特有屬,其中,真特有屬[25]有沙芥屬(Pugionium)、沙鞭屬(Psammochloa)、連蕊芥屬(Synstemon)、綿刺屬(Potaninia)[26]、四合木屬(Tetraena)、革苞菊屬(Tugarinovia)、百花蒿屬(Stilpnolepis)、紊蒿屬(Elachanthemum);準特有屬[25]有膿瘡草屬(Panzeria)和芯芭屬(Cymbaria)。
3 討論
3.1 特有現象
蒙古高原分布的761屬中僅有10個屬是蒙古高原特有屬,只占總屬數的1.5%,遠低于全國特有屬所占的比例(8.1%)[22],特有程度較低。根據各特有屬分布區域的不同,又劃分為7個特有分布變型。如沙芥屬(Pugionium)和沙鞭屬(Psammochloa)為蒙古高原沙地分布的特有屬;連蕊芥屬(Synstemon)為阿拉善荒漠南緣低山丘陵分布的特有屬;綿刺屬(Potaninia)[26]為阿拉善荒漠特有屬;四合木屬(Tetraena)和革苞菊屬(Tugarinovia)為東阿拉善-西鄂爾多斯特有屬;百花蒿屬(Stilpnolepis)為東阿拉善-鄂爾多斯特有屬;紊蒿屬(Elachanthemum)為戈壁蒙古分布的特有屬;膿瘡草屬(Panzeria)和芯芭屬(Cymbaria)[28]則是主要分布在蒙古高原的準特有屬。
10個特有屬在區系上全部都是單種屬或雙種中群,分類上比較孤立,但全部是被子植物中各個所屬科中的最進化類群,在生態上絕大多數是荒漠種和半荒漠種,均為旱生植物,且沙生植物占優勢,它們的生活型以草木植物占絕對優勢。這10個屬在地理分布上集中分布于蒙古高原的阿拉善-鄂爾多斯地區,東阿拉善-西鄂爾多斯是其分析中心,起源上多數是東亞與地中海植物區系的后裔。因此,無論在區系分類上,還是生態生活型上,這些特有屬都是比較年輕的類群[27]。
3.2 區系地理聯系
蒙古高原種子植物區系與世界大部分地區均有一定聯系,尤與北溫帶的關系最為密切,如前文所述,北溫帶成分不僅廣布于蒙古高原各地區,而且大多數北溫帶成分的喬木和灌木能夠形成森林植被和灌叢植被的優勢成分或建群成分,表現了北溫帶區系與蒙古高原區系有著非常密切的歷史淵源。其次是與地中海、西亞至中亞區系之間的聯系,蒙古高原屬于與地中海、西亞和中亞相聯系的屬有137屬,占本區溫帶性質成分的21.7%,這些地中海、西亞和中亞的成分是地中海脫離海侵局面之后,第三紀的熱帶成分、東亞成分、溫帶成分逐步向西北遷移與演變以及適應地中海區氣候旱化發生蛻變和演化而來的,這與地中海孑遺成分有著共同的淵源。再次是與東亞及北美大陸溫帶區系之間的聯系,共有25屬,這些成分可能與第三紀晚期至第四紀氣候變冷之后的溫帶植物南移與演變以及間冰期植物的南北、東西移動有著密切的聯系。最后是與北極的聯系,蒙古高原與北極相聯系的有22屬,這些成分與在受第四紀多次冰期影響的氣候變化條件下的植物進退有著密切關系。關于蒙古高原與熱帶亞洲、熱帶非洲、熱帶大洋洲和熱帶美洲一些地區存在微弱的聯系,蒙古高原的熱帶成分屬的比例為7.2%,提供了蒙古高原植物區系在發展歷史上與熱帶區系相聯系的遺跡和證據。
3.3 起源與演化
蒙古高原的現代種子植物區系是古生物區系的延續和殘遺,蒙古高原大部分地區曾被Tethys海所占據,經歷了始新世中期的喜馬拉雅造山運動后,大陸性氣候愈加強烈[29],至此,許多成分便在干旱的古地中海沿岸或遺跡上發生。如分布于蒙古高原中央戈壁區的麻黃屬(Ephedra)植物-膜果麻黃(Ephedraprzewalskii)在三疊紀時便發源于古地中海沿岸[30]。白堊紀尤其是老第三紀孑遺種或者是古老的或分類上孤立的殘遺植物如綿刺(Potaniniamongolica)、沙冬青(Ammopiptanthusmongolicus)等均是起源于古地中海沿岸或遺跡上[31]。本區分布的古地中海孑遺植物如沙拐棗(Calligonummongolicum)、鹽節木(Haloenemumstrobilaceum)、花花柴(Kareliniacaspia)、甘草(Glycyrrhizauralensis)[32]等無疑是在古地中海沿岸及遺跡上發生的。中亞也是Tethys海遺跡的一部分[33],因而起源于古地中海沿岸或遺跡上的中亞分布類型無疑也是古地中海植物區系的孑遺,或者說是一些植物種在干旱的荒漠環境中蛻變演化出來的植物種類,并不是古老區系的起源中心。
第三紀后半期,隨著喜馬拉雅的上升,蒙古高原脫離了海侵的局面,東亞植物區系的溫帶成分得以向西遷移[34],并在此獨特環境中,物種的形成過程朝向旱生化發展,產生了現代地中海植物區系的基本成分,典型的代表有錦雞兒屬(Caragana)、野丁香屬(Leptodermis)和羅布麻屬[35]。如錦雞兒屬是發生于始新世早期亞洲東部森林成分,后沿古老的“Зысса”山系的山脊向西遷移,隨著喜馬拉雅和其他第三紀褶皺山系的形成,其中,遷向西北的一支向著適應干旱荒漠氣候發展,產生了小葉系(Ser.Microphyllae)較進化的類型[36],其中有些種已經成為適應干旱氣候的荒漠灌木[37]。又如僅局限分布于阿拉善荒漠區的綿刺作為一個蒙古高原古老的殘遺旱生植物區系成分也是由東亞中生植物區系成分的銀露梅(Potentillaglabra)演化而來[38]。
蒙古高原的溫帶成分發源于中國亞熱帶,亞熱帶常綠闊葉林成分向西北遷移,在適應本地區逐漸變旱的氣候,進而蛻變發展形成適應此特殊生境的次生植物種類[33]。如沙冬青是亞洲荒漠地區僅有的常綠闊葉灌木,這應該是亞熱帶常綠闊葉林植物種干旱變型的有效證據[39];廣布于中央戈壁的膜果麻黃也是屬于老第三紀亞熱帶常綠植被干旱變型的殘遺成分[39]。此外,北溫帶成分在蒙古高原大量存在,且多數的木本植物在森林和灌叢植被中,往往形成群落的優勢成分或建群成分,由此可見溫帶成分在本區植物區系中占重要地位,反映了蒙古高原所處的地理位置(主要是緯度和海拔高度)與其氣候條件相適宜的特點。
蒙古高原植物區系還有部分是從本地的荒漠環境中發展起來的,如蒺藜科(Zygophyllaceae)的霸王屬(Zygophyllum)、駱駝蓬屬(Peganum),藜科(Chenopodiaceae)的鹽爪爪屬(Kalidium)、鹽穗木屬(Halostachys)、豬毛菜屬,石竹科(Caryophyllaceae)的刺葉屬(Acanthophyllum),菊科(Compositae)的刺頭菊屬(Cousinia)、粉苞苣屬(Chondrilla),百合科(Liliaceae)的郁金香屬(Tulipa)及蓼科(Polygonaceae)的沙拐棗屬(Calligonum)和木蓼屬(Atraphaxis)等是在干旱的荒漠環境中形成的[39]。蒙古高原特有屬中的新生進化成分亦是如此,因為它們并不分布到蒙古高原以外的地方,它們是蒙古高原植物區系的一部分。
4 結論
蒙古高原處于北溫帶半干旱和干旱的內陸地區,自然環境較為嚴酷,種子植物僅115科761屬4 280種,占世界植物同類群總種數的1.9%,植物區系較貧乏,特有程度較低。蒙古高原所處的地理位置決定了其區域植物具有典型的溫帶性質,其區系地理成分復雜,聯系廣泛,一定程度上亦受到熱帶成分的影響,與熱帶成分具有一定的親緣關系。植物區系優勢現象不顯著,以單型屬和少型屬為主。區域內植被特點是以旱生和超旱生的灌木、小灌木植物占優勢,其中有很多是古老的或分類上孤立的殘遺植物,使得蒙古高原植物區系表現出強烈的旱生性和古老性。從植物區系發生角度分析,蒙古高原植物區系的起源可歸結為古地中海成分、東亞成分、溫帶成分和本地起源。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
