土壤水分對入侵植物節節麥表型可塑性和化感作用的影響
2018-07-28 08:18:36,*,,
草地學報 2018年2期
關鍵詞:植物
, *, ,
(1. 河南科技大學 林學院, 河南 洛陽 471000; 2. 洛陽市園林局, 河南 洛陽471002; 3. 洛陽隋唐城遺址植物園, 河南 洛陽471002)
表型可塑性是植物同一基因型對異質環境產生不同表型的特性,是植物通過改變形態、生理及發育等癥狀適應環境變化的能力體現[1-3]。植物表型可塑性包括形態、生理及生態可塑性3個方面[4]。表型可塑性可使植物擁有更寬的生態幅和更強的耐受性,為植物占據更廣闊的地理范圍及更多樣化的生境,并最終成為廣幅種奠定基礎[4-5]。不同生境中,外來入侵植物往往表現出較強的形態和生理可塑性反應,從而獲取更多的營養和資源,達到提高其競爭力和入侵能力的目的[6-8]。目前,入侵植物表型可塑性的研究多集中于非生物因子,如溫度、光照、水分等方面[9-12]。
節節麥(AegilopstauschiiCoss.),禾本科山羊草屬1年生或越年生植物,起源于東歐、西亞等地,是世界惡性雜草[13]。自1955年,節節麥在河南省新鄉市被葉德嫻等[14]首次發現并采集到標本,如今已成功入侵至我國陜西、山東、河北等地,并呈迅速蔓延的態勢。由于節節麥與小麥親緣關系較近,二者又具有類似的外部形態及生長習性,更重要的是節節麥擁有較強的分蘗、繁殖及適應性等特點,共同導致其成為麥田中最難防除的禾本科惡性雜草之一[14-15]。因嚴重威脅我國糧食生產安全,節節麥已被列入《中華人民共和國進境植物檢疫性有害生物名錄》。土壤水分是決定植物分布的關鍵因子之一,依據入侵植物與入侵地土壤水分間關系的研究,對預測其分布范圍及防控其進一步擴散具有重要意義。至今,有關節節麥表型可塑性的研究尚未有報道。本文通過研究不同土壤含水量對節節麥表型可塑性以及化感作用的影響,探討其生物量分配模式和生態適應策略,為闡明節節麥的入侵能力及入侵機制提供參考依據。
1 材料與方法
1.1 材料
節節麥幼苗于2017年1月采自于河南省新鄉市。白菜(Brassicarapavar.glabra)種子購自洛陽市種子公司。
1.2 方法
1.2.1幼苗生長實驗 試驗所用土壤為褐土,采自于洛陽市洛龍區周邊農田。環刀法測定農田的田間最大持水量(water holding capacity, WHC)。將供試土壤置于100℃烘箱烘干至恒重。試驗設置包括:田間最大持水量的75%~80%(CK,正常水平)、55%~60%中度脅迫(moderate stress,MD)和45%~50%重度脅迫(serious stress,SD)3個水分梯度[16-17],每處理3次重復,同時設置相應的空白盆。選用Φ 25 cm花盆,干土5.50 Kg·盆-1。選取植株大小接近、長勢良好節節麥幼苗移栽至花盆中,10株·盆-1,盆底放置托盤。隨機選取5株,測定其總生物量和葉面積,求其平均值。移栽后澆透水,緩苗約1周后逐漸控水,稱重實現水分脅迫條件。試驗正式開始后,稱重1次·2 d-1,當土壤水分含量低于設置范圍,采用緩慢澆水法補充水分,并記錄每次澆水量。為排除植株生長量造成的水分補充誤差,每隔15 d隨機拔取每盆中1株植株,稱重換算每盆增加質量,用于調整設定盆栽質量。40 d后結束試驗,每處理隨機選取10株進行相關指標測定,利用掃描儀、Photoshop軟件進行葉面積測定[18]。將節節麥植株的根、莖、葉和穗分開并裝紙袋,80℃烘箱中烘干至恒重。
計算各構件生物量比(構件生物量/植株總生物量)及如下參數[19]:葉重分數(leaf mass fraction, LMF)即葉干重/植株地上部分總干重;葉面積比(leaf area ratio, LAR) 即總葉面積/植株總干重;葉根比(leaf area root mass ratio, LARMR) 即葉總面積/根干重;根冠比(root mass /crown mass, R/C)即根干重/植株地上部分總干重;比葉面積(specific leaf area, SLA)即總葉面積/總葉重;平均葉面積(mean leaf area,MLA)即總葉面積/葉片數。此外,還包括:
平均相對生長速率(relative growth rate,RGR)= (lnW2-lnW1)/t;
凈同化速率(net assimilation rate,NAR)= (W2-W1)/(L2-L1) *(lnL2-lnL1)/t;
水分利用效率(water use efficiency,WUE)= 干物質量(g)/耗水量(kg)
其中W1、L1分別表示處理前測定的節節麥植株總生物量(g)及葉面積(cm2)的平均值;W2、L2分別表示處理后單個重復的總生物量和葉面積。t表示處理的天數[20-21]。
表型可塑性指數(phenotypic plasticity index, PPI)=(某一指標在幾種處理中的最大值-最小值)/最大值。PPI的大小表示某一變量對某一指標表型可塑性影響作用的大小[22-23]。
1.2.2生測實驗
1.2.2.1 水浸提液的制備
莖葉和根系浸提液:試驗結束后收集不同處理的節節麥植株,蒸餾水清洗干凈,室內自然晾干。分別剪取莖葉、根系,各稱取100 g,充分研磨后加入1 000 mL蒸餾水,浸泡48 h,2 500 r·min-1離心4 min,取上清液,重復3次,棄沉淀,將上清液定溶至1 000 mL,得到100 mg·mL-1莖葉和根系的浸提液。
根際土壤浸提液:抖根法[24]獲取根際土壤,自然風干研細后過60目篩,取100 g土樣,加入1 000 mL蒸餾水浸泡48 h后,2500 r·min-1離心4 min,取上清液,重復3次,棄沉淀,將上清液定容至1 000 mL,得到100 mg·mL-1根際土壤水浸提液。
將上述浸提液分別稀釋成10.00,25.00,50.00 mg·mL-1的3個濃度梯度。置于4℃的冰箱保存備用,使用時提前取出室內靜置一段時間,避免驟冷對種子造成影響。
1.2.2.2 室內生測實驗
選取大小均勻、籽粒飽滿的白菜種子,1%次氯酸鈉溶液浸泡10 min消毒,蒸餾水反復沖洗后室內自然晾干。將種子置于鋪有2層濾紙的培養皿(Φ 9 cm)內,50粒·皿-1,分別加入不同濃度的浸提液2.50 mL,透明密封膜封閉,以添加等量蒸餾水處理為對照(CK),每處理3次重復。25℃,光暗各12 h的光照培養箱進行培養。每隔24 h統計1次種子萌發情況(以胚根達到種子的一半為萌發標準),第7 d統計萌發率,并隨即選取10株幼苗測定苗高、根長、鮮重。計算出發芽率(germination rate,GR)即7 d內供試種子發芽數/供試種子數)×100%;發芽勢(germination energy, GE)即前3 d發芽種子總數/供試種子總數×100%。
參照Williamson等[25]的方法計算化感效應指數(response index,RI):
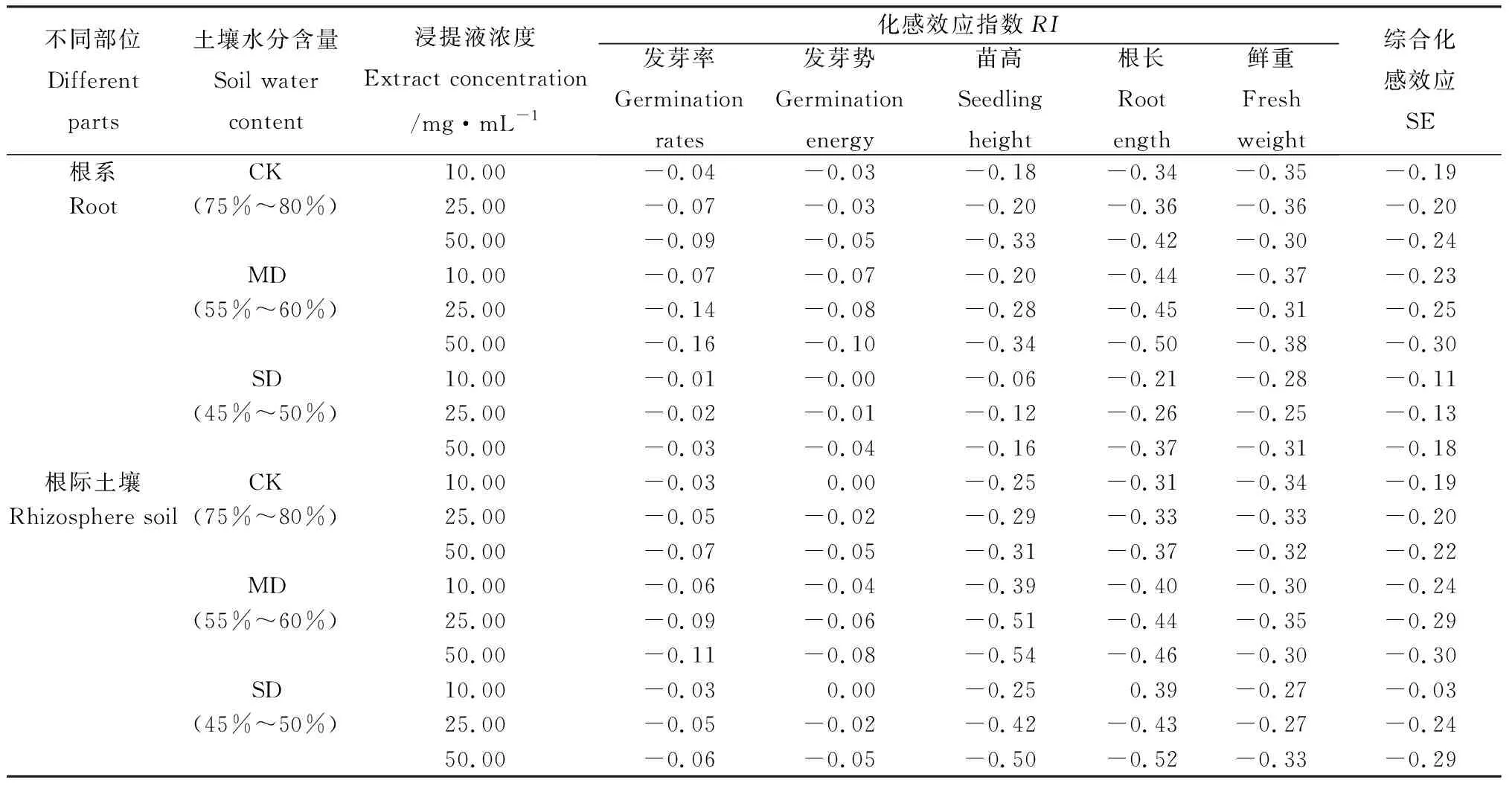
RI=1-C/T(T≥C)或RI=T/C-1(T 式中,C為對照值,T為處理值。RI> 0為促進作用,RI< 0為抑制作用,其絕對值大小反應化感作用的強弱[26]。 綜合化感效應(synthetical allelopathic effect,SE):供體植物對受體植物的各個測試指標RI的算 術平均值[27]。本研究通過SE值反映不同土壤含水量中不同部位浸提液化感效應的強弱,其計算公式為SE=(RI發芽率+RI發芽勢+RI苗高+RI根長+RI鮮重)/ 5 SPSS 18.0進行單因素方差分析,Duncan多重比較法檢驗進行差異顯著性分析,所有數據均為平均值±標準誤。 2.1.1對生物量分配的影響 節節麥植株總生物量隨土壤水分的下降而逐漸降低(表1)。當土壤含水量降低至MD、SD時,總生物量較CK分別降低了16.10%,21.19%,但均未達到差異顯著水平(P<0.05)。構件生物量分配中,根、穗生物量比均隨土壤含水量下降呈逐漸增加的變化,土壤含水量下降至MD時,根、穗生物量比與CK均達到差異顯著水平(P<0.05),而SD差異不顯著(P<0.05);莖、葉生物量比均隨土壤含水量下降而呈逐漸降低的變化,與CK相比,其中,莖生物量比在土壤含水量下降至SD時達到差異顯著水平(P<0.05),而葉生物量比則是在土壤含水量降低至MD時。葉重分數隨土壤含水量的下降呈逐漸降低的變化,至SD時,較CK降低了19.35%,未達到差異顯著水平(P<0.05)。根冠比則隨土壤含水量下降呈逐漸增加的變化,至SD時,較CK降低了19.35%,但未達到差異顯著水平(P<0.05)。生物量分配比中,以葉生物量比的表型可塑性指數最大,其次為穗、根和莖。 表1 不同土壤水分含量對節節麥生物量及其分配影響 Table 1 Effects of different soil water contents on biomass and allocation of A.tauschii Coss. 注:不同小寫字母表示差異顯著(P<0.05),下同 Note:The different lowercase letters mean significant differences at the 0.05 level, the same as below 2.1.2對葉片相關形態指標的影響 土壤含水量的下降導致了節節麥MLA的逐漸降低(表2),至MD,SD時,較CK分別下降了8.21%,10.26%,均未達到差異顯著水平(P<0.05)。此外,SLA,LMF及LAR均隨土壤含水量的降低呈逐漸下降的變化,且均在土壤含水量降至MD時與CK達到差異顯著水平(P<0.05)。其中,至SD時,SLA,LMF分別較CK降低了72.09%,20.22%,但與MD差異均未達到顯著水平(P<0.05)。至SD時,LAR較CK降低了78.68%,且與MD差異達到顯著水平(P<0.05)。以LAR表型可塑性指數最大,其次為SLA,LMF和MLA。 表2 不同土壤水分含量對葉片及根相關形態指標Table 2 Effects of different soil water contents on leaf and root morphological characteristics of A.tauschii Coss. 2.1.3對生長特性的影響 隨土壤水分含量的降低,RGR,NAR均呈下降變化,而WUE則呈逐漸增加趨勢(表3)。至SD時,RGR較CK降低了7.97%,未達到差異顯著水平(P<0.05);NAR較CK降低了68.32%,達到差異顯著水平,并與MD時差異顯著(P<0.05);WUE較CK增加了20.21%,未達到差異顯著水平(P<0.05)。其中,以NAR的表型可塑性指數最大,以RGR的最小。 表3 不同土壤水分含量對相對生長速率、凈同化速率等指標的影響Table 3 Effects of different soil water contents on RGR, NAR and other index of A.tauschii Coss. 從表4可知,莖葉水浸提液處理中,相同土壤含水量情況下,SE值隨浸提液濃度的增加而呈逐漸降低的趨勢,表明節節麥莖葉對白菜整體呈濃度效應的化感作用。相同浸提液濃度條件下,SE值則隨土壤含水量的降低呈先降后升的變化,如25.00 mg·mL-1水浸提液處理下,SE值從土壤含水量為CK的-0.29,降至MD的-0.33,隨后又升至SD的-0.24,表明節節麥莖葉對白菜的化感作用隨土壤含水量的降低整體呈先增強后減弱的變化。從SE值可以看出,根系及根際土壤對白菜表現出與莖葉類似的化感作用。此外,從SE值可看出,相同濃度及土壤含水量情況,莖葉處理的SE值均明顯大于根系及根際土壤,由此表明節節麥不同部位中以莖葉化感作用最強。 表4 不同土壤水分含量對節節麥化感作用的影響Table 4 Effects of different soil water contents on allelopathy of A.tauschii Coss. 續表4 表型可塑性在植物對異質環境的適應過程中發揮著重要的作用[28]。植物自身生物量分配格局的改變是對環境變化適應的結果[29-30]。如此通過個體或構件形態的改變,有利于植物對必須資源最大限度的利用[31-32]。而成功的入侵植物往往具有較廣幅的環境耐受性及對多樣化生境的占有特性[33]。在眾多環境因子中,水分對植物生長發育的影響至關重要,依據不同水分條件下植物形態、結構以及生殖分配等可塑性差異的變化,可為該植物對水分環境適應機制研究提供參考依據[34-35]。 本試驗結果表明,節節麥植株總生物量與土壤水分含量呈正相關,這是由于土壤中可用水分的減少,造成其生長發育減緩而引起生物量積累下降的結果。而至SD時,總生物量較CK下降未達到差異顯著水平(P<0.05),且試驗中未發現干旱脅迫處理中有植株死亡的現象,這也反映出節節麥具有一定的干旱適應能力。生物量分配方面,隨土壤含水量的降低,莖、葉生物量比呈下降變化,而根、穗生物量比則呈增加的變化,此外,R/C則均呈明顯的上升變化,這也與其他入侵植物[11,36-38]的研究結論一致。由此表明,節節麥同樣是通過增加根系生物量的分配,以拓展地下空間獲取生存所需水分,同時減少地上生物量分配,盡量縮小地上空間減少水分喪失,從而達到規避干旱生境脅迫的效果,這也是節節麥加強對資源獲取的生態策略之一。不同構件生物量比中,以葉生物量比表型可塑性指數最大,其次為穗、根和莖,表明節節麥主要通過葉片表型可塑性的調整適應土壤水分脅迫。葉片形態的相關指標中,平均葉面積隨土壤水分含量的下降而逐漸減小,至SD時,與CK差異未達到顯著水平(P<0.05),而比葉面積與葉面積比均較CK下降達到差異顯著水平(P<0.05),且以比葉面積的表型可塑性指數最大,進一步說明節節麥主要通過調整葉片表型可塑性,控制并降低水分散失的主要渠道,達到適應土壤干旱環境的結果,這與王林龍[39]、蔡海霞[40]研究結果一致。相對生長速率是植物形態學、生理學和解剖學特征的一個綜合反映[41],也是衡量植物總生物量積累狀況的重要指標,它受到凈同化速率等指標的影響[42]。隨土壤水分含量的降低,節節麥相對生長速率、凈同化速率均呈明顯的下降趨勢,這也與前人研究結果一致[38,43-44]。表明干旱脅迫下,較低的土壤水分含量不能滿足節節麥植株生長所需,導致其生長受到抑制。水分利用效率的高低是植物節水能力及水分生產水平的體現,較高的水分利用效率往往是耐旱植株對生境適應的重要策略[45-46]。研究表明,適度的干旱脅迫下,植物往往會提高自身水分利用效率以適應土壤中水分的減少[47-49]。本研究中,節節麥水分利用效率呈隨土壤含水量降低而逐漸升高的變化。 化感作用在植物的入侵過程中起著較為重要作用[50-51]。有研究認為,逆境脅迫下植物的化感作用會加強[52-53],如李軍等[11]對干旱脅迫中五爪金龍的化感作用研究,但也有研究認為,逆境脅迫下植物會降低對化感物質的投入[54]。如金鐘藤的相關研究[55-56],化感物質增加了其競爭力,但也需要較高的成本投入。從SE值可知,節節麥不同部位的化感作用隨土壤含水量的降低整體呈先升后降的變化,表明適度的干旱脅迫會增強節節麥的化感作用,但隨著脅迫的加劇,化感作用則逐漸減弱,表明節節麥為了生存而相應減少了對化感物質的投入。此外,相同條件下,節節麥不同部位中以莖葉的化感作用最強。1.3 數據分析
2 結果與分析
2.1 土壤水分含量對節節麥表型可塑性影響



2.2 土壤水分含量對節節麥化感作用的影響

3 討論與結論
3.1 土壤水分對節節麥表型可塑性影響
3.2 土壤水分對節節麥化感作用影響
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
