水稻地方品種‘月亮谷’純系對田間稻瘟病菌的選擇
2018-08-01 12:07:18王云月李成云鄭鳳萍劉永勝
植物保護 2018年4期
關鍵詞:水稻
夏 欣, 陳 平, 楊 偉, 徐 返, 王云月,李成云, 鄭鳳萍, 劉永勝, 謝 勇*
(1. 云南農業大學植物保護學院, 昆明 650201; 2. 云南省農業科學院農業環境資源研究所, 昆明 650205;3. 合肥工業大學食品科學與工程學院, 合肥 230009)
元陽縣地處滇南低緯高原山區,經緯度在102°27′~103°13′E、22°49′~23°19′N之間,海拔144~2 000 m。北回歸線以南,屬山地季風氣候類型。全縣耕地以山地為主,山高谷深,溝壑縱橫,海拔高度差異大,立體氣候明顯。元陽縣是一個以水稻OryzasativaL.為主要糧食作物的少數民族聚居地區,梯田稻作系統久負盛名,在20世紀80年代,水稻農家品種(landrace)多達196個,現在仍以種植農家品種為主。2013年被批準列入聯合國教科文組織《世界遺產名錄》中的文化遺產目錄。盡管水稻種植歷史悠久,絕大多數種植區都處于稻瘟病易于發生的生態環境下,但至今為止還沒有有關稻瘟病Magnaportheoryzae大面積暴發的記載[1]。尤為值得關注的是,一些地方品種種植時間已超百年,在當地稻作系統中一直未被淘汰。其中‘月亮谷’(‘Acuce’)具有優良的農藝性狀和豐富的群體間和群體內遺傳異質性(inter-and intra-heterogeneity),中抗稻瘟病菌,是在元陽哈尼梯田海拔1 600~1 900 m區域種植的主栽秈稻O.indica品種[2]。
已有的分子證據表明,稻瘟病菌-水稻寄主互作符合經典的基因對基因假說(gene for gene hypothesis),導致同一水稻品種對不同稻瘟病菌菌株表現出明顯的抗感差異[3-6]。現代水稻抗病品種抗性的“喪失”是寄主抗性基因與稻瘟菌致病基因互作在群體水平上的表現結果,即寄主品種群體對由稻瘟病菌不同生理小種(race)構成的混合群體產生定向選擇(directional selection)作用[7-8]。元陽梯田水稻種植歷史悠久,但稻瘟病發生并不嚴重,其中的科學問題值得深入探索。關于稻瘟菌的培養表型和生物學特性研究,很多學者探討了病菌的產孢條件、菌落形態及顏色、黑色素合成、產孢量、菌絲生長速率等與致病性之間的關系[9-12]。有研究表明,菌落顏色及黑色素與病原真菌的致病性緊密相關,黑色素是許多病原真菌的毒力因子,在病原真菌的致病過程中起著重要作用[13-15]。本研究采用孢子捕捉法和組織分離法,獲得梯田環境中和水稻植株上的稻瘟病菌,從表型上比較兩種來源的稻瘟菌之間的差異,初步了解農家品種‘月亮谷’對環境中稻瘟菌群體的選擇作用。
1 材料與方法
1.1 水稻品種(系)
1.1.1 田間試驗品種
供試水稻品種(系)共5個,包括來源于元陽哈尼梯田地方品種‘月亮谷’的單粒傳(single seed descend method, SSD)F8代純系L1、L2、L3、L4,由農戶自留種經8代單粒傳獲得;‘月亮谷’自然群體G5(來自于農戶自留種)。
1.1.2 抗稻瘟病單基因鑒別品系
單基因鑒別品系(NILS)采用國際水稻研究所(IRRI)以‘麗江新團黑谷’(LTH)為輪回親本培育的水稻近等基因品系:IRBLsh-s、IRBLz-Fu、IRBLb-B、IRBL12-M、IRBLt-k59、IRBLa-A、IRBLi-F5、IRBL3-CP4、IRBL5-M、IRBLks-S、IRBLkm-Ts、IRBL1-CL、IRBLkh-k3、IRBLk-k、IRBLkp-k60、IRBLta-k1、IRBL7-M、IRBL9-W、IRBLz5-CA-1、IRBLzt-T、IRBLta2-Pi、IRBLta2-Re、IRBLta-CP1、IRBL19-A、IRBL20-IR24共25個(種子由IRRI 的Kobayashi博士提供),感病對照品種為‘麗江新團黑谷’。
1.2 田間試驗設計
供試的5個水稻品種(系)按元陽哈尼梯田(元陽縣新街鎮箐口村,23°07′N,102°44′E,海拔1 637.7 m)當地常規育苗方法育苗、移栽。每品種(系)為1個處理,3次重復,每小區30 m2,完全隨機區組排列。4周保護行種植‘月亮谷’自然群體。所有施肥、水分管理均為當地傳統方法。
1.3 稻瘟菌的分離
1.3.1 供試水稻品種(系)上稻瘟菌的分離
在穗頸瘟發生期(9月),從試驗田間分別采集5個水稻品種(系)的感病樣本,采用毛細管單孢分離法進行單孢分離,獲得來源于水稻寄主的稻瘟病菌株(寄主菌株)[16]。
1.3.2 梯田環境中稻瘟菌的獲得
采用便攜式孢子捕捉儀(YFBZ3,河南云飛科技發展有限公司),按說明書進行田間孢子捕捉,持續時間為2周。具體方法為:在上述試驗田中按5點取樣法放置孢子捕捉儀,每次捕捉時間從上午9時開始定時開機5~8 h,然后采用單孢分離法在體視顯微鏡下分離稻瘟菌,獲得來源于梯田環境中的稻瘟病菌株(環境菌株)[17]。
1.4 稻瘟菌的培養與形態觀察
米糠培養基:米糠經過粉碎后,稱取20 g,加水煮15 min,然后加入瓊脂16 g,酵母浸膏2 g,定容到1 L,混勻分裝,于121℃高壓滅菌40 min。
將分離純化的菌株接種到米糠培養基上,在L∥D=12 h∥12 h,24~26℃下培養9 d后用直徑為9 mm的打孔器分別打取菌餅,菌餅朝下轉移到新鮮米糠培養基上,在26℃下培養9 d后,用數碼照相機拍照,記錄菌落形態特征。刮除培養基上菌落的表面菌絲,觀察黑色素顏色并拍照。
1.5 稻瘟菌菌絲生長速率測定
分別將寄主菌株(4、8、40、45、128)和環境菌株(139、141、203、244、287)的稻瘟菌單孢菌株接種于米糠培養基,26℃培養7 d,用打孔器(直徑5 mm)分別從菌落的邊緣處打取菌餅,將菌餅朝下在無菌條件下轉移到米糠培養基中央,置于26℃條件下培養9 d。采用十字交叉法垂直測量菌落直徑。每隔3 d測量1次菌落生長直徑,根據所測結果計算純生長量。
純生長量(mm)=菌落平均直徑(mm)-菌餅直徑(mm);
日均生長量=純生長量(mm)/生長天數。
1.6 稻瘟菌產孢量測定
將兩種不同來源的各5個稻瘟菌單孢菌株分別在米糠培養基上培養7 d,從菌落邊緣同一圓周上用打孔器取菌碟,轉移到新的培養基上,設置6次重復,放在28℃培養箱,培養9 d后,將10 mL無菌水倒入平板中,用接種環刮取菌絲和孢子,振蕩儀振蕩2 min,紗布過濾,定容至10 mL,在10倍顯微鏡下,用血球計數板計算孢子濃度。孢子濃度=(5個小方格孢子總數/80)×4×106。
1.7 抗稻瘟病單基因鑒別品系的接種與致病型測定
將水稻單基因鑒別系浸種催芽后播種于塑料盆中(60 cm×40 cm×25 cm),栽培土壤為滅菌稻田土,每70 L中加入2 L火山石混勻。每品種播15粒,設3次重復。在溫室內育苗,2 葉期開始每周追施尿素0.5 g/盒,共2~3次。分別從寄主菌株和環境菌株群體中各隨機挑選5個菌株作為接種菌株,于水稻秧苗3.5葉期進行噴霧接種,秧苗接種后置恒溫保濕培養箱內于25℃、相對濕度≥95%,黑暗條件下保濕培養24 h,隨后移出置溫室內培養,每天用清水噴霧以保持溫室的濕度。接種7 d后參照Gallet等[18]的方法進行病害調查。
病害調查標準為[18]:1級,沒有病斑;2級,有少量針尖點狀褐色病斑;3級,有大量針尖點狀褐色病斑,或較大褐色病斑;4級,在褐色病斑中間有灰白色壞死組織;5級,灰色壞死組織擴大,病斑呈典型梭形病斑;6級,灰白色病斑或很大的梭形病斑,周圍沒有或有少量褐色包圍。
抗感分級標準:1~3級為抗病反應(R),4~6級為感病反應(S)。根據抗感反應表型分別統計接種菌株對某個鑒別品系的毒力頻率(virulence frequency,VF)和測試菌株對所有鑒別品系的聯合致病性(pathogenicity association,PA)[19]。
毒力頻率=(有毒力菌株數量/菌株總數量)×100%;
聯合致病性=(接種菌株群內對某品種的致病菌株數量/該群內總菌株數量)×100%。
1.8 數據整理與統計分析
采用Excel軟件整理原始數據,顯著性檢驗和分析由SPSS(Version 18.0)軟件的Wilcoxon成對測驗法(Wilcoxon Matched Pairs Test)完成。
2 結果與分析
2.1 兩種不同來源的稻瘟菌株菌落形態及黑色素產生情況
環境菌株(4、128、40、45、8)和寄主菌株(203、139、287、141、244)的菌落正面都近似圓形,菌落顏色從中心向周圍逐漸變淺,氣生菌絲逐漸變稀疏,其中部分菌株出現同心輪紋。環境菌株在米糠培養基表面生長時菌落疏松,呈絨毛狀,放射散生在培養基表面,菌落顏色從中心到邊緣依次呈白色、灰白色,菌落背面顏色較淺,產生少量黑色素;寄主菌株在米糠培養基上的菌落較致密,呈地毯狀,匍匐蔓延在培養基表面,菌落顏色從中心到邊緣依次呈現灰白色、褐色,背面顏色較深,產生黑色素較多。與環境菌株4、128、40、45、8相比,寄主菌株203、139、287、141、 244菌落顏色普遍較深,說明黑色素合成較多(圖1)。

圖1 不同來源的稻瘟病菌菌落形態及產生黑色素情況Fig.1 Observation of colony morphology and melanin formation of Magnaporthe oryzae isolates
2.2 稻瘟菌株菌落生長速率比較
稻瘟菌株分別在培養3、6、9 d時,菌落直徑總體差異不大,到后期生長曲線總體趨于平緩。環境菌株生長速率波動不明顯,而寄主菌株生長速率波動相對較大,其中菌株287生長速率最快,平均生長直徑均大于其他菌株,寄主菌株139生長最慢(圖2)。總體上,環境菌株菌絲生長速率與寄主菌株菌絲生長速率之間沒有顯著差異。
2.3 兩種不同來源的稻瘟菌產孢量比較
在參測的10個菌株中,寄主菌株244產孢量最大,顯著高于其他菌株,寄主菌株287產孢量最低,顯著低于環境菌株40、45、128和寄主菌株139、203、141和244,另一個菌株203產孢量也較低。比較梯田環境菌株,除菌株45產孢量與菌株4、8差異顯著外,與其他梯田環境菌株差異不顯著。而對于寄主菌株139、141,產孢量顯著低于菌株244,但顯著高于287、203。以上結果表明,梯田環境菌株總體產孢量與寄主菌株產孢量之間沒有顯著差異,但個別菌株之間產孢量差異較大。

圖2 不同來源的稻瘟菌菌株菌絲生長速率Fig.2 Mycelial growth rate of Magnaporthe oryzae strains obtained from different sources

圖3 環境稻瘟菌菌株與寄主菌株產孢量的比較Fig.3 The spore quantity produced by Magnaporthe oryzae isolates obtained from different sources
2.4 兩種不同來源的稻瘟菌致病性分析
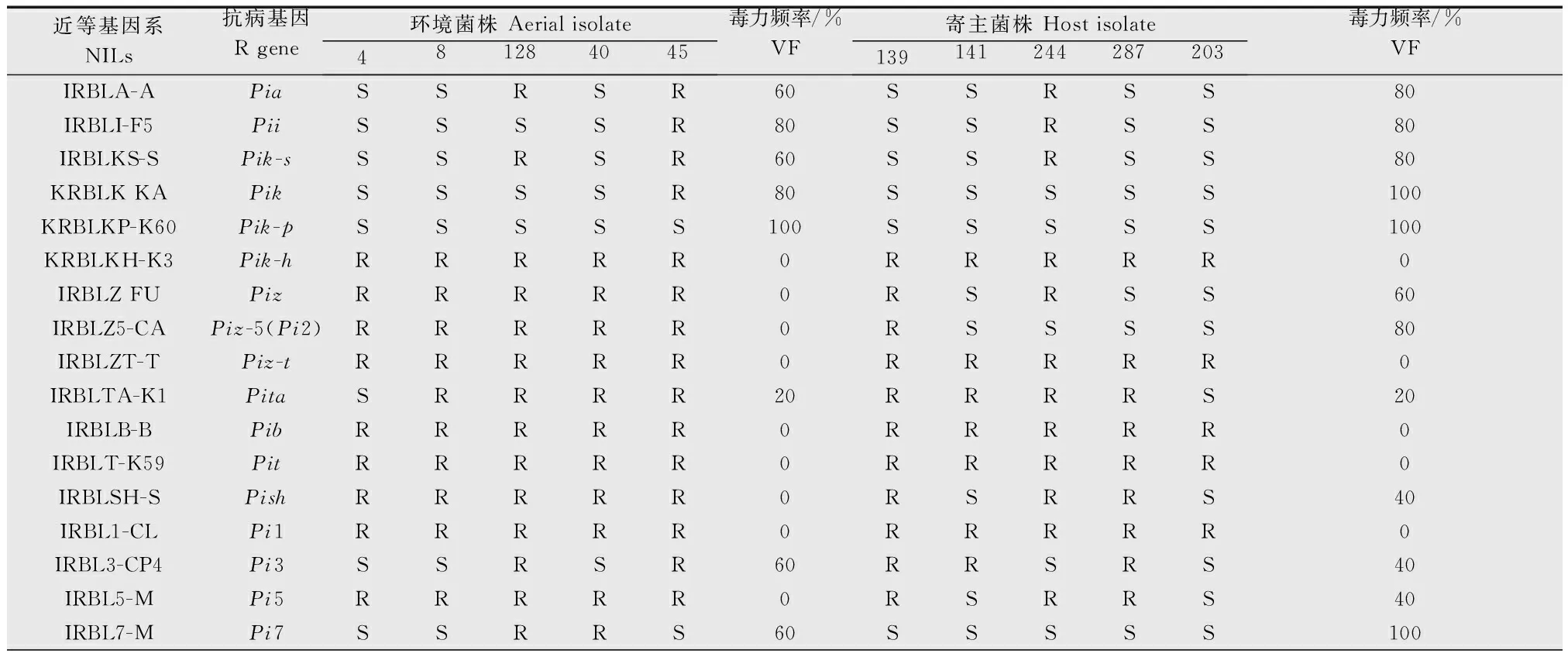
從環境菌株和寄主菌株中各隨機挑選5個菌株,用25個近等基因系(NILs)測定其致病性。結果表明,總體上近等基因系鑒別品種對來源于環境中的稻瘟菌菌株的抗性高于水稻寄主上的稻瘟菌菌株,平均毒力頻率為24.8%,而來源于水稻寄主上的菌株毒力達到38.4%,聯合致病性也顯示出來源于水稻寄主上的菌株比來源于環境的菌株致病性更高。初步結果表明,寄主水稻上的菌株其致病性強于梯田環境菌株(表1)。
根據基因對基因理論,只有抗病基因與相應的無毒基因才能產生非親和性互作(incompatible interaction)。從本試驗結果可以看出,在元陽梯田種植攜帶質量抗性基因Pik-h、Piz-t、Pib、Pit、Pi1、Piz-5(Pi2)、Pi11、Pi12、Pik-m、Pi20、Pita-2的品種其稻瘟病發生的風險將顯著降低,若僅攜帶Pik、Pik-p、Pi7和Pi19則病害發生的風險較高。
2.5 稻瘟菌菌株致病性與生物學特性之間的關系
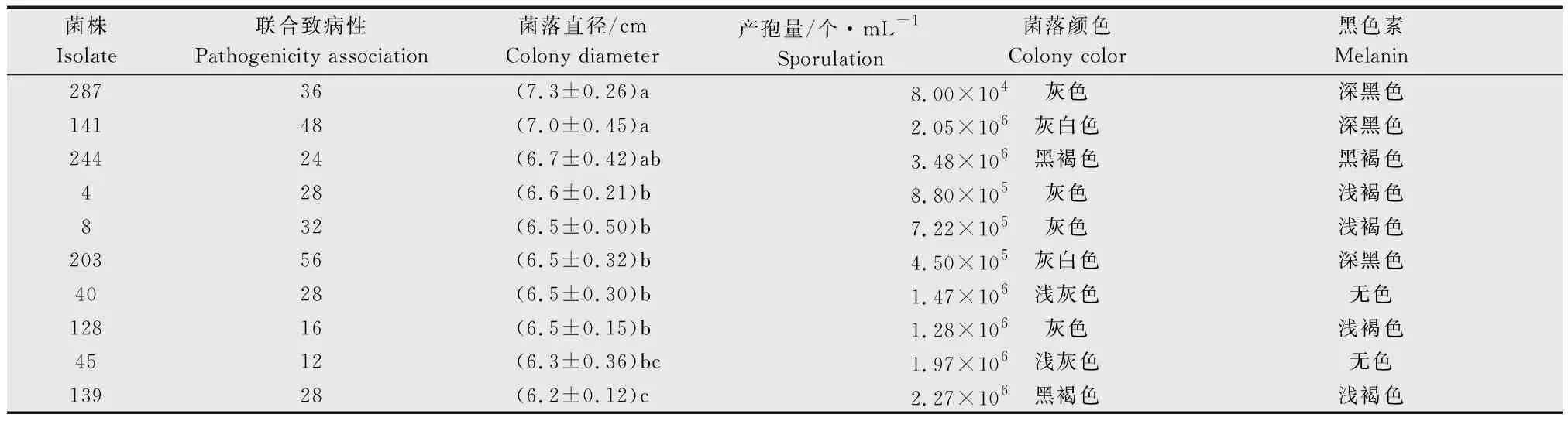
為了解培養性狀與菌株致病性間的關系,分析了不同菌株的聯合致病性與菌落顏色、黑色素深淺、菌株生長速率、產孢量之間的關系。結果(表2)表明,菌株的致病性與生長速率和產孢量之間無明顯相關關系。而與黑色素的產生有一定關系,即黑色素產生越多,顏色越深,致病性越強。如來源于水稻寄主的菌株203、141、287和244的聯合致病性均高于24.0%。而產生黑色素較少的菌株,聯合致病性相對較低。
表1稻瘟菌菌株在鑒別近等基因系上人工接種的致病表型鑒定結果
Table1PathogenicityassayofMagnaportheoryzaeisolatesonNILSbyusingartificialinoculation
近等基因系NILs抗病基因R gene環境菌株 Aerial isolate481284045毒力頻率/%VF寄主菌株 Host isolate139141244287203毒力頻率/%VFIRBLA-APiaSSRSR60SSRSS80IRBLI-F5PiiSSSSR80SSRSS80IRBLKS-SPik-sSSRSR60SSRSS80KRBLK KAPikSSSSR80SSSSS100KRBLKP-K60Pik-pSSSSS100SSSSS100KRBLKH-K3Pik-hRRRRR0RRRRR0IRBLZ FUPizRRRRR0RSRSS60IRBLZ5-CAPiz-5(Pi2)RRRRR0RSSSS80IRBLZT-TPiz-tRRRRR0RRRRR0IRBLTA-K1PitaSRRRR20RRRRS20IRBLB-BPibRRRRR0RRRRR0IRBLT-K59PitRRRRR0RRRRR0IRBLSH-SPishRRRRR0RSRRS40IRBL1-CLPi1RRRRR0RRRRR0IRBL3-CP4Pi3SSRSR60RRSRS40IRBL5-MPi5RRRRR0RSRRS40IRBL7-MPi7SSRRS60SSSSS100
續表1Table1(Continued)

近等基因系NILs抗病基因R gene環境菌株 Aerial isolate481284045毒力頻率/%VF寄主菌株 Host isolate139141244287203毒力頻率/%VFIRBL9-WPi9RRRRR0RSRRS40IRBL12-MPi12RRRRR0RRRRR0IRBL19-APi19SSSSS100SSSSS100IRBLKM-TSPik-mRRRRR0RRRRR0IRBL20-IR24Pi20RRRRR0RRRRR0IRBLTA2-PIPita-2RRRRR0RRRRR0IRBL11-ZHPi11RRRRR0RRRRR0IRBLZ5-CAPiz-5(Pi2)RRRRR0RRRRR0LTH-SSSSS100SSSSS100聯合致病性/%Pathogenicity association36321628122848243656平均 Average24.838.4
表2菌株致病性與菌株培養性狀及表型的比較1)
Table2ComparisonbetweenpathogenicityandphenotypictraitsofMagnaportheoryzaeisolates
菌株Isolate聯合致病性Pathogenicity association菌落直徑/cmColony diameter產孢量/個·mL-1Sporulation菌落顏色Colony color黑色素Melanin28736(7.3±0.26)a8.00×104灰色深黑色14148(7.0±0.45)a2.05×106灰白色深黑色24424(6.7±0.42)ab3.48×106黑褐色黑褐色428(6.6±0.21)b8.80×105灰色淺褐色832(6.5±0.50)b7.22×105灰色淺褐色20356(6.5±0.32)b4.50×105灰白色深黑色4028(6.5±0.30)b1.47×106淺灰色無色12816(6.5±0.15)b1.28×106灰色淺褐色4512(6.3±0.36)bc1.97×106淺灰色無色13928(6.2±0.12)c2.27×106黑褐色淺褐色
1) 圖中英文字母a、b、c表示在P<0.05水平上差異顯著。
The letters a, b and c indicate significant difference at 0.05 level.
3 討論
寄主-病原菌之間的協同進化(co-evolution)是兩者遺傳變異的驅動因素之一。在長期的水稻栽培過程中,稻瘟菌與水稻寄主在同一生態條件和環境因子的作用下,稻瘟菌的優勢生理小種或遺傳宗群(lineage)也會發生相應變化[19]。原因是不同水稻品種可能攜帶不同的抗病基因,對稻瘟菌群體產生不同的選擇壓力。因此,稻瘟菌生理小種的分化明顯。有報道,我國南方與北方稻作區稻瘟菌小種的組成差異較大[20-21]。元陽哈尼梯田稻作系統相對封閉,農家品種豐富,采用獨有的生產方式。推測在這種環境條件下,農家品種對稻瘟菌群體在時間和空間(temporal and spatial)結構上的影響將會表現出不同的作用方式。農家品種是一個不同基因型的混合群體[2, 22-24],本文分析了‘月亮谷’單粒傳純系對梯田環境中稻瘟菌群體的選擇作用,初步研究了稻瘟菌的表型性狀與致病性的關系。結果表明,不同來源菌株在相同培養基上的培養性狀有明顯差異,菌落形態差異較大,說明元陽哈尼梯田稻瘟菌群體具有豐富的多樣性。但菌落特征、生長速率、產孢能力等形態特征與其來源沒有對應關系。
黑色素(melanin)在植物病原真菌中廣泛存在,與病原菌致病能力密切相關。稻瘟菌通過提高細胞膨壓,機械突破水稻寄主表皮[25-26]。本研究結果表明,在米糠培養基上,寄主菌株菌落顏色呈白色、灰白色,顏色深淺不均勻,除個別菌株外,產生黑色素能力均較強,梯田環境菌株菌落顏色普遍較淺,稻瘟菌株產生黑色素能力相對較弱。由于‘月亮谷’對稻瘟病總體上屬于中抗(MR)水平[2],因此從‘月亮谷’單粒傳品系上分離到的稻瘟菌菌株比從環境中(可能來源于其他地方品種)捕獲的菌株致病性強。
為明確本研究中稻瘟菌菌株可能攜帶的致病因子或無毒基因,采用以‘麗江新團黑谷’為輪回親本構建的單基因鑒別品系進行人工接種試驗。分別測定了來源于梯田環境和寄主‘月亮谷’純系的稻瘟菌菌株的聯合致病性、毒力頻率以及測定菌株可能攜帶的無毒基因種類。結果顯示,梯田環境中的稻瘟菌具有豐富的致病表型,總體上來源于‘月亮谷’的菌株具有更高的毒力頻率(38.4 vs 24.8)。顯然,從‘月亮谷’上分離到的菌株是梯田環境中菌株經過抗性選擇的結果,如果僅考慮與這25個抗性位點相關的遺傳位點,說明盡管‘月亮谷’在當地種植了上百年,與稻瘟病菌間的協同進化仍然處于動態進化過程中,遠未達到哈迪-溫伯格平衡(Hardy-Weinberg equilibrium)。這也許與哈尼族的輪種或換種習慣有關[22,27]。
根據基因對基因假說,從對25個單基因鑒別品系的致病性測定結果可以推導出參測的稻瘟菌菌株的無毒基因種類,即抗病基因Pik-h、Piz-t、Pib、Pit、Pi1、Piz-5(Pi2)、Pi11、Pi12、Pik-m、Pi20、Pita-2相對的無毒基因。由于本研究的樣本量較小,還需擴大樣本數,采用群體遺傳研究策略,進一步測定代表單倍型(haplotype)及其可能攜帶的無毒基因種類,在清楚當地主要農家品種的抗性基因組成的基礎上,才能更好地指導哈尼梯田農家品種科學合理布局,有助于當地水稻生產和元陽哈尼梯田非物質文化遺產的持續保護。
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
