玉米葉片原生質體的制備及瞬時轉化體系的建立
2018-08-28 06:50:30雷海英白鳳麟郭亞沖喬楠茜
長治學院學報 2018年2期
雷海英,白鳳麟,馮 宇,郭亞沖,喬楠茜
(長治學院 生物科學與技術系,山西 長治 046011)
1 引言
植物原生質體是指植物細胞用一些手段如酶解法、機械法等除去細胞壁,形成由單層細胞膜包裹的細胞質[1]。原生質體由于沒有細胞壁,相對容易攝取外源遺傳物質,使外源DNA在原生質體中快速表達并可按既定方向進行培養,是植物作為遺傳轉化的理想材料[2],同時原生質體也是獲得細胞無性系和選育突變的優良起始材料[3],因此,原生質體是開展基礎研究的理想材料。玉米原生質體由于制備方法相對簡單,轉化效率較高,常用于植物瞬時表達系統。
植物瞬時表達檢測是一種快速的、高通量、轉化簡單的分析系統,具有方便快捷的優點。結合報告基因的使用,使得研究基因的表達體系變得更加快捷,可廣泛用于基因功能的研究[4]。原生質體的制備和純化對植物瞬時表達體系的建立起著決定性的作用,對研究基因的功能和調控途徑的研究具有重要意義。盡管玉米原生質體的制備研究已經有報道,但要獲得質量且轉化能力也高的原生質體尚需要不斷摸索。
本實驗以玉米自交系鄭58為材料,對玉米原生質體的制備過程進行了優化,通過PEG法介導質粒轉化原生質體,建立了玉米原生質體可高效轉化的瞬時表達系統。
2 實驗方法
2.1 材料與試劑
試驗所用玉米(Zea mays L.)自交系材料鄭58種子由山西省農業科學院生物技術研究中心提供。在本實驗室花盆種植,培養箱中28℃遮光培養7 d左右,待玉米長至二葉期至三葉期時,取第二片嫩葉制備原生質體。所用纖維素酶R-10、離析酶R-10購自Yakult,Japan,其余試劑購自國內經銷商。
2.2 玉米原生質體的制備
酶解液的配制:纖維素酶R-10、離析酶R-10、D- 甘露醇、0.05 moL/L KCl、0.4 moL MES(pH=5.7)混勻后55℃水浴至溶液清亮,待冷卻至室溫后加入0.2 moL/L CaCl2、β-巰基乙醇、BSA混勻備用。取種植7天的玉米第二片伸展的葉片約0.3 g,去除頭尾,用11號無菌刀片蘸取適量的W5(NaCl 154 mmoL/L、CaCl2 125 mmoL/L、MES 2 mmoL/L),在無菌培養皿中將玉米葉片切成1 mm左右的細絲,將切好的玉米葉片迅速完全浸沒在酶解液中,28℃,55 rpm/min,振蕩,于黑暗中酶解4-6 h。停止反應后,用W5潤濕200目的無菌濾網,取培養好的酶解液過濾,加入等體積的W5稀釋,在光學顯微鏡下觀察原生質體。
2.3 玉米原生質體的純化
將過濾后的原生質體原液不同離心力離心2 min,棄上清,緩慢加入1 mL W5,指彈混勻,離心2 min,棄上清,加入100 μL W5觀察,利用原生質體簡易計數法計數[5]。
2.4 原生質體提取條件的優化
采用不同酶解濃度(纖維素酶和離析酶的濃度分別為0.5%、1.0%、1.5%)組成7種組合,對原生質體產量與質量進行分析;確定最佳的酶解組合過程中同時對不同滲透壓濃度、離心力大小進行分析,得出提取完整好的原生質體最佳離心力,最終得出提取玉米原生質體最佳的體系。
2.5 原生質體的轉化
取完整的純化后的原生質體液中加入100 μL MMg(甘露醇 0.4 moL/L、MgCl2 15 mmoL/L、MES 2 mmoL/L)重懸,轉化本實驗室構建好的含有目的基因的 GFP標簽重組載體:pCAMBIA1302-mGFP-m-c、pCAMBIA1302-mGFP-C、pCAMBIA1302-mGFP(空載體),轉化質粒濃度為 10 μg/mL。混勻后加入 30%PEG-4000 CaCl2,23℃避光放置15 min。加入2倍體積W5終止反應,100 g離心2 min,棄上清,加入100 μL WI(4 mmoL/L MES、0.4 moL/L 甘露醇、5 mmoL/L KCl),混勻后加入24孔板中,加入WI至終體積為250 μL,23℃暗培養12-14 h,在激光共聚焦顯微鏡下(LSM 880,卡爾蔡司,德國)觀察轉化效果。
3 結果與討論
所有的原生質提取結果如圖1所示,產率最高一般為折線圖最高點,但從轉化結果看來,滲透壓為0.5 mol/L時,轉化效率低于0.4 mol/L時的甘露醇。具體結果如下。
3.1 不同酶組合對原生質體的影響
取不同濃度的離析酶R-10、纖維素酶R-10的組合配制酶解液,通過對原生質體進行觀察和計數,得出提取原生質體時的最佳酶濃度為纖維素酶1.0%、離析酶 0.5%(表 1)。
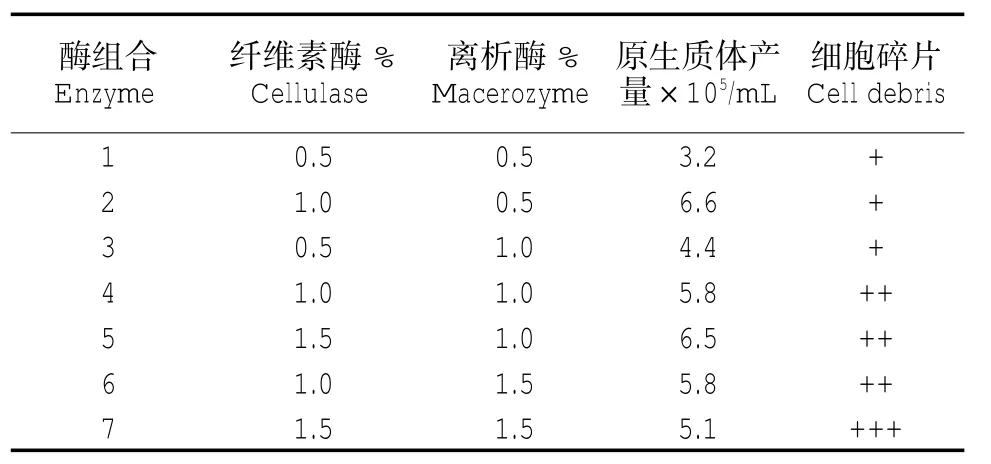
表1 不同酶組合對原生質體分離效果的影響Tab.1 Effects of different enzyme combinations for protoplast isolation effect
3.2 不同酶解時間對原生質體制備提純的影響
酶解時間以0.5 h為時間間隔依次取樣3 h、3.5 h、4 h、4.5 h、5 h、5.5 h、6 h 的原生質體,每次取2 mL樣品,離心后用W5懸浮,用量100 μL,再在普通光學顯微鏡下觀察原生質體的完整度并用血球計數板統計原生質體的產量。結果表明,當酶解時間過短時酶解不充分,酶解液中葉片幾乎完整呈黃綠色,原生質體沒有全部釋放出來,原生質體產量不高;當酶解時間過長時破碎嚴重,推測是由于長時間的酶解使部分已解離的完整原生質體受到損傷進而破碎。最適酶解時間為5 h,其原生質體活力較高,完整度好,飽滿(圖1中A-C)。
3.3 不同滲透壓對原生質體的影響
當甘露醇濃度在0.3 mol/L時,提取的原生質體的體積會吸水膨脹,可知酶解液滲透壓低于原生質體滲透壓;當甘露醇濃度在0.4 mol/L時,提取出的原生質體完整度較高,破損率低,原生質體滲透壓與酶解液滲透壓逐漸平衡,原生質體吸水漲破現象減少;當甘露醇濃度為0.5 mol/L時,提取的原生質體產量提高,但原生質體會收縮。
甘露醇濃度在0.4 mol/L時玉米原生質體與酶解液的滲透壓相近,提取的原生質體完整度最高,產量適宜,破損率低(圖1中D-F)。
3.4 不同離心條件對原生質體的影響
通過取離心力分別為 50×g、100×g、150×g、200×g、250×g、300×g,離心時間 1 min、2 min、3 min、4 min、5 min,從普通光學顯微鏡下觀察到的原生質體完整度看來,當離心力為100×g時完整度較高,隨著離心力的增大,玉米原生質體的完整度逐漸降低,當離心力達到400×g時原生質體破碎度嚴重,因而最佳離心力大小為100×g、離心1 min(如圖 1 中 G-I)。
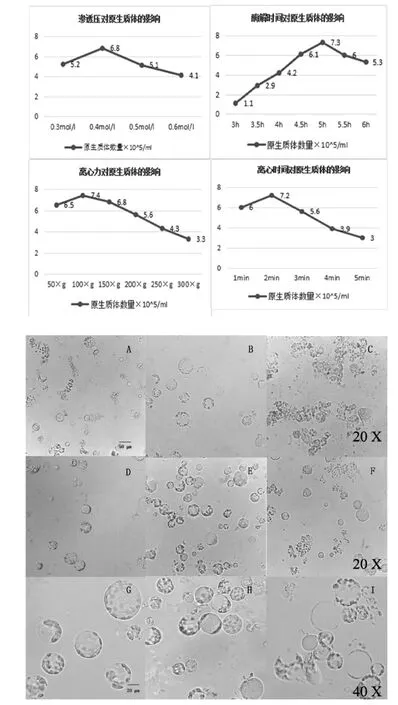
圖1 不同條件下對玉米原生質體產量的影響及顯微觀察結果Fig.1 Yield of different factors protoplasts isolation from leaves of maize
3.5 原生質體瞬時轉化
由圖2所示,將本實驗室研究基因[6-7]功能用帶有綠色熒光蛋白標簽(GFP)的空載體pCAMBIA1302-mGFP(A-C)有微弱的自發熒光,而含有研究的目的基因玉米中心蛋白基因(centrin)的重組載體pCAMBIA1302-mGFP-C(D-G)在細胞核位置表達較亮的綠色熒光,表明瞬時體系轉化成功,且蛋白表達定位與細胞核中。
玉米原生質體制備及轉化的關鍵,首先要選擇暗培養一周左右的玉米的第二片葉片,此時的玉米葉片中的細胞活力高,利于原生質體的提取。其次玉米葉片要切的足夠細,且切絲時間不能太長,會導致葉片變干從而影響原生質體的產量。離心:由于玉米原生質體非常脆弱,為減少對原生質體的破壞,實驗過程中所有的離心步驟所用的均是平轉子。滲透壓:隨著酶解液滲透壓的增大,原生質體的產量逐漸提高。這是因為當酶解液滲透壓低于玉米原生質體內部滲透時,部分原生質體會吸水破裂,酶解液滲透壓增大后,原生質體滲透壓與酶解液滲透壓逐漸平衡,原生質體吸水漲破現象減少,玉米原生質體產量會提高[8]。可見滲透壓濃度在0.4 mol/L時玉米原生質體與酶解液的滲透壓相近。PEG濃度:PEG介導[9]原生質體轉化時,實驗中PEG的濃度不能太高,30%比較合適,因為它對細胞有一定的毒害作用,高濃度的PEG會導致細胞破碎。最后操作過程:所有操作過程都需柔和,以免激烈震蕩使原生質體破損,影響后續實驗。
4 結論
對玉米(Zea mays L.)自交系-鄭58葉肉細胞原生質體的制備條件的優化后,沿著葉脈橫切葉片,最佳酶濃度組合為纖維素酶1.0%,離析酶0.5%;最佳酶解時間為5 h;最佳滲透壓甘露醇濃度為0.4 mol/L;離心力為100×g、離心1 min時原生質體的產量最高,并轉化質粒獲得了理想的結果,為今后研究目的基因的亞細胞定位及功能分析等奠定了基礎。
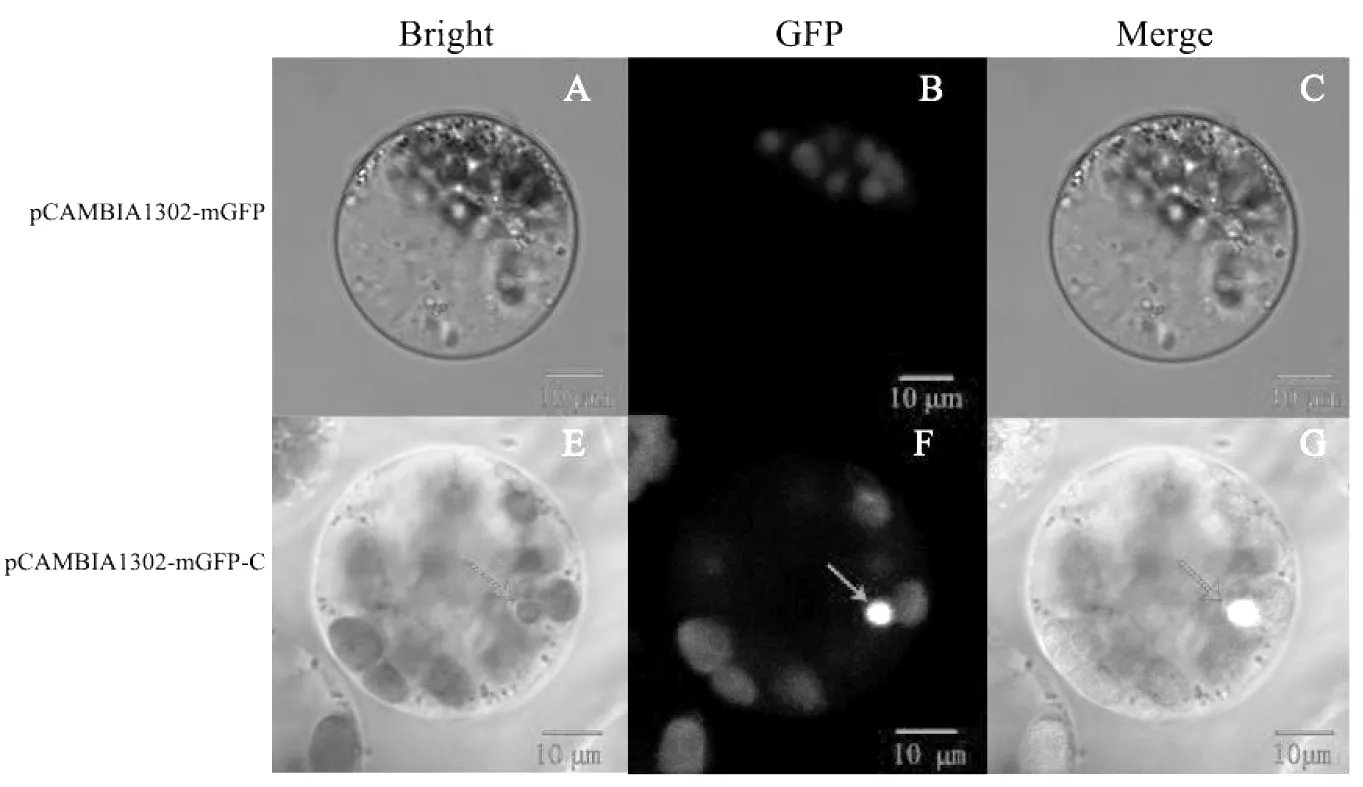
圖2 載體瞬時轉化玉米原生質體Fig.2 maize protoplasts transiently transfected with pCAMBIA1302-mGFP and pCAMBIA-1302-mGFP-C plasmids
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
今日農業(2021年14期)2021-11-25 23:57:29
中國民間療法(2021年5期)2021-06-09 09:21:26
石油與天然氣地質(2021年1期)2021-02-22 14:14:44
今日農業(2020年20期)2020-11-26 06:09:10
中國果業信息(2019年10期)2019-11-13 01:21:34
聚氯乙烯(2018年9期)2018-02-18 01:11:34
食藥用菌(2016年6期)2016-03-01 03:24:27
護士進修雜志(2015年5期)2015-03-18 19:22:53
中國藥業(2014年24期)2014-05-26 09:00:09
