人腦自適應多尺度功能連接的性別差異*
2018-08-30 01:59:02段凱凱董昊銘苗麗雯蘇學權左西年
心理科學進展 2018年9期
段凱凱 董昊銘 苗麗雯 蘇學權 相 潔 左西年,7
?
人腦自適應多尺度功能連接的性別差異*
段凱凱1董昊銘2,3,4,5苗麗雯1蘇學權6,7相 潔1左西年2,3,4,5,7
(1太原理工大學, 計算機科學與技術學院, 太原 030024) (2中國科學院行為科學重點實驗室, 北京 100101)(3中國科學院大學, 心理學系, 北京 100049) (4中國科學院心理研究所磁共振成像研究中心, 北京 100101)(5中國科學院心理研究所, 腦與心智畢生發展研究中心, 北京 100101) (6廣西師范學院, 體育學院, 南寧 530000) (7廣西師范學院, 腦與教育重點實驗室, 南寧 530001)
已有腦成像研究展示了男女腦功能差異, 但功能磁共振信號的頻率劃分通常基于主觀經驗, 使腦功能性別差異的生物學解釋遭遇瓶頸。本文提出人腦自適應多尺度功能連接算法, 刻畫功能連接的時空多尺度特性, 揭示出0.06~0.10 Hz的性別差異:男性較強的連接主要與邊緣網絡和腹側注意網絡有關, 女性較強的連接主要與腹側注意網絡、視覺網絡和額頂網絡有關。
性別差異; 功能連接; 腦網絡; 功能磁共振成像; 低頻特性

1 引言
近年來, 隨著認知神經科學的快速發展和對功能磁共振成像(functional Magnetic Resonance Imaging, fMRI)了解的深入, 有關腦信號的頻率研究也被廣泛關注。fMRI信號來源于大腦血液中脫氧血紅蛋白含量的動態變化, 是一種非平穩非線性變化的信號, 具有復雜的時頻特征。另外, fMRI信號是大腦復雜的機理性反映, 不同頻段節律信號的生理意義尚無定論, 對fMRI信號進行時頻分析是當下一種較為熱門的研究方向, 但是在已有的研究中, 研究人員多根據主觀經驗或經典方法將其劃分, 缺乏統一的分頻標準, 致使該領域內相關分頻研究的可重復性較差, 同時由于經典算法對數據的假設較多, 未能充分發掘功能磁共振信號的自身特性, 也阻礙了我們進一步了解它的內在生理意義。例如, 在對fMRI低頻成分的腦功能網絡研究中, Wu等人(2008)用切比雪夫濾波器將fMRI信號分為12個頻段, 并探討了功能連接的頻率特性, 發現具有高功能連接系數的腦功能活動集中在特定的低頻段(0.01~0.06 Hz); Salvador, Suckling, Schwarzbauer和Bullmore (2005)以及Salvador等人(2008)基于傅里葉方法對信號分頻揭示了較高功能連接系數的腦功能活動分布在低頻段(<0.10 Hz); Achard, Salvador, Whitcher, Suckling和Bullmore (2006)利用小波變換方法分為6個頻段, 證明了低頻成分(0.03~0.06 Hz)的全腦相關性最高; Zuo等人(2010)基于濾波器方法揭示了低頻振幅在(0.027~0.073 Hz)頻段有更強的幅值; Liang等人(2012)采用濾波器方法驗證了低頻段(0.027~ 0.073 Hz)形成的腦網絡比0.01~0.027頻段形成的腦網絡更穩定。上述研究均強調了fMRI信號低頻成分的功能連接和網絡特性, 但是在頻率邊界的界定上未能形成統一, 為后續的驗證性和發現性研究設置了一定障礙。此外, 上述研究中采用的分頻方法如傅里葉變換、切比雪夫濾波器等多假設信號線性平穩, fMRI信號并不能滿足這一點, 這也限制了對其結果的解釋和推廣, 沒有充分揭示fMRI信號的完整特性。
本文采用了一種數據驅動的多維集合經驗模式算法對fMRI信號進行分頻, 相比傳統分頻方法, 該算法以信號自身特性為出發點, 從時間維度和空間維度共同提取信號有效信息, 可以更加準確刻畫出信號的物理過程。為展示這一新方法的應用, 我們針對“性別差異”這一生命科學, 特別是心理學, 領域中長期以來感興趣的科學問題, 應用上述多尺度自適應的分頻方法探究了大腦低頻成分功能連接的性別差異, 嘗試為該算法在腦功能領域的應用提供參考。
2 材料與方法
2.1 數據采集和預處理
本研究招募了66名健康右利手被試, 年齡范圍18~35歲, 其中男性37例, 女性29例, 均無高血壓、心臟病、腦血管病和神經精神類疾病, 均簽署知情同意書。所有腦影像掃描均在中國科學院心理研究所磁共振中心完成, 磁共振掃描儀為GE (3T)MR750掃描儀, 掃描前用海綿墊將其頭部固定好, 并告知在靜息態磁共振掃描中保持頭部不動, 注視屏幕中的十字, 不能入睡。人腦解剖形態的T1加權結構磁共振成像參數設置如下:翻轉角為8度; 層厚為1 mm; 層間距為1 mm; 重復時間為6.896 mm; 回波時間為2.992 mm; 視野為256 mm, 采集矩陣為256 × 256。人腦靜息態功能的磁共振成像參數設置如下:翻轉角為10度; 層厚為3.5 mm; 層間距為0.5 mm; 回波時間為30 mm; 重復時間為2000 mm; 視野為220 mm, 采集矩陣為64 × 64。圖像采用中國科學院心理研究所腦功能連接組及發展實驗室研制的連接組計算平臺(Connectome Computation System, CCS) (https:// github.com/zuoxinian/CCS) (Zuo et al., 2013; Xu et al., 2015)進行預處理, 首先結構像使用volbrain腦影像在線處理平臺(http://volbrain.upv.es/)進行圖像去噪, 信號值矯正, 顱骨分割等步驟, 再調用FreeSurfer (Fischl, 2012)的Recon-all程序對大腦皮層進行分割和二維表面重建。靜息態fMRI圖像預處理包括:(1)剔除每次掃描的前5個時間點共10 s的數據。(2)進行野點矯正, 掃描時間矯正、頭動矯正, 其中頭動使用平均幀平移(meanFD)進行量化, meanFD超過0.5 mm的數據將被排除(Jo, Saad, Simmons, Milbury, & Cox, 2010; Power, Schlaggar, & Petersen, 2015)。(3)使用基于灰白質邊界的配準方法將個體的功能像配準至T1加權結構像(FreeSurfer:bbregister) (Greve & Fischl, 2009)。(4)對個體的功能像進行灰質、白質、腦脊液的分割(Fischl et al., 2002)。(5)回歸個體白質和腦脊液平均時間序列、使用Frston24頭動參數模型進行頭動回歸(Yan et al., 2013; Zang et al., 2007)。(6)進行時間序列的帶通濾波(0.01~0.1 Hz)濾波。
2.2 分頻腦網絡的構建
本文采用快速多維集合經驗模式分解(Fast Multi-Dimensional Ensemble Empirical Mode Decomposition, FMEEMD)方法對fMRI信號進行分頻, 不同于傳統的Fourier變換和小波變換方法, 完全以信號自身特征為依據進行分解, 分解過程中會產生不同特征尺度的本征模態函數(IMF)并對其進行Hilbert變換可得信號瞬時頻率等特性, 是對信號的真實物理過程的反映(Wu, Huang, & Chen, 2009; Wu & Huang, 2009)。在求解fMRI信號的頻段時, 對大腦的每個感興趣區域(Region Of Interest, ROI)的BOLD信號分解得到IMF, 再進行Hilbert變換得到瞬時頻率, 同時引入希爾伯特加權頻率(Hilbert weighted frequency, HWF) (Xie & Wang, 2006)來反映IMF的平均振蕩頻率(Song, Zhang, & Liu, 2014)。具體計算方法如下:
(1)對每個IMF進行Hilbert變換如公式(1)所示
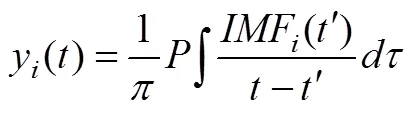
其中P代表柯西準則值。
(2)計算其解析信號:
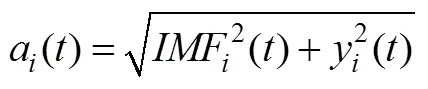
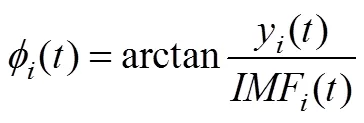
(3) IMF的瞬時頻率計算如公式(5)所示:
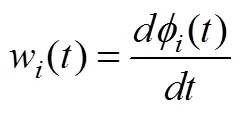
(4)每個IMF的m個數據點的HWF計算:
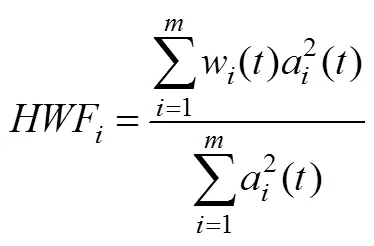
本文計算了每個ROI的IMF1到IMF5的HWF值, 得到整個大腦信號的IMF1到IMF5的頻率分布, 采用了如圖1所示的方法構建不同頻段的腦網絡, 主要過程包括數據預處理、根據腦圖譜提取感興趣區域的時間序列、時間序列分頻處理、計算不同頻段范圍節點之間的關聯矩陣形成腦網絡、計算不同頻段腦網絡全局和局部特征等, 上述算法將隨未來CCS的新版本發布。人腦連接組計劃(Human Connect Project, HCP) (Glasser et al., 2016)提出的最新多模態皮層分區涵蓋了迄今最為全面的多模態腦影像學信息, 基于此我們提取每個ROI內所有體素的平均時間序列作為該ROI的時間序列。HCP皮層分區將全腦皮層數據劃分成360個ROI作為腦節點, 左右半腦各180個。由于功能像采集中顳極、眶額等部分的信號丟失, 最終剩余354個腦區。對每個被試的354個腦區的時間序列采用如上的FMEEMD方法進行分頻處理, 得到4個頻段的ROI的時間序列, 然后在各個頻段下計算每對ROI時間序列之間的皮爾遜相關系數, 構建腦功能網絡。
注:彩圖見電子版, 下同
2.3 統計檢驗
將每個頻段的功能連接矩陣通過fisher Z變換轉換為Z值, 進行雙樣本(男女) t檢驗, 使用FDR矯正進行ROI級別的多重比較校正, 校正后的統計性顯著性水平為0.05。為便于討論研究發現, 最后再將功能連接矩陣涉及的HCP分區聯系到Yeo的7個腦功能網絡(Yeo et al., 2011), 即根據一個HCP的分區所占腦功能網絡的比例來決定它的網絡歸屬, 這樣就可以結合腦區和網絡來共同探討不同頻段的大腦功能連接的性別差異。
3 結果
3.1 不同頻段的腦網絡構建結果
采用FMEEMD方法對所有被試的數據分頻范圍顯示頻率范圍0.01~0.10 Hz, 詳細頻率范圍分布如圖2所示。其中IMF1的頻段范圍為0.06~ 0.10 Hz, IMF2范圍為0.036~0.06 Hz, IMF3范圍為0.025~0.036 Hz, IMF4范圍為0.01~0.025 Hz。
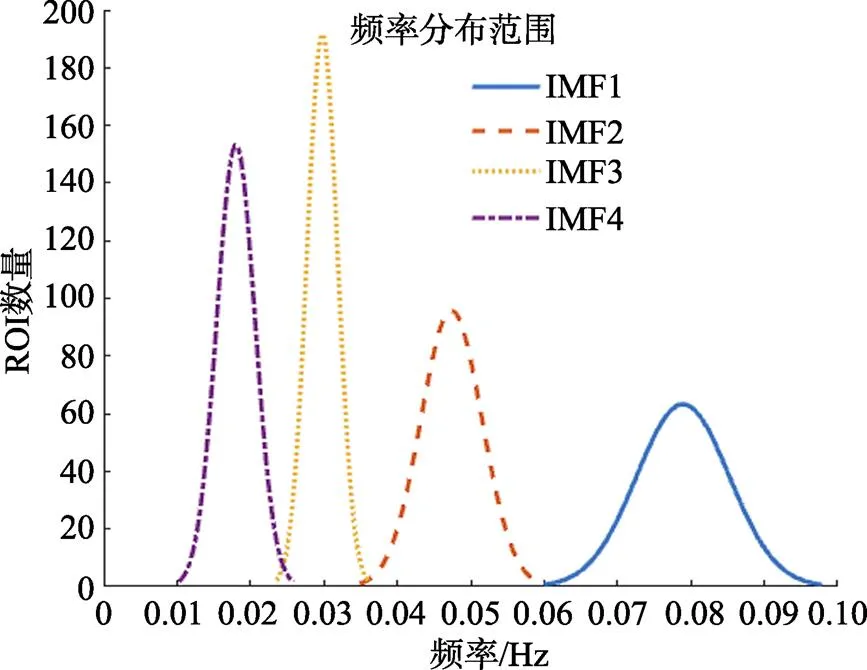
圖2 頻率范圍分布
圖3展示了4個頻段上的組水平平均功能連接矩陣, 按照Yeo的7網絡分區將所有腦區重新排序, 矩陣從左至右(從上至下)依次為視覺網絡、感覺運動網絡、背側注意網絡、腹側注意網絡、邊緣網絡、額頂控制網絡和默認網絡內腦區與其他腦區的組水平平均相關系數(z值)。研究結果顯示:隨著頻率段降低, 網絡間的功能連接逐步以負連接為主, 而網絡內部區域之間也逐步出現較多的負連接, 七大皮層功能網絡間模塊化逐漸減弱, 在IMF1 (0.06~0.10 Hz)內可發現7個網絡內的功能連接明顯強于網絡間的功能連接, 其中默認網絡的網絡間功能連接, 特別是與邊緣網絡之間的功能連接強于其他網絡, 在該頻段內, 網絡間的長程連接數量和強度均多于其他頻段; 在IMF2 (0.036~0.06 Hz)內, 全腦功能連接均有增強, 同時邊緣網絡、額頂控制網絡和默認網絡之間的網絡間功能連接強度上升, 該三網絡間的區分度下降; 在IMF3 (0.025~0.036 Hz)內, 網絡間的區分度進一步下降, 只有視覺網絡、感覺運動網絡、背側注意網絡、腹側注意網絡保持了較好的網絡間區分度, 同時網絡內的功能連接矩陣也逐漸出現分化, 提示在各網絡內也存在著具有不同連接特性的子模塊; 在IMF4 (0.01~0.025 Hz)內, 只有視覺網絡和感覺運動網絡保持了較好的網絡完整性, 默認網絡保持了部分子模塊特性, 其他網絡則已經很難區分。
3.2 男女差異的統計檢驗結果
磁共振成像掃描期間, 男女未表現出顯著的頭動差異(= ?0.58,= 0.57), 因此未將頭動作為影響因素考慮在功能連接比較中。將不同頻段下的功能連接矩陣經Fisher Z轉換后做獨立樣本t檢驗檢驗性別差異, 發現僅在IMF1 (0.06~0.10 Hz)頻段上功能連接在性別間有顯著的差異(校正后), 其他頻段并無顯著性差異。精確顯示差異的腦區位置可在HCP腦區ROI分布中定位, 為L_FOP1_ ROI和R_V3CD_ROI, L_6ma_ROI和L_10pp_ROI, L_6ma_ROI與 R_10pp_ROI, L_8BM_ROI與 R_8C_ROI。上述腦區詳細位置信息和所屬網絡如表1所示, 性別差異數據信息如表2所示, 其中p值經FDR校正(< 0.05, 校正后)。研究結果顯示男性較女性組的功能連接較強的腦區在左側6ma和左右側10pp區域, 而在左側FOP1和V3CD、8BM和8C區域間的功能連接在女性中強于男性。進一步從網絡視角發現, 男性大于女性的功能連接存在于腹側注意網絡和邊緣網絡的網絡間功能連接, 女性比男性較強的功能連接則在腹側注意網絡和視覺網絡、額頂控制網絡的網絡內功能連接, 在其他頻段并未發現顯著差異。
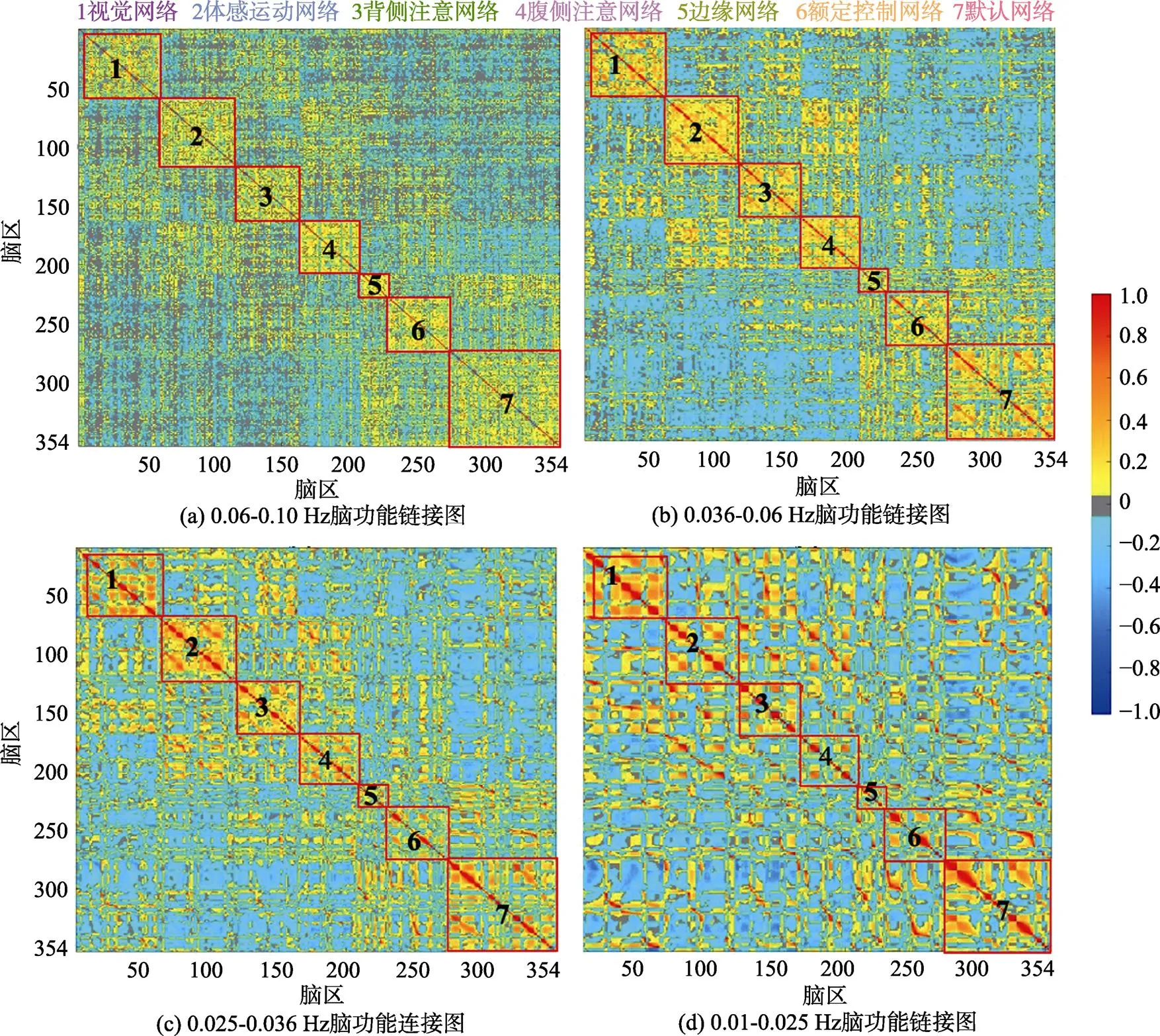
圖3 不同頻段的功能連接圖

表1 腦區分布信息表
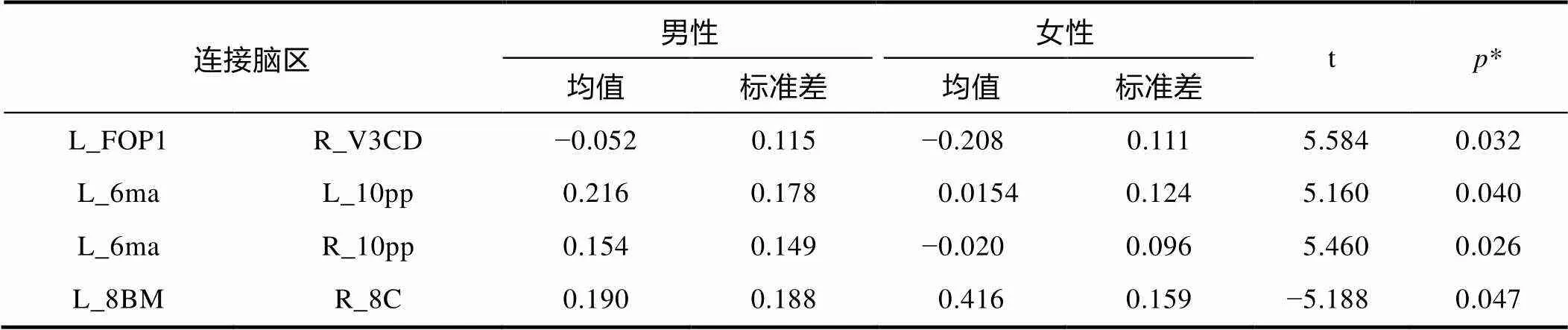
表2 功能連接性別差異結果
注:*經過FDR矯正
4 討論
本研究首先使用FMEEMD方法對所有被試的功能磁共振數據進行了頻率劃分, 根據已有研究表明, fMRI的高頻信號可能和頭動、血流等因素有關, 同時不同預處理流程對分頻結果產生較大影響, 由于本研究的主要目的并不在于使用FMEEMD分頻方法區分fMRI原始信號中的噪音成分, 而是探究在不同頻率段內fMRI信號可能具有的網絡特性和男女差異, 因此使用了通常的數據預處理流程, 同時重點關注已有發現的具有較強生理意義的0.01~0.1 Hz低頻段的信號。分頻結果發現高頻信號內具有較好的腦網絡模塊化, 同時網絡間的功能連接較強, 高級認知皮層的模塊化程度最強。而隨著頻率下降, 網絡模塊化趨勢減弱, 但是局部連通性和網絡內子模塊化特性逐漸凸顯, 最低頻頻段內, 只保留了初級感知覺皮層如視覺網絡和感覺運動網絡的模塊化, 且網絡內的子模塊性最強, 如默認網絡內的部分區域間功能連接在該頻段內強度較高, 提示在該低頻頻段內的功能連接, 更傾向于反映初級感知覺皮層以及局部大腦區域間的功能連通性。因此, 我們認為, 不同頻段內的大腦功能磁共振信號具有不同的功能特性, 這種功能特性可能對應著大腦認知功能的層級性。例如, 在高頻成分內的較強的網絡間連接可能反映的是不同網絡間的信息傳遞性較強, 對應著高級認知功能協調不同大腦網絡協同工作的特征; 隨著頻率降低, 皮層七大網絡間模塊化減弱, 可能反映在不同頻段下人腦的模塊化信息處理的尺度差別, 信息主要是相同認知功能的成分間傳遞; 而在最低頻段內僅保留初級感知覺皮層的模塊化, 而網絡內不同成分間的連通性都已經較低, 可能預示著在該頻段內信息只在負責特定具體功能的區域間具有較好的連通性, 而負責高級認知功能的網絡模式已很難觀察到。因此隨著頻段由高至低, 可能對應著大腦功能從高級認知控制(協調負責不同功能的腦區間活動)到復雜認知功能實現(實現特定認知能力)再到單一認知能力(初級感知覺加工)的功能層級性。在此基礎上, 我們進一步分析了不同頻段下功能連接的性別差異, 發現只在IMF1頻段出現了差異, 且多為網絡間的長程連接差異, 網絡內功能連接差異體現在左右腦間的額頂控制網絡內, 一定程度上也佐證了上述分頻結果, 即IMF1較好保留了大腦網絡特性, 能反映網絡間的長程功能連接。
FOP1和V3CD區域間存在負向功能連接, 且具有顯著的男女差異。該連接強度在男性中較弱, 女性中較強, 說明男性中該兩區域間的功能關聯性較低, 而在女性中存在反向的功能同步性。FOP1比周圍區域的髓鞘化更強, 一般在進行運動相關任務時激活。該區域從屬于腹側注意網絡, 主要負責自下而上的認知加工, 接收初級皮層的感覺信息輸入并對其進行反應(Eckert et al., 2009)。V3CD區域屬于枕外側顳葉皮質, 毗鄰視覺聯絡區V4 (Abdollahi et al., 2014; Malikovic et al., 2016), 有更多的髓磷脂和較少的皮層厚度, 負責對復雜的視覺的加工。我們認為, 女性在FOP1和V3CD間的反向功能同步性可能預示著在女性中處理復雜視覺信息的腦區與注意相關的腦區活動間存在一定的競爭關系, 而在男性中這兩個區域間的關聯性很低, 說明可能僅在女性中存在支持腦功能活動的競爭關系(Sorge & Totsch, 2017; Herlitz & Rehnman, 2008)。
在6ma區域和左右側10pp區域間的功能連接在男性中顯著強于女性, 6ma區域屬于輔助運動區(SMA)的一部分, 在人體中SMA的神經元直接投射到脊髓上, 主要控制運動的內部生成規劃、運動序列的規劃和身體兩側的協調, 起到直接控制運動的作用, 如攀爬和跳躍等具體行為(Serrien, Strens, Oliviero, & Brown, 2002; 張琪涵等, 2014)。該區域也屬于腹側注意網絡, 參與自下而上的認知調控。10pp區域位于眶額和額極皮質區域, 在HCP分區中該區域被劃分為11個子區域, 10pp是其中一個, 眶額區域被認為是參與許多高級的認知工作, 比如未來行動的規劃和組織, 以及類比的能力等, 是基于抽象的規劃、情節記憶和短時記憶信息的基礎(Bludau et al., 2014), 但是相較于眶額部其他區域, 10pp區在工作記憶、語言故事等任務中表現出了去激活, 僅在運動任務中表現出了一定的激活, 我們推測10pp區域可能在眶額皮層區域負責協調信息傳入和認知規劃的作用(van Essen, Glasser, Dierker, Harwell, & Coalson , 2012)。我們認為, 6ma和雙側10pp區域間的功能連接在男性中強于女性, 與上述FOP1和V3CD間的功能連接結果相呼應V3CD和FOP1間的功能連接是基本感知覺信息上傳至高級認知控制區域的中轉, 是信息開始進入腹側注意系統自下而上加工的開始, 并且FOP1主要受運動相關任務激活, 而6ma區域則恰好是屬于輔助運動區, 它和左右側10pp區域的功能連接很可能是腹側注意系統加工后的信息進入高級認知控制的通路, 之后再由眶額部分其他區域進行運動規劃。該通路體現了從刺激輸入, 到引起注意分配, 再到高級皮層形成運動規劃的自下而上加工的完整信息加工通路。而該通路的連通性在女性中弱于男性, 暗示男性在接收外界刺激信息, 并由此作出相關反應的功能通路連通性強于女性, 也符合日常經驗, 即男性更擅長、可以更迅速對外界輸入的視覺信息進行組織并作出應對。
相比于男性, 女性在左側8BM和右側8C區域之間的功能連接更強。8BM和8C區域都屬于額頂控制網絡, 8BM區域主要位于前扣帶回和內側前額葉皮質的頂后側。該區域在結構上髓鞘化程度較強, 在工作記憶任務中激活, 在數學問題背景中的激活強于聽覺理解, 并且具有偏側化趨勢, 如在關聯任務強于匹配任務的激活中, 右側8BM的激活區域更廣泛。8C區域主要位于背外側前額葉皮質, 背外側前額葉皮質是人類大腦皮層中負責高級認知功能的皮層, 主要負責執行和認知功能, 包括工作記憶和認知靈活性(Barbey, Koenigs, & Grafman, 2013)等, 是實施認知控制的最重要腦區之一。8C區域在結構上髓鞘化程度較低, 在工作記憶任務上局部激活較強, 我們推測, 左側8BM和右側8C之間的功能連接, 特別是半球間連接, 體現了大腦在高級認知成分, 特別是在工作記憶和數字加工上的腦區間協調及整合性上, 女性要強于男性。
綜上所述, 我們認為FMEEMD方法可以有效地提取大腦功能磁共振時間信號的頻率特性, 同時, 基于FMEEMD方法所得到的男女功能連接差異表明, 男性在由下至上的加工中具有較強的功能連通性, 而女性則在工作記憶相關的高級腦區間表現出了較強的半球間整合性。本研究具有一定的局限性, 例如樣本量較少, 影像學結果無法通過多重比較校正, 可能會導致未在其他頻段發現性別差異; 同時本研究結果基于靜息態功能磁共振數據, 缺少行為數據和相關實驗結果的佐證, 未能充分討論腦影像結果在行為中的表現; 此外在方法學上, 我們使用了較為保守傳統的數據預處理方式, 可能會導致一定的假陰性結果, 未來可以使用FMEEMD方法對原始高質量(如multiband)序列采集的功能磁共振數據進行分析, 探討該方法在數據預處理上的應用前景。
5 結束語
本文采用FMEEMD分解方法有效避免了經驗驅動方式中需要提前定義頻段的缺陷。對大腦的靜息態fMRI低頻成分(< 0.1 Hz)進行分頻, 得到了4個頻率段范圍, 從高到低依次為0.06~0.10 Hz, 0.036~0.06 Hz, 0.025~0.036 Hz和0.01~0.025 Hz, 并構建了4個頻段的大腦功能網絡, 討論了網絡內和網絡間的功能連接屬性, 隨頻率提高, 功能連接矩陣中的網絡化程度上升, 網絡內功能連接下降, 網絡間功能連接增強。在此基礎上, 我們進一步比較了不同頻段內功能連接的性別差異, 結果顯示在0.06~0.10 Hz頻段上, 男性比女性較強的功能連接分別集中在腹側注意網絡和邊緣網絡間, 體現了自下而上的加工方式; 女性比男性較強在腹側注意網絡和視覺網絡、以及額頂控制網絡內的功能連接, 體現了高級皮層在認知功能上的協調和整合性。本研究展示了FMEEMD分頻算法在功能磁共振數據上的應用, 為男女差異提供了基于影像的人腦功能連接組學證據。
張琪涵, 劉穎, 周菘, 章鵬, 彭國慧, 宋星, 白學軍. (2014). 主運動區與輔助運動區在運動執行與運動想象任務中的作用:一個近紅外光譜技術的研究. 見. 北京: 中國心理學會.
Abdollahi, R. O., Kolster, H., Glasser, M. F., Robinson, E. C., Coalson, T. S., Dierker, D., Orban, G. A. (2014). Correspondences between retinotopic areas and myelin maps in human visual cortex., 509–524.
Achard, S., Salvador, R., Whitcher, B., Suckling, J., & Bullmore, E. (2006). A resilient, low-frequency, small-world human brain functional network with highly connected association cortical hubs., 63–72.
Barbey, A. K., Koenigs, M., & Grafman, J. (2013). Dorsolateral prefrontal contributions to human working memory., 1195–1205.
Bludau, S., Eickhoff, S. B., Mohlberg, H., Caspers, S., Laird, A. R., Fox, P. T., Amunts, K. (2014). Cytoarchitecture, probability maps and functions of the human frontal pole., 260–275.
Eckert, M. A., Menon, V., Walczak, A., Ahlstrom, J., Denslow, S., Horwitz, A., & Dubno, J. R. (2009). At the heart of the ventral attention system: The right anterior insula., 2530–2541.
Fischl, B. (2012). FreeSurfer., 774–781.
Fischl, B., Salat, D. H., Busa, E., Albert, M., Dieterich, M., Haselgrove, C., Dale, A. M. (2002). Whole brain segmentation: Automated labeling of neuroanatomical structures in the human brain., 341–355.
Glasser, M. F., Coalson, T. S., Robinson, E. C., Hacker, C. D., Harwell, J., Yacoub, E., van Essen, D. C. (2016). A multi- modal parcellation of human cerebral cortex., 171–178.
Greve, D. N., & Fischl, B. (2009). Accurate and robust brain image alignment using boundary-based registration., 63–72.
Herlitz, A., & Rehnman, J. (2008). Sex differences in episodic memory., 52–56.
Jo, H. J., Saad, Z. S., Simmons, W. K., Milbury, L. A., & Cox, R. W. (2010). Mapping sources of correlation in resting state fMRI,with artifact detection and removal., 571–582.
Liang, X., Wang, J. H., Yan, C. G., Shu, N., Xu, K., Gong, G. L., & He, Y. (2012). Effects of different correlation metrics and preprocessing factors on small-world brain functional networks: A resting-state functional MRI study., e32766.
Malikovic, A., Amunts, K., Schleicher, A., Mohlberg, H., Kujovic, M., Palomero-Gallagher, N., Zilles, K. (2016). Cytoarchitecture of the human lateral occipital cortex: Mapping of two extrastriate areas hOc4la and hOc4lp., 1877–1897.
Power, J. D., Schlaggar, B. L., & Petersen, S. E. (2015). Recent progress and outstanding issues in motion correction in resting state fMRI., 536-551.
Salvador, R., Martínez, A., Pomarol-Clotet, E., Gomar, J., Vila, F., Sarró, S., Bullmore, E. (2008). A simple view of the brain through a frequency-specific functional connectivity measure., 279–289.
Salvador, R., Suckling, J., Schwarzbauer, C., & Bullmore, E. (2005). Undirected graphs of frequency-dependent functional connectivity in whole brain networks., 937–946.
Serrien, D. J., Strens, L. H. A., Oliviero, A., & Brown, P. (2002). Repetitive transcranial magnetic stimulation of the supplementary motor area (SMA) degrades bimanual movement control in humans., 89–92.
Song, X. P., Zhang, Y., & Liu, Y. J. (2014). Frequency specificity of regional homogeneity in the resting-state human brain., e86818.
Sorge, R. E., & Totsch, S. K. (2017). Sex differences in pain., 1271–1281.
Van Essen, D. C., Glasser, M. F., Dierker, D. L., Harwell, J., & Coalson, T. (2012). Parcellations and hemispheric asymmetries of human cerebral cortex analyzed on surface-based atlases., 2241–2262.
Wu, C. W., Gu, H., Lu, H. B., Stein, E. A., Chen, J. H., & Yang, Y. H. (2008). Frequency specificity of functional connectivity in brain networks., 1047–1055.
Wu, Z. H., & Huang, N. E. (2009). Ensemble empirical mode decomposition: A noise-assisted data analysis method., 1–41.
Wu, Z. H., Huang, N. E., & Chen, X. Y. (2009). The multi- dimensional ensemble empirical mode decomposition method., 339–372.
Xie, H. B., & Wang, Z. Z. (2006). Mean frequency derived via Hilbert-Huang transform with application to fatigue EMG signal analysis., 114–120.
Xu, T., Yang, Z.,Jiang, L. L., Xing, X. X., Zuo, X. N. (2015). A connectome computation system for discovery science of brain., 86-95.
Yan, C. G., Cheung, B., Kelly, C., Colcombe, S., Craddock, R. C., Di Martino, A., Milham, M. P. (2013). A comprehensive assessment of regional variation in the impact of head micromovements on functional connectomics., 183–201.
Yeo, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D., Hollinshead, M., Buckner, R. L. (2011). The organization of the human cerebral cortex estimated by intrinsic functional connectivity., 1125–1165.
Zang, Y. F., He, Y., Zhu, C. Z., Cao, Q. J., Sui, M. Q., Liang, M., Wang, Y. F. (2007). Altered baseline brain activity in children with ADHD revealed by resting-state functional MRI., 83–91.
Zuo, X. N., Di Martino, A., Kelly, C., Shehzad, Z. E., Gee, D. G., Klein, D. F., Milham, M. P. (2010). The oscillating brain: Complex and reliable., 1432–1445.
Zuo, X. N., Xu, T., Jiang, L. L., Yang, Z., Cao, X. Y., He, Y., Milham, M. P. (2013). Toward reliable characterization of functional homogeneity in the human brain: Preprocessing, scan duration, imaging resolution and computational space., 374–386.
Sex differences in adaptive multi-scale functional connectivity of the human brain
DUAN Kaikai1; DONG HaoMing2,3,4,5; MIAO Liwen1; SU Xuequan6,7; XIANG Jie1; ZUO XiNian2,3,4,5,7
(1College of Computer Science and Technology, Taiyuan University of Technology, Taiyuan 030024, China) (2CAS Key Laboratory of Behavioral Science, Institute of Psychology, Beijing 100101, China) (3Department of Psychology, University of Chinese Academy of Sciences, Beijing 100049, China)(4Magnetic Resonance Imaging Research Center, Institute of Psychology, Chinese Academy of Sciences, Beijing 100101, China) (5Research Center for Lifespan Development of Mind and Brain, Institute of Psychology, Chinese Academy of Sciences, Beijing 100101, China) (6Institute of Physical Education, Guangxi Teachers Education University, Nanning 530000, China) (7Key Laboratory of Brain and Education, Guangxi Teachers Education University, Nanning 530001, China)
Recent advances on functional magnetic resonance imaging (fMRI) demonstrated sex differences in the brain function. However, no standard on fMRI signal’s frequency division limited further biologically plausible explanation of these observations. In this work, we proposed a fast-multi-dimensional ensemble empirical mode decomposition to extract their multi-scale features of fMRI signal. We found that: this method can perform adaptive frequency allocation for the resting-state fMRI signal, whereby the built multi-scale function network in the frequency brain of 0.06 ~ 0.10 Hz showed significant sex differences regarding its connectivity; males had strong functional connectivity primarily within the limbic network and ventral attention network whereas females presented their strong functional connectivity mainly related to the visual network, ventral attention network and frontoparietal control network. These findings present a new method for the analysis of functional MRI images and provided brain imaging evidence on sex differences in functional connectomics.
sex differences; functional connectivity; brain network; functional magnetic resonance imaging; low frequency characteristics
2018–01–22
* 國家自然科學基金(61503272, 61305142, 61373101)、山西省自然科學青年基金(2015021090, 201601D202042)、山西省重點研發計劃項目(201603D111014)、博士后基金(2016M601287)、虛擬現實技術與系統國家重點實驗室開放課題(BUAA-VR-15KF-16)和山西省回國留學人員科研資助項目(No.2016-037)資助。
左西年, E-mail: zuoxn@psych.ac.cn; 相潔, E-mail: xiangjie@tyut.edu.cn
B845
10.3724/SP.J.1042.2018.01567
猜你喜歡
中華詩詞(2022年6期)2022-12-31 06:41:24
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
電子制作(2018年11期)2018-08-04 03:25:42
中國科技論壇(2017年7期)2017-07-25 08:49:53
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
電測與儀表(2015年5期)2015-04-09 11:30:52
中國中醫藥現代遠程教育(2014年22期)2014-03-01 04:32:55
