自噬在骨保護素對破骨細胞及其前體凋亡過程的調控機制
2018-09-01 08:09:14劉慶羊孫自強趙鴻雁宋瑞龍顧建紅劉宗平
畜牧獸醫學報 2018年8期
劉慶羊,孫自強,趙鴻雁,宋瑞龍,顧建紅,劉宗平*
(1.揚州大學獸醫學院,揚州 225009; 2. 揚州大學比較醫學研究院,揚州 225009;3.江蘇高校動物重要疫病與人獸共患病防控協同創新中心,揚州 225009; 4. 揚州大學教育部農業與農產品安全國際合作聯合實驗室,揚州 225009)
破骨細胞(osteoclasts,OCs)是具有骨吸收功能的多核巨細胞。它不僅參與生理性骨改建,而且在病理性骨吸收過程中也具有重要作用。當OCs數量增加或活性增強,可導致骨質疏松、炎癥性關節炎等代謝性骨病。骨保護素(osteoprotegerin,OPG)是由成骨細胞分泌,能抑制OCs的分化和OCs的活性[1-2]。研究發現,破骨細胞的骨吸收與自噬密切相關,DeSelm等[3]證明自噬相關蛋白Atg5、LC3等通過影響破骨細胞皺褶緣的形成和炎性物質的分泌調控OCs的骨骼重吸收功能。但OPG對OCs自噬的作用機制尚未闡明。本實驗室前期研究發現,(0、20、40、80 ng·mL-1) OPG處理RAW264.7誘導的OCs 24 h后,OPG呈劑量依賴性地抑制OCs的形成和分化[4],也有文獻報道,OPG可競爭性地結合腫瘤壞死因子(TNF) 相關凋亡誘導配體(TNF-related apoptosis inducing ligand,TRAIL),抑制TRAIL誘導的細胞凋亡[5-6]。研究表明,凋亡和自噬在不同的情況下存在密切和復雜的關系[7]。饑餓、低氧、感染和氧化應激等異常情況可誘導細胞發生死亡[8]。自噬一般被認為是細胞的自我保護機制,但自噬過度激活也會導致細胞死亡[9]。在OPG作用于OCs和 OCPs后,自噬對凋亡的調控機制尚不明確。本文以小鼠單核巨噬細胞系RAW264.7細胞誘導分化形成OCs和OCPs,研究OPG對OCs、OCPs自噬和凋亡的影響及自噬對細胞凋亡的調控作用。
1 材料與方法
1.1 細胞株
RAW264.7細胞株,購自中國科學院上海生命科學研究院生物化學與細胞生物學研究所細胞庫(ATCC Number:TIB-71)。
1.2 主要儀器與試劑
二氧化碳培養箱、超低溫冰箱(Thermo,USA);DMI3000B倒置相差顯微鏡(Leica,Germany);電泳儀(Bio-Rad,USA);5810R型冷凍離心機(Eppendorf,Germany);CyAn ADP7型流式細胞儀(Beckman Coulter,USA);DMEM、α-MEM (Life Technologies,USA);10%胎牛血清(Gibco,USA),M-CSF、RANKL、OPG(Peprotech,USA);氯喹(MedChemExpress,USA);雷帕霉素(Sigma,USA);兔抗L3B(Sigma,#L7543,USA),兔抗Atg5(Cell Signaling,#12994s,USA),兔抗Beclin-1(Cell Signaling,#3495s,USA),兔抗β-actin(Cell Signaling,#4970L,USA);Annexin V-FITC 熒光染色試劑盒(BD,USA)。
1.3 RAW264.7 細胞培養與處理
將液氮凍存的RAW264.7 細胞37 ℃水浴融化轉移至15 mL離心管,加入7 mL DMEM培養液室溫1 000 r·min-1離心5 min后棄上清,加入5 mL培養液輕輕重懸,轉移至細胞培養瓶,置于37 ℃、 5% CO2飽和濕度的恒溫培養箱中培養10 h換液,繼續培養待RAW264.7細胞鋪滿細胞培養瓶底80%~90% 時傳代。RAW264.7培養至第5代,細胞重懸于α-MEM培養液(含10%FBS),將細胞以2×104·mL-1的濃度接種到6孔板培養,培養基中加入細胞因子M-CSF(25 ng·mL-1)和RANKL(30 ng·mL-1),培養3~5 d,每2 d 更換培養液一次,并添加同樣濃度的細胞因子。
RAW264.7細胞誘導3或5 d后分化成OCPs或OCs,經CQ(50 μmol·L-1) 預處理30 min,或RAP(5 μmol·L-1) 預處理1 h,添加OPG(80 ng·mL-1)作用細胞6 h,試驗分組:Con、OPG、CQ、CQ+OPG、RAP和RAP+OPG組。
1.4 Western blot檢測OCs和OCPs自噬相關蛋白的表達
取處理過的細胞,棄去培養基,4 ℃PBS清洗2次 (去除不貼壁的細胞),加入含有蛋白酶抑制劑的RIPA裂解液,置于冰上裂解30 min,收集裂解液,于超聲裂解20 s,冷凍離心機4 ℃ 1 200 r·min-1離心10 min,收集上清液。BCA蛋白濃度定量試劑盒測定各組蛋白濃度,并調整為一致濃度,按體積加入6× SDS-PAGE Loading Buffer(1∶5),隔水煮沸10 min ,-80 ℃冰箱儲存備用。
取等量樣品加入凝膠泳道,初始階段恒壓90 V電泳,待蛋白抵達分離膠恒壓110 V電泳,電泳結束后,采用恒壓濕轉法將蛋白從SDS-PAGE膠轉移至PVDF膜,轉膜條件110 V恒壓90 min,(轉膜時間依據目的蛋白分子量的不同需適當調整),根據目的蛋白相對分子質量大小剪裁PVDF膜,用含5%脫脂牛奶的TBST封閉緩沖液室溫振蕩封閉2 h,加入含 5% BSA的TBST配制的一抗(1∶1 000),4 ℃ 孵育過夜,回收一抗,TBST緩沖液洗滌PVDF膜6次(5 min·次-1),辣根過氧化物酶HRP標記的二抗(1∶5 000)室溫孵育2 h,回收二抗,TBST緩沖液洗滌PVDF膜6次(5 min·次-1),將PVDF膜與化學發光底物檢測試劑混合液充分反應,然后將膜置于化學發光圖像分析儀內,拍攝照片。用Image Lab軟件計算灰度值。
1.5 Annexin V-FITC/PI雙標記法檢測OPG對OCs和OCPs凋亡率的影響
參照Annexin V-PI雙染色試劑盒的說明進行細胞處理。收集上清,用0.25% 胰蛋白酶消化收集細胞,1 200 r·min-1,4 ℃離心5 min ,棄上清,加入1 mL PBS重懸細胞,1 200 r·min-1,4 ℃ 離心5 min ,棄上清,加100 μL Binding Buffer制備細胞懸液,再加入5 μL Annexin V-FITC和5 μL PI混勻,室溫避光反應20~30 min,每5 min輕輕顛倒混勻,加入400 μL Binding Buffer緩沖液終止反應,細胞懸液經200目篩網過濾分裝到流式管,立即用流式細胞儀檢測。
1.6 數據分析
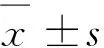
2 結 果
2.1 OPG對OCs及OCPs自噬的影響
2.1.1 OPG對OCPs自噬水平的影響 RAW264.7細胞誘導3 d后,經CQ或RAP預處理后,添加OPG(80 ng·mL-1)作用細胞6 h。Western blot結果顯示(圖1),與Con相比,OPG組OCPs的Atg5和Beclin-1蛋白水平極顯著或顯著上調(P<0.01或P<0.05),LC3-Ⅱ無顯著差異(P>0.05) ,CQ組和CQ+OPG組的LC3-Ⅱ、Atg5和Beclin-1蛋白水平極顯著或顯著上調(P<0.01 或P<0.05),RAP和RAP+OPG組的LC3-Ⅱ、Atg5和Beclin-1水平極顯著上調(P<0.01);與OPG組相比,CQ+OPG組的LC3-Ⅱ和Atg5水平極顯著上調(P<0.01),RAP+OPG組的LC3-Ⅱ水平極顯著上調(P<0.01)。提示OPG處理促進OCPs發生自噬。
2.1.2 OPG對OCs自噬水平的影響 RAW264.7細胞誘導5 d后,經CQ或RAP預處理后,添加OPG(80 ng·mL-1)作用細胞6 h。Western blot結果顯示(圖2),與Con相比,OPG組OCs的 LC3-Ⅱ、Atg5和Beclin-1蛋白水平極顯著下調(P<0.01),CQ和CQ+OPG組的LC3-Ⅱ蛋白極顯著上調(P<0.01),Atg5和Beclin-1蛋白極顯著下調(P<0.01),RAP和RAP+OPG組Atg5和Beclin-1蛋白極顯著下調(P<0.01),RAP組LC3-Ⅱ蛋白極顯著上調(P<0.01),RAP+OPG組LC3-Ⅱ無顯著差異(P>0.05);與OPG組相比,CQ+OPG組的LC3-Ⅱ、Atg5和Beclin-1蛋白極顯著上調(P<0.01), RAP+OPG組的LC3-Ⅱ顯著上調(P<0.05), Atg5和Beclin-1蛋白極顯著下調(P<0.01)。 提示OPG抑制了OCs的自噬產生。
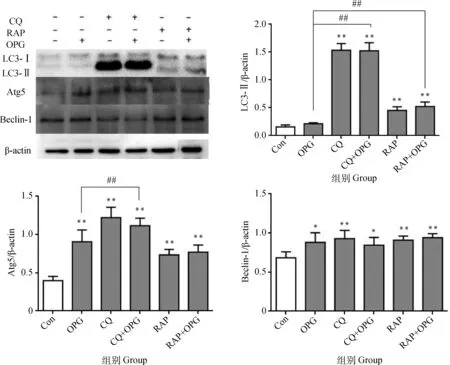
與對照組相比, * *.P<0.01,*.P<0.05;與OPG組相比,## .P<0.01* *. P<0.01, *. P<0.05 versus control; ##.P<0.01 versus OPG圖1 OPG對OCPs自噬水平的影響Fig.1 Effects of OPG on the autophagy in osteoclast precursors
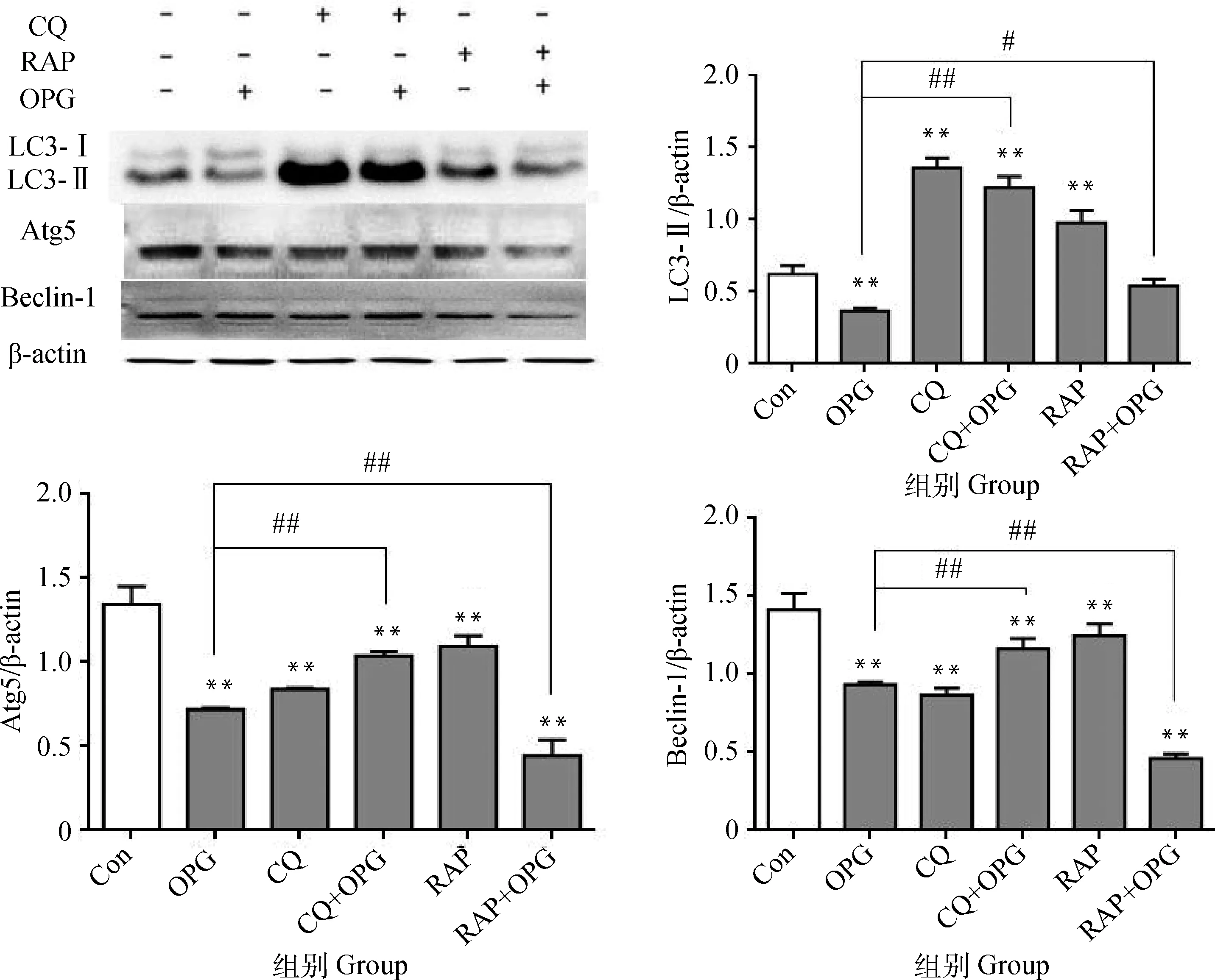
與對照組相比, **.P<0.01;與OPG組相比,## .P<0.01,#.P<0.05**. P<0.01 versus control; ##. P<0.01, #. P<0.05 versus OPG圖2 OPG對OCs自噬水平的影響Fig.2 Effects of OPG on the autophagy in osteoclasts
2.2 OPG對OCs和OCPs凋亡的影響
2.2.1 OPG對OCPs凋亡的影響 RAW264.7細胞誘導3 d后,經CQ或RAP預處理后,添加OPG(80 ng·mL-1)作用細胞6 h。流式細胞儀分析測定結果顯示(圖3),與Con相比,OPG、RAP 和RAP+OPG組早期凋亡率極顯著上升(P<0.01),CQ組和CQ+OPG組早期凋亡率無顯著差異(P>0.05);較OPG組,CQ+OPG組早期凋亡率極顯著降低(P<0.01),RAP+OPG 組早期凋亡率極顯著增強(P<0.01)。提示OPG能夠引起OCPs凋亡,抑制自噬對OCPs凋亡的影響不明顯,而促進自噬能夠導致OCPs凋亡。

與對照組相比, * *.P<0.01,*.P<0.05;與OPG組相比,# # .P<0.01* *. P<0.01, *. P<0.05 versus control; ##.P<0.01 versus OPG圖3 OPG對OCPs凋亡的影響Fig.3 Effects of OPG on the apoptotic ratio of osteoclast precursors
2.2.2 OPG對OCs凋亡的影響 RAW264.7細胞誘導5 d后,經CQ或RAP預處理后,添加OPG(80 ng·mL-1)作用細胞6 h。流式細胞儀分析測定結果顯示(圖4),與Con相比,OPG、CQ、CQ+OPG 、RAP和RAP+OPG組早期凋亡率極顯著下降(P<0.01)。提示OPG處理使OCs凋亡率下降,抑制和促進自噬都能顯著降低OCs的凋亡率。

與對照組相比, * *.P<0.01,*.P<0.05* *. P<0.01, *. P<0.05 versus control圖4 OPG對OCs凋亡的影響Fig.4 Effects of OPG on the apoptotic ratio of osteoclasts
3 討 論
OPG是一種分泌型糖蛋白,能夠抑制骨吸收、增加骨密度,它能夠與細胞核因子-κB 受體活化因子(RANK)競爭性地結合RANK配體(RANKL),并抑制OCs的形成[10]。研究表明,OPG顯著提高小鼠骨髓誘導的OCs的自噬水平[11],也能夠抑制鴨胚OCs分化并促進其凋亡[12]。本文利用小鼠單核巨噬細胞系RAW264.7誘導分化形成OCPs和OCs,添加OPG處理,表明OPG能夠促進OCPs發生自噬和凋亡,抑制OCs發生自噬與凋亡。
自噬是真核細胞中一種自我清除的過程,通過溶酶體介導的降解通路,主要降解細胞內多余或受損的蛋白及細胞器,而自噬過度激活會導致細胞死亡。研究表明,自噬相關基因LC3、Atg5、Beclin-1等在OCs分化過程中起到了重要的作用[13-14]。LC3有LC3-Ⅰ和LC3-Ⅱ兩種亞型,分別定位于前自噬體和自噬體膜表面,LC3-Ⅱ的表達量在某種程度上反映了細胞的自噬活性[15]。Beclin-1在自噬起始階段起到重要的作用,主要通過調節其他的自噬蛋白定位到前自噬體膜上[16]。在動物機體內,OCs的數量受嚴格的控制,以維持骨骼形成和發育,防止骨吸收過度所致的病理性骨丟失[17]。研究發現,RAP影響骨骼生長的功能,減少OCs的數量,破壞長骨的生長發育[18-19]。RAP通過抑制 mTOR的活化促進自噬的發生,添加RAP使OCs活性下降[20-21]。CQ能夠提高溶酶體的pH,使溶酶體中的酸性水解酶失去活性,因此位于自噬體或自噬溶酶體膜上的 LC3不能被及時降解,繼而LC3蛋白表達升高。還有研究表明,CQ通過抑制自噬來阻止OCs的分化,起到治療骨質疏松癥的作用[22]。本實驗室前期研究發現[23],RAW264.7細胞培養1、3、5、7 d后,經80 ng·mL-1的OPG處理24 h,結果顯示OPG對于分化早期的OCPs和分化中末期的OCs具有不同的調控作用,促進OCPs存活和黏附結構形成,抑制OCs活性。本研究在OCs分化至第3、5天,CQ或RAP預處理后,加入80 ng·mL-1OPG處理6 h,發現OPG促進OCPs的自噬水平,而抑制OCs的自噬水平。以上研究結果證明OPG對于OCs分化不同階段的生物效應有所差異,在OCPs的分化過程中自噬起著正向調控作用,促進OCPs的分化;OPG阻止OCs向末期階段分化,抑制OCs的活性,此時自噬主要發揮維持穩態的作用。
細胞凋亡是維持細胞穩態的另一種重要機制。發生凋亡時,細胞膜上的磷脂酰絲氨酸(phosphatidylserine,PS)發生外翻與鈣磷脂結合蛋白結合,Annexin V利用一種對PS具有較高親和性的磷脂結合蛋白作為敏感探針,檢測PS的變化[24]。本課題組前期研究表明[25],40 ng·mL-1OPG處理RAW264.7細胞誘導的OCs,能夠通過抑制OCs的分化過程而阻止細胞凋亡的發生。自噬與凋亡關系在不同的組織和細胞中存在較大差異,順鉑可以引起膀胱癌細胞T24發生自噬,并且促進自噬能促進順鉑誘導的細胞凋亡[26];聯合應用烏蘇酸與唑來膦酸,通過增強自噬與氧化應激水平,促進烏蘇酸誘導的人成骨肉瘤細胞凋亡[27]。另外,饑餓誘導小鼠成骨細胞,前2 h自噬抑制凋亡,隨饑餓時間延長自噬保護作用逐漸降低,最終細胞發生凋亡[28]。本研究采用流式細胞術Annexin V-FITC/PI雙標記法檢測OPG對RAW264.7誘導的OCs和OCPs凋亡率的影響,結果顯示OPG促進OCPs凋亡,抑制OCs凋亡,此外,促進自噬能引起OCPs的凋亡,而抑制和促進自噬皆顯著抑制OCs的凋亡。
4 結 論
OPG促進OCPs發生自噬和凋亡,抑制自噬對OCPs凋亡的影響不明顯,而促進自噬能夠導致OCPs凋亡。此外,OPG抑制了OCs的自噬發生和凋亡,而抑制和促進自噬都能顯著降低OCs的凋亡率。OPG作為骨質代謝、炎癥免疫調節以及細胞凋亡中的重要調節因子,通過干預自噬流對OCs和OCPs凋亡有相反的調控作用,這為骨質疏松、骨硬化癥以及腫瘤等多種疾病的診斷、治療提供了新的思路。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
中學生數理化·八年級物理人教版(2022年3期)2022-03-16 05:55:08
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
當代陜西(2021年2期)2021-03-29 07:41:24
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
人大建設(2019年12期)2019-05-21 02:55:32
中國塑料(2016年3期)2016-06-15 20:30:00
中國火炬(2010年8期)2010-07-25 11:34:30
