糧粒水分擴散方程的由來及研究進展
2018-09-10 20:09:55李興軍
糧食科技與經濟 2018年2期
李興軍


[摘要]糧粒具有吸濕性,它隨周圍環境的變化而解吸或吸收水分。單一糧粒或糧堆的解吸或吸附行為,依賴于糧食種類的物理特征。文章介紹了恒定干燥速率、速率降低干燥階段的概念,以及描述速率降低干燥階段的理論和經驗方程,尤其是水分擴散方程或薄層干燥方程的推導過程,并指出了各種水分擴散方程描述糧粒解吸或吸附速率的優缺點。
[關鍵詞]糧食;吸濕性;解吸;吸附;擴散方程;裂紋
中圖分類號:F307.11 文獻標識碼:A DOI:10.16465/j.gste.cn431252ts.20180220
1恒定干燥速率階段
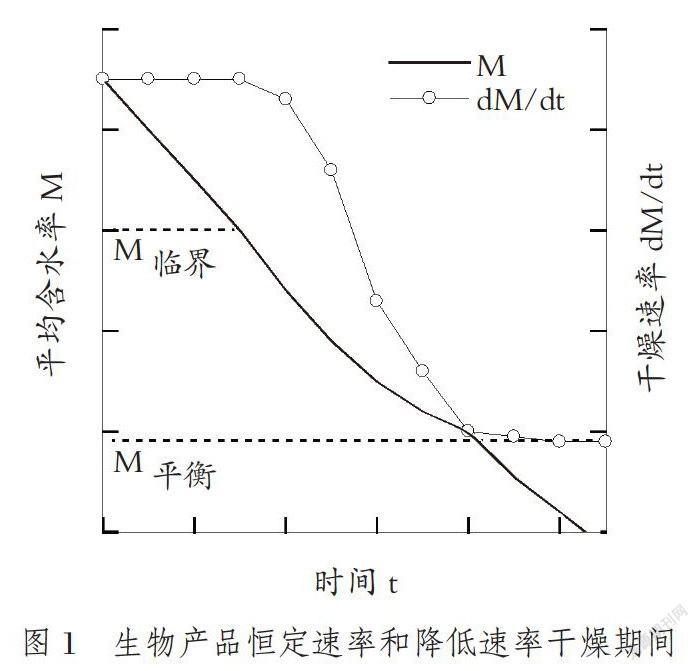
糧食是吸濕的毛細管多孔產品,孔中部分充滿了液態水或空氣/水-蒸汽混合物。當籽粒與周圍空氣之間存在偏水汽壓差時,干燥(解吸)期間,水分從籽粒表面和孔中蒸發離開。干燥同時是熱和水分的轉移過程。包括谷物在內的生物材料,以批量干燥,而不是單一籽粒,批量糧食最初階段顯示恒定速率干燥。含水率高于70%~75%濕基的生物材料最初干燥速率,是空氣流速、溫度及濕度3個干燥參數的函數。在產品表面似乎是薄水層覆蓋。如果周圍條件是恒定的,則干燥速率是恒定的。產品以恒定速率干燥的條件是,水分轉移的內部阻力遠小于從產品表面除去水汽的外部阻力。恒定的干燥速率見圖1。
產品的干燥速率從恒定速率降到降低速率的含水率,叫產品臨界含水率(M臨界)。臨界含水率與產品的固體特征(如形狀和質地)、干燥條件有關。糧食顯示恒定速率干燥期間,則表明在非常不成熟狀態收獲。Simmonds等報道,小麥恒定干燥速率期間對應含水率50%~55%濕基。其他農產品如馬鈴薯、甜菜在恒定周圍條件脫水時最初顯示恒定干燥速率。糧食恒定干燥速率不重要,所以此類研究少。
2干燥速率降低階段
水分蒸發需要熱,水分從產品表面流進干燥介質如空氣中。一些生物產品,在外部恒定條件下以單一顆粒被干燥,在干燥最初階段水分以恒定速率失去,接著是干燥速率降低時期。所謂速率降低干燥階段,是由于水分轉移的內部阻力大于外部阻力,產品不再顯示表面覆蓋的薄層水。產品的含水率降低至低于臨界點(M臨界),產品表面的蒸汽壓強Pv低于vwb,干燥過程的驅動勢能ΔPv減少,這導致干燥速率降低。而且,正在干燥產品內出現水分梯度,產品溫度升高至高于濕球溫度。收獲的糧食僅在降低速率期間完全干燥,在干燥過程中它們的干燥速率持續減少。谷物的干燥和干燥速率曲線見圖2。
在降低速率干燥階段,預測吸濕的毛細管多孔產品的干燥速率,較恒定速率干燥階段更加復雜。分析中既要考慮外部傳遞機制(熱和質量對流傳遞),也要考慮產品內部傳遞機制(熱傳導傳遞和質量擴散傳遞)。已有的研究中,提出的許多理論和方程用于預測速率降低干燥階段谷物糧食的干燥行為,僅半理論和經驗方程被用于糧食干燥設計。速率降低干燥理論用于毛細管多孔產品(砂子、陶瓷)、膠體非多孔產品(肥皂、尼龍)、吸濕毛細管多孔產品(木材、糧食)。
3干燥速率降低方程
3.1理論干燥方程
Luikov模型和Philip-Devries模型解釋了糧食籽粒干燥。解釋吸濕毛細管-多孔非收縮產品如糧食的水分遷移,包括許多物理和熱機制:(1)由于毛細管作用的液體運動;(2)由于水分濃縮差異液體運動(液體擴散);(3)由于滲透力的液體運動;(4)由于重力的液體運動;(5)由于蒸汽濃縮差異液體運動(蒸汽擴散);(6)由于溫度差異的蒸汽運動(熱擴散)。
前蘇聯Luikov等1966年基于物理原理提出的數學模型描述糧食的干燥,假定籽粒不收縮,水分流動是由于總壓強差異。采用兩個穩定態通量方程描述水分和能量傳遞,
從上面討論看出,谷物干燥過程中水分從籽粒解吸,幾乎總是在沒有游離水的干燥速率降低期間發生。水分吸附是這一過程的逆過程。由于產品中細胞吸附和解吸之間的差異,吸附以不同速率發生。解吸速率降低的理論已被徹底修訂。由于干燥過程中傳熱與傳質速率不同,通常分開了描述熱和水分擴散的兩個偶聯偏微分方程,這就產生了薄層干燥方程,這里稱為擴散方程。
與理論方程相比,在應用方面,經驗方程擬合精度更高,而且減少了計算量。指數干燥方程在某些情況下描述薄層干燥,它描述干燥過程最初階段欠佳。指數干燥方程用于糧食儲存更準確,與干燥相比,糧食儲藏具有較多的邊界層阻力,導致儲存情況下空氣運動緩慢甚至停滯。Page方程是干燥的首選經驗方程,比指數干燥方程更準確。
使用擴散方程時,通常認為糧粒是均質的。Walton和Casada發明了基于細胞擴散的方程,這一方程基于葉片材料內部細胞結構的非均質性,對表面水分的有限阻力,解釋了生物材料對水分擴散的阻力不同。Walton等將細胞擴散基于的干燥模型應用于玉米粒的系列干燥數據,發現它比Page方程更適合。Osborn等將這個細胞擴散模型應用于大豆水分吸附,發現Page方程擬合試驗結果,略好于這個細胞擴散模型。
在基于細胞的有限表面阻力擴散模型中,通過實驗確定兩個參數:擴散系數(D9和質量傳遞修正的Biot數目(Bim)。參數D′基本上是內部阻力的倒數,Bim是內部阻力與外部阻力的比率。這些有限表面阻力的模型,截然不同于常用的其他谷物干燥模型,因為它們模擬了內部組織(胚乳和胚)水分向外擴散,并考慮到周圍組織(如麩皮、殼)的外部阻力。修正的Biot數目包括麩皮和殼的阻力,以及表面的對流傳質阻力。
零表面阻力的擴散方程只包含一個參數D′,因為它們根據假設,在均質顆粒內對擴散只有一個組分的阻力(內部)。因此,Bim被認為是無限的,而不是一個待定的參數。被忽略的外部阻力被認為只是對流阻力,這一阻力并不包括細胞內阻力,如糠層的阻力。這種假設在干燥的情況下被接受,因為與內部阻力相比,對流阻力通常是忽略不計的,但如果考慮麩皮和外殼的阻力,它就不是精確的。這些零表面阻力擴散方程,不能解釋對表面存在水分的阻力,由于儲糧情況下存在適當的Biot參數,它們不能解釋內部細胞結構的不均質性。
5展望
解吸期間糧粒內水分運輸被廣泛研究,但是吸附期間糧粒內水分運輸則研究較少,尤其是國內缺乏相應研究。水分吸附環境存在于糧食收獲前的田間,以及隨后的收獲、運輸、干燥及儲藏過程中。水分吸附是稻米在收獲前期和收獲后處理、加工、儲存過程中裂紋產生的一個主要原因。為了更好地了解米粒的裂紋,需要定量分析稻米在水分轉移期間單一籽粒內部應力(stress)/張力(strain)分布,分析米粒在水分吸附或解吸環境下擴散行為。
世界上二分之一的谷物收獲后進入存儲環節,這一環節可發生重大的質量減損。近年來,采用計算機模擬糧食儲存,改善了糧食儲存環節、減少了儲存損失。然而,模擬糧食儲存能力還有限,其原因是對主要糧食種類存儲期間水分吸附速率預測的數據不足。計算機模擬糧食深床干燥和通風時,稻米水分吸附速率可用于計算與暴露時間,以及給定的暴露空氣條件相關的糧食含水率。但只是有限地研究分析了水分吸附速率與平衡相對濕度、暴露相對濕度、暴露時間及稻米類型(稻谷、糙米及精米)之間的關系。
玉米籽粒的裂紋由外部和內部脅迫引起的。裂紋或脅迫裂縫籽粒在處理期間容易破碎,引起貯藏、運輸及加工的問題。籽粒內溫度梯度和水分梯度引起籽粒內不合理的膨脹和收縮,導致內部脅迫的發生。水分梯度對籽粒的膨脹和收縮具有重要影響,而溫度梯度可以忽略。籽粒內發生的脅迫若能準確計算,則設計更好的工藝以減少裂紋發生。這個估算需要研究玉米基本的特性參數如蒸汽擴散率、籽粒的線性和立方吸濕膨脹系數。
猜你喜歡
品牌研究(2022年27期)2022-09-28 00:30:14
品牌研究(2022年26期)2022-09-19 05:54:48
艦船科學技術(2022年13期)2022-08-11 09:29:16
快樂語文(2021年36期)2022-01-18 05:49:06
成都信息工程大學學報(2021年4期)2021-11-22 07:44:46
小天使·一年級語數英綜合(2021年10期)2021-10-20 02:41:35
World Journal of Diabetes(2019年7期)2019-07-23 11:52:08
中國糧食經濟(2018年11期)2018-12-27 08:58:42
四川輕化工大學學報(自然科學版)(2017年3期)2017-06-29 12:00:57
新農業(2016年17期)2016-08-16 12:02:38
