EGCG對Ro52介導的THP-1細胞凋亡及炎癥因子的影響
2018-09-18 00:57:48張懿劉暢
中國現代醫學雜志 2018年26期
張懿,劉暢
(1.錦州醫科大學,遼寧 錦州 121000;2.錦州醫科大學附屬第一醫院內分泌科,遼寧 錦州121000)
系統性紅斑狼瘡(systemic lupus erythematous,SLE)累及多系統、多器官,發病機制復雜[1-2]。茶多酚活性成分表沒食子兒茶素沒食子酸酯(epigallocatechin gallate, EGCG)對狼瘡性腎炎的調控作用顯著[3-4]。SSA/Ro52(Ro52)是SLE的關鍵調控靶點[5-9],其調控機制可能與單核-巨噬細胞凋亡和炎癥反應密切相關[10-13]。EGCG能顯著抑制正常體細胞中Ro52的表達[14]。然而,單核-巨噬細胞中EGCG與Ro52的功能和調控關系并不清楚。本研究旨在探索不同濃度EGCG對Ro52介導的單核-巨噬細胞THP-1凋亡及炎癥反應的調控效應,為SLE發病機制的研究提供新思路。
1 材料與方法
1.1 主要材料與試劑
1.1.1 主要材料 人單核-巨噬細胞THP-1(中國科學院上海微生物所菌種保藏中心),Ro52過表達質粒(pCMV-Ro52)(北京Biovector公司)。
1.1.2 主要試劑及儀器 腫瘤壞死因子-α(tumor necrosis factor-α, TNF-α)、白介素 -1β(Interleukin-1β, IL-1β)、IL-13檢測試劑盒購自美國Trevigen公司,M-MLV逆轉錄酶(美國Clontech公司),SYBR Green Master Mix(美國 Life Technologies公司),BCA蛋白檢測試劑盒、anti-Ro52、anti-甘油醛-3-磷酸脫氫酶(glyceraldehyde-3-phosphate dehydrogenase, GAPDH)抗體、辣根過氧化酶二抗購自美國Santa Cruz Biotechnology公司,RPMI 1640培養基(美國Abcam公司),膜聯蛋白-熒光素異硫氰酸酯(Annexin V-fluorescein isothiocyanate, AV-FITC)(美國 BD Pharmingen 公司),碘化丙啶(propidium iodide, PI)(美國Sigma公司),Trizol試劑盒(美國Invitrogen公司),逆轉錄試劑盒(德國Fermentas公司),聚偏二氟乙烯膜(polyvinylidene fluoride, PVDF)(美國Millipore公司),酶聯免疫吸附法(enzyme-linked immunosorbent assay, ELISA)試劑盒(美國Santa Cruz公司)Image-Pro Plus 6軟件(美國 Media Cybernetics公司),BD FACSCalibur?系統(美國BD公司),Beckman CXP軟件(美國Brea公司)。
1.2 人單核-巨噬細胞THP-1的培養與分組
1.2.1 人單核-巨噬細胞THP-1的培養 在37℃、5%二氧化碳CO2條件下,用含10% FBS和1%青-鏈霉素的RPMI 1640培養基培養人單核-巨噬細胞THP-1。細胞2 d或3 d換液1次,細胞融合率達80%時,進行傳代,將細胞濃度調整為1.0×106個/ml用于后續實驗。
1.2.2 細胞分組 正常培養的THP-1細胞為對照組,20μg pCMV-Ro52為 pCMV-Ro52組,20μg pCMVNC為pCMV-NC組[15],以及Ro52過表達48 h后繼續使用0、5、10、20和50 mg/L EGCG處理24 h的0、5、10、20 和 50 mg/L EGCG 組[16]。
1.3 Ro52過表達質粒的構建與細胞轉染
采用實時熒光定量聚合酶鏈反應(quantitative real-time polymerase chain reaction, qRT-PCR)對人源Ro52的cDNA(NM_003141.3)進行擴增,并且將產物插入pCMV質粒,進行轉染效率優化篩選。將濃度為1.0×106個/ml的THP-1細胞轉入24孔板中培養,參考謝軍等[17]的研究,使用pCMV-NC質粒或者pCMV-Ro52分別對THP-1細胞進行轉染。48 h后采用qRT-PCR進行擴增。Ro52正向引物:5’-AGAGAGACTTCACCTGTTCTGT-3’,反向引物:5’-TCAGTTCCCCTAATGCCACCT-3’。采用 Western blot檢測THP-1細胞中Ro52的表達。
1.4 qRT-PCR
參照qRT-PCR試劑盒說明書抽提細胞總RNA,采用Trizol試劑盒和逆轉錄試劑盒將5.0μg RNA作為模板合成cDNA,用SYBR Green Master Mix進行基因表達的實時分析。反應條件:95℃變性5 min,95℃延伸5 s,60℃退火1 min,共40個循環。采用2-ΔΔCt法計算mRNA的相對表達量,每組數據以倍數變化進行展示。GAPDH用于qRT-PCR結果的標準化處理。
1.5 Western blot檢測
收集各處理組細胞進行裂解,并抽提總蛋白質。采用12%十二烷基硫酸鈉聚丙烯酰胺凝膠電泳分離不同大小的蛋白質,用聚偏二氟乙烯膜(polyvinylidene fluoride, PVDF)進行濕法轉膜。用3%脫脂牛奶對轉膜完成的PVDF膜室溫封閉1.5 h,PBS洗滌3次,5 min/次,采用一抗包括goat源性的Ro52(1∶900)和GAPDH(1∶2 000)4℃孵育過夜。洗去一抗,在室溫條件下用辣根過氧化酶二抗(1∶18 000)孵育1.5~2.0 h,洗滌抗體液。采用增強化學發光法顯影。Western blot檢測結果用Image-Pro Plus 6軟件進行圖像分析。
1.6 細胞凋亡實驗
收集各組細胞,采用BD FACSCalibur?系統、AV-FITC及PI試劑盒,依據試劑盒說明書進行細胞凋亡檢測。主要步驟為:不同處理條件下的細胞以1.0×106個/ml的密度懸浮于100 ml結合緩沖液,將細胞用AV-FITC和PI孵育15 min,在FC-500流式細胞儀上使用Beckman CXP軟件進行分析。
1.7 ELISA
收集各組細胞,將細胞懸液6 070 r/min離心15 min。將上清液收集并儲存于80℃,用于炎癥因子分析。根據試劑盒說明書進行操作,炎癥因子TNF-α、IL-1β、IL-13采用ELISA試劑盒進行檢測。
1.8 統計學方法
數據分析采用SPSS 17.0統計軟件,計量資料以均數±標準差(±s)表示,比較用方差分析,兩兩比較用LSD-t檢驗,P<0.05為差異有統計學意義。
2 結果
2.1 Ro52過表達效果
2.1.1 Ro52 mRNA 檢測pCMV-Ro52轉染48 h后對Ro52 mRNA和蛋白表達的影響。對照組、pCMV-NC組、pCMV-Ro52組Ro52 mRNA相 對 表達 量 分 別 為(0.988±0.021)、(1.003±0.029) 和(2.558±0.418),經方差分析,差異有統計學意義(F=8.543,P=0.000)。進一步兩兩比較經LSD-t檢驗,pCMV-Ro52組Ro52 mRNA表達水平較對照組升高(P<0.05);而pCMV-NC組與對照組比較,差異無統計學意義(P>0.05);與pCMV-NC組相比,pCMVRo52組Ro52 mRNA表達水平升高(P<0.05)。
2.1.2 Ro52蛋白 對照組、pCMV-NC組、pCMVRo52組Ro52蛋白相對表達量分別為(0.989±0.021)、(1.000±0.065)和(2.291±0.815),經方差分析,差異有統計學意義(F=62.134,P=0.000)。進一步兩兩比較經LSD-t檢驗,pCMV-Ro52組Ro52蛋白表達水平較對照組升高(P<0.05);而pCMV-NC組與對照組比較,差異無統計學意義(P>0.05);與pCMVNC組相比,pCMV-Ro52組Ro52蛋白表達水平升高(P<0.05)。結果表明,pCMV-Ro52轉染THP-1細胞48 h可上調Ro52的表達。見圖1。
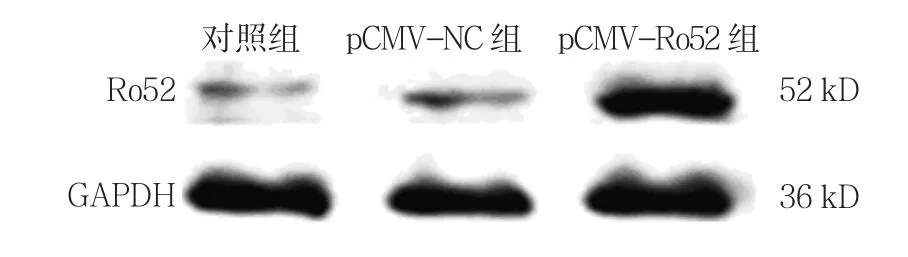
圖1 各組Ro52蛋白的表達
2.2 Ro52過表達對THP-1細胞凋亡的影響
Ro52過表達質粒pCMV-Ro52轉染細胞48 h后檢測其對THP-1細胞凋亡比例的影響。對照組、pCMV-NC組、pCMV-Ro52組THP-1細胞凋亡率分 別 為(2.401±1.030)%、(2.532±0.512)% 和(10.337±0.987)%,經方差分析,差異有統計學意義(F=62.134,P=0.000)。進一步兩兩比較經LSD-t檢驗,pCMV-Ro52組THP-1細胞凋亡率較對照組升高(P<0.05);而pCMV-NC組與對照組比較,差異無統計學意義(P>0.05);與pCMV-NC組相比,pCMVRo52組THP-1細胞凋亡率升高(P<0.05)。結果表明,pCMV-Ro52轉染THP-1細胞48h可上調THP-1細胞凋亡率。見圖2。
2.3 Ro52過表達對THP-1細胞炎癥因子水平的影響
Ro52過表達質粒pCMV-Ro52轉染細胞48 h后對照組、pCMV-NC組、pCMV-Ro52組TNF-α、IL-1β、IL-13水平比較,經方差分析,差異有統計學意義(P<0.05)。進一步兩兩比較經LSD-t檢驗,pCMV-Ro52組 TNF-α、IL-1β、IL-13水 平較對照組升高(P<0.05);pCMV-NC組與對照組TNF-α、IL-1β、IL-13水平比較,差異無統計學意義(P>0.05);與pCMV-NC組相比,pCMV-Ro52組 TNF-α、IL-1β、IL-13水 平 升 高(P<0.05)。結果表明,pCMV-Ro52轉染THP-1細胞48 h可上調THP-1細胞分泌的炎癥因子TNF-α、IL-1β與IL-13。見表1。

圖2 Ro52過表達對THP-1細胞凋亡的影響
表1 各組炎癥因子水平的變化 (ng/ml,±s)

表1 各組炎癥因子水平的變化 (ng/ml,±s)
注:?與對照組、pCMV-NC組比較,P <0.05
組別 TNF-α IL-1β IL-13對照組 1.034±0.021 1.011±0.032 1.002±0.072 pCMV-NC 組 1.030±0.064 1.026±0.064 1.074±0.050 pCMV-Ro52組 2.233±0.491? 2.157±0.336? 2.335±0.316?F值 5.347 9.257 14.352 P值 0.002 0.001 0.001
2.4 EGCG抑制Ro52的過表達
使用0、5、10、20和50 mg/L EGCG處理過表達Ro52的細胞24 h,對照組,以及0、5、10、20和50 mg/L EGCG組Ro52蛋白相對表達量分別為(0.974±0.069)、(2.425±0.562)、(2.032±0.683)、(1.907±0.637)、(1.715±0.367)、(1.308±0.102),經方差分析,差異有統計學意義(F=31.342,P=0.000)。進一步兩兩比較經LSD-t檢驗,與0 mg/L EGCG組相比,20和50 mg/L EGCG組Ro52蛋白水平降低(P<0.05);0 mg/L EGCG 組 與 10 mg/L EGCG 組 比較,差異無統計學意義(P>0.05);0 mg/L EGCG組與5 mg/L EGCG組比較,差異無統計學意義(P>0.05)。見圖3。
2.5 EGCG調控THP-1細胞凋亡
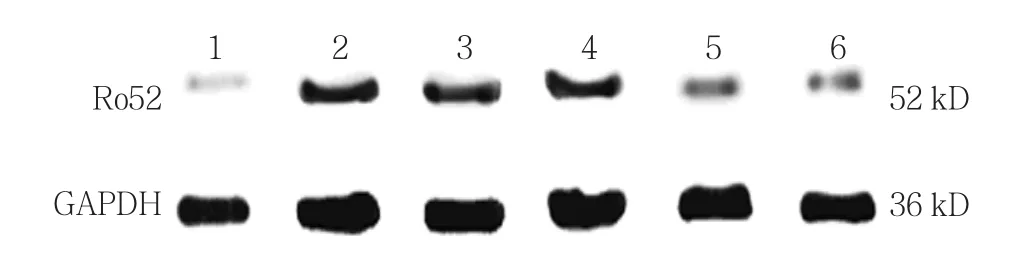
圖3 不同濃度EGCG對Ro52蛋白表達的影響
使用不同濃度(0、5、10、20和 50 mg/L)EGCG處理過表達Ro52的THP-1細胞24 h后,對照組,以及0、5、10、20和50 mg/L EGCG組THP-1細胞凋亡率分別為(2.362±0.213)%、(10.380±0.682)%、(10.061±0.123)%、(9.650±0.781)%、(6.352±0.902)%、(5.234±0.321)%,經方差分析,差異有統計學意義(F=82.354,P=0.000)。進一步兩兩比較經LSD-t檢驗,與0 mg/L EGCG組相比,20和50 mg/L EGCG組細胞凋亡率降低(P<0.05);0 mg/L EGCG組與10 mg/L EGCG組比較,差異無統計學意義(P>0.05);0 mg/L EGCG組與5 mg/L EGCG組比較,差異無統計學意義(P>0.05)。見圖4。
2.6 EGCG阻斷Ro52對細胞炎癥因子的影響
2.6.1 TNF-α 使用不同濃度(0、5、10、20和 50 mg/L)EGCG處理過表達Ro52的THP-1細胞24 h后,對照組,以及0、5、10、20和50 mg/L EGCG組TNF-α、IL-1β、IL-13水平比較,經方差分析,差異有統計學意義(P<0.05)。進一步兩兩比較經LSD-t檢驗,與0 mg/L EGCG組相比,20和50 mg/LEGCG組TNF-α水平降低(P<0.05);0 mg/L EGCG組與10 mg/L EGCG組比較,差異無統計學意義(P>0.05);0 mg/L EGCG組與5 mg/L EGCG組比較,差異無統計學意義(P>0.05)。見表2。
2.6.2 IL-1β對照組,以及0、5、10、20和50 mg/L EGCG組IL-1β水平比較,經方差分析,差異有統計學意義(P<0.05)。進一步兩兩比較經LSD-t檢驗,與0 mg/L EGCG組相比,20和50 mg/L EGCG組 IL-1β 水平降低(P<0.05);0 mg/L EGCG 組與10 mg/L EGCG組比較,差異無統計學意義(P>0.05);0 mg/L EGCG組與5 mg/L EGCG組比較,差異無統計學意義(P>0.05)。見表2。
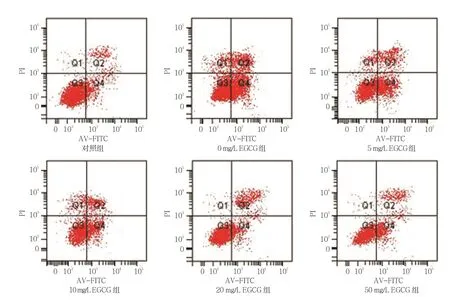
圖4 EGCG對THP-1細胞凋亡的影響
表2 各組炎癥因子水平的變化 (ng/ml,±s)
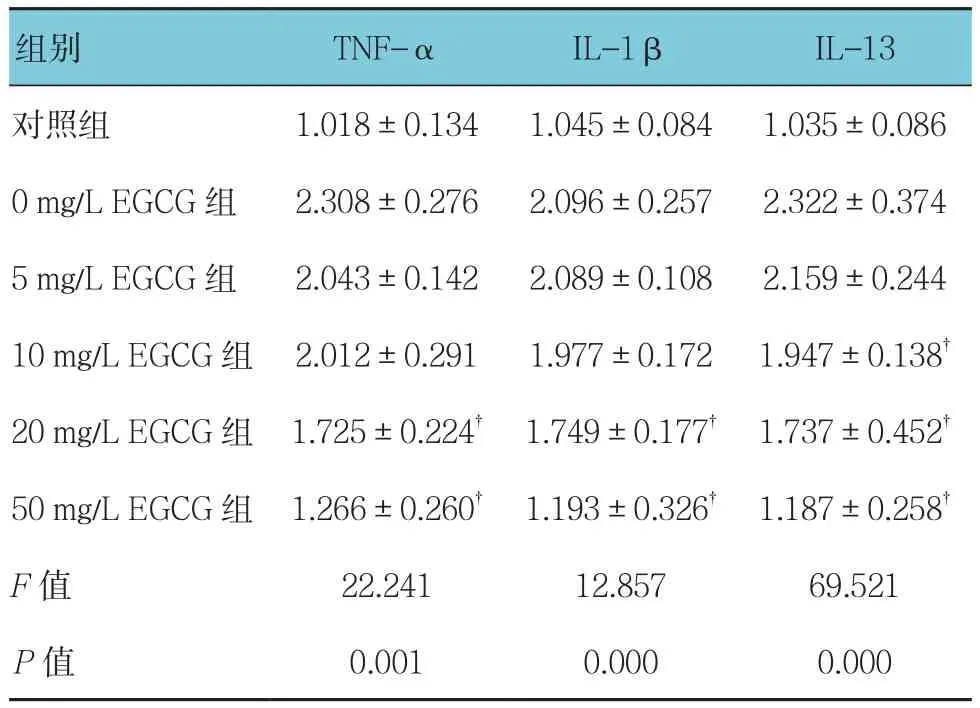
表2 各組炎癥因子水平的變化 (ng/ml,±s)
注:?與0 mg/L EGCG組比較,P <0.05
組別 TNF-α IL-1β IL-13對照組 1.018±0.134 1.045±0.084 1.035±0.086 0 mg/L EGCG 組 2.308±0.276 2.096±0.257 2.322±0.374 5 mg/L EGCG 組 2.043±0.142 2.089±0.108 2.159±0.244 10 mg/L EGCG 組 2.012±0.291 1.977±0.172 1.947±0.138?20 mg/L EGCG組 1.725±0.224? 1.749±0.177? 1.737±0.452?50 mg/L EGCG組 1.266±0.260? 1.193±0.326? 1.187±0.258?F值 22.241 12.857 69.521 P值 0.001 0.000 0.000
2.6.3 IL-13 對照組,以及0、5、10、20和50 mg/L EGCG組IL-13水平比較,經方差分析,差異有統計學意義(P<0.05)。進一步兩兩比較經LSD-t檢驗,與0 mg/L EGCG組相比,10、20和50 mg/L EGCG組IL-13水平降低(P<0.05);0 mg/L EGCG組與 5 mg/L EGCG組比較,差異無統計學意義(P>0.05)。見表2。
3 討論
3.1 Ro52參與單核-巨噬細胞凋亡和破壞正常的炎癥因子水平
本研究發現,過表達Ro52既可以促進THP-1細胞凋亡,又可以促進THP-1細胞TNF-α、IL-1β和IL-13的分泌。這說明,Ro52在單核-巨噬細胞的功能紊亂和炎癥因子分泌中發揮調控作用。單核-巨噬細胞在SLE的起病過程中發揮重要作用,包括調節炎癥反應,誘導免疫應答、清除凋亡細胞等[18],同時還發現巨噬細胞在免疫應答啟動過程中是多種細胞因子的重要來源[19]。TABAS等[20]研究表明,SLE相關的并發癥,如狼瘡腎炎的早期和進展期均可出現大量巨噬細胞的異常凋亡,巨噬細胞本身負責清除凋亡細胞并調節自身生理性正常凋亡,而SLE患者體內的巨噬細胞吞噬功能異常,以及自身免疫疾病中巨噬細胞的自身凋亡調控功能同樣顯示異常,這暗示巨噬細胞自身異常凋亡可能與SLE密切相關。
董曉薇[9]等研究表明,SLE患者血清中靶抗原Ro52蛋白表達水平顯著升高,這可能與SLE患者心血管疾病加劇并伴有單核-巨噬細胞異常凋亡增加,以及細胞炎癥因子分泌增加有關。Ro52蛋白是SLE等多種自身免疫性疾病過程中的主要靶點[6-8],可能在調節凋亡和炎癥反應過程中扮演重要角色[21]。對SLE心臟受累患者的研究中發現,抗Ro52抗體與胎兒凋亡的心肌細胞表面Ro52抗原結合后激活巨噬細胞[22]。同時發現,Ro52在細胞內免疫過程中發揮重要的凋亡調節作用[10]。BLAKE等[10]研究發現,石棉暴露導致的自身免疫疾病可促進巨噬細胞凋亡,并且凋亡的巨噬細胞表面Ro52表達上調。本研究明確Ro52可以調控單核-巨噬細胞的凋亡及炎癥因子TNF-α、IL-1β、IL-13的分泌。結果表明,Ro52作為重要的前凋亡分子在自身免疫疾病的巨噬細胞凋亡和炎癥反應過程中起促進作用,而且很有可能是SLE免疫調控的一個重要途徑。
3.2 EGCG減緩Ro52對單核-巨噬細胞凋亡和炎癥因子水平的調控
EGCG是一種綠茶多酚類分子[3,23],被稱為自身免疫性疾病的潛在治療武器,已經在治療干燥綜合癥等疾病、增強免疫系統功能與抑制腫瘤發生等領域中有深入研究[24]。本研究發現,EGCG能夠在20和50 mg/L 2個濃度顯著減緩Ro52過表達質粒轉染THP-1細胞導致的Ro52超高表達,并且可以阻止THP-1細胞被Ro52過表達后導致的細胞凋亡增加。與pCMV-Ro52組相比,細胞炎癥因子TNF-α、IL-1β、IL-13水平被EGCG抑制。結果表明,EGCG是一種有效的抗凋亡和抗炎分子,可以減緩阻斷Ro52的促細胞凋亡效應和促炎癥因子效應。
以往研究同樣發現,EGCG能夠有效抑制巨噬細胞凋亡,減輕免疫細胞的炎癥反應[11,13]。HASHIMOTO等[25]研究發現,抑制EGCG可明顯誘導小鼠巨噬細胞樣細胞的凋亡。研究發現EGCG對Ro52具有明顯的抑制效應[14]。本研究結果發現,EGCG顯著抑制Ro52誘發的THP-1細胞凋亡,與上述結果一致。HSU等[14]在研究正常人原發性表皮角質形成細胞與永生化人涎腺腺泡細胞功能時發現,EGCG能夠顯著抑制Ro52等多種自身抗原的表達。結果表明,Ro52可被EGCG負向調控,并且EGCG可通過抑制Ro52,介導人單核-巨噬細胞的凋亡與炎癥反應。
綜上所述,Ro52過表達可促進人單核-巨噬細胞THP-1的凋亡與炎癥因子TNF-α、IL-1β、IL-13的分泌。EGCG可減弱Ro52介導的THP-1細胞凋亡,減少炎癥因子TNF-α、IL-1β、IL-13的分泌。本實驗為進一步研究SLE等自身免疫疾病中巨噬細胞的功能、SLE的預防與治療,以及SLE的發病機制提供實驗依據。
猜你喜歡
新少年(2022年9期)2022-09-17 07:10:54
音樂探索(2022年2期)2022-05-30 21:01:37
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
小天使·一年級語數英綜合(2020年6期)2020-12-16 02:56:41
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
人大建設(2019年12期)2019-05-21 02:55:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
北極光(2014年8期)2015-03-30 02:50:51
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國火炬(2010年8期)2010-07-25 11:34:30
