淮河源區群體遺傳多樣性研究
2018-09-20 04:10:42方冬冬顧錢洪周傳江孟曉林李學軍聶國興
水產科學 2018年5期
方冬冬,顧錢洪,周傳江,孟曉林,李學軍,聶國興
( 河南師范大學 水產學院,河南省水產動物養殖工程技術研究中心,河南 新鄉 453007 )
1 材料與方法
1.1 樣本采集
2015年7月—2016年4月,采集河南省信陽市淮河源區不同河流10個地理群體的樣本,采樣站位分別位于譚家河、南灣、高店鄉、九龍河、小潢河、潑陂河、鄔橋、李渡口、九曲河、灌河。具體采樣信息見圖1、表1。
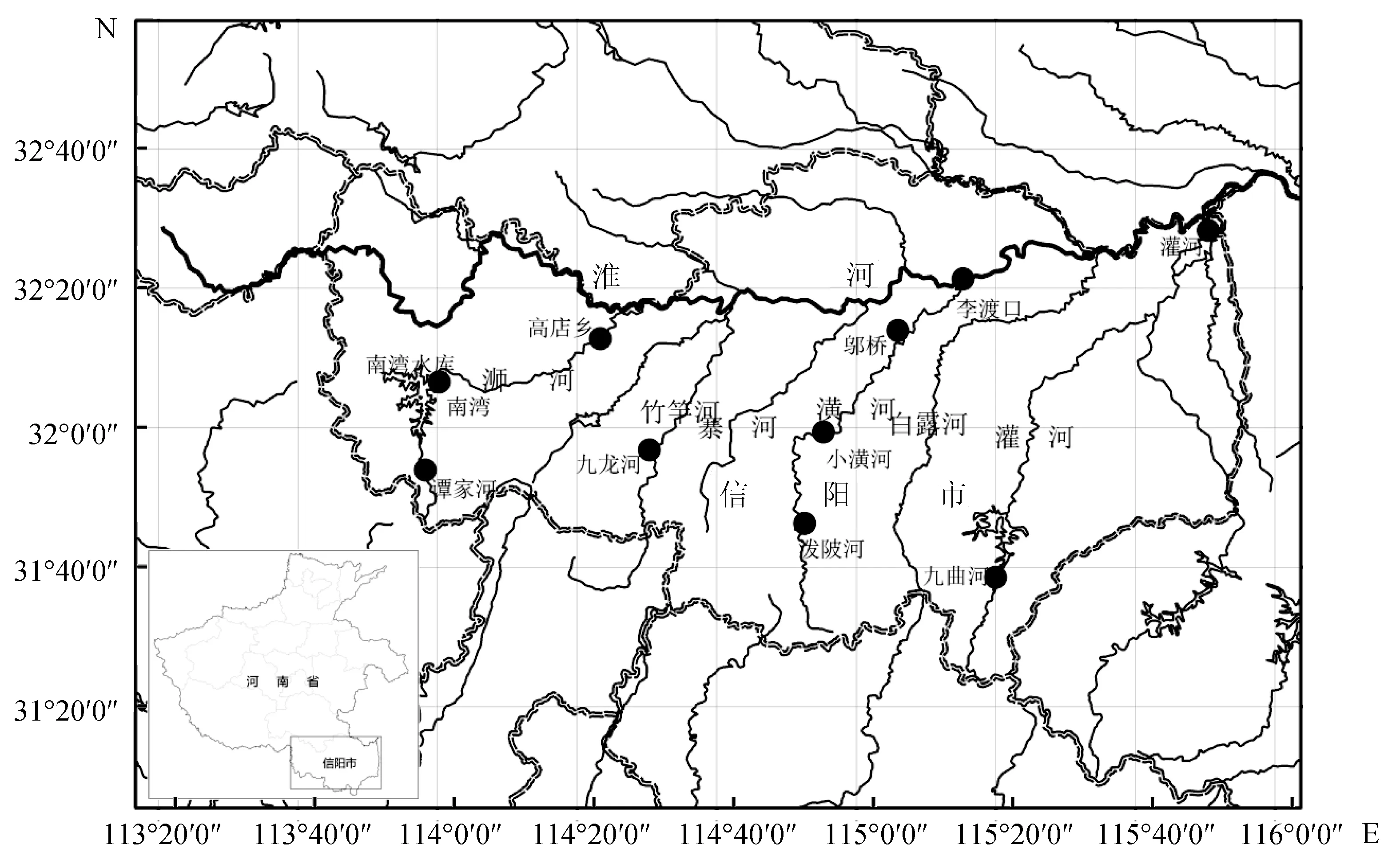
圖1 樣品采集圖(圖中黑色圓圈表示采樣站位)
1.2 DNA的提取、擴增與測序
DNA提取的方法是取其左側鰭條或是背側肌肉,置于無水乙醇中。采用標準的酚—氯仿法提取基因組DNA。用1.2%瓊脂糖凝膠電泳和紫外分光光度計檢測DNA質量和含量,并將合格的基因組DNA稀釋到100 μg/L,保存好后用于后續試驗。用核基因RAG2和線粒體基因Cytb作為分子標記,擴增目的片段。RAG2基因和Cytb基因分別采用通用引物:RAG2-108f(5′-CCVARACGCTCATGTCCAAC-3′),RAG2-1324r(5′-TGGARCAGWAGATCATKGC-3′)[11];Cytb_F2(5′-AACCACCGTTGTATTCAACTACAA-3′),Cytb_R2(5′-ACCTCCGATCTTCGGATTACAAGACCG-3′)[12]。PCR反應總體積均為50 μL,其中10×Taq Buffer 5 μL,dNTP 4 μL,上下游引物(10 μmol/L)各1 μL,TaqDNA聚合酶(5 U/μL)0.2 μL,DNA 模板 20 ng,ddH2O 補足體積。樣品在BIO-RAD PCR儀上進行擴增,RAG2的反應條件:94 ℃預變性3 min,94 ℃變性30 s,51 ℃退火30 s,72 ℃延伸1 min,34個循環,72 ℃最后延伸7 min。Cytb的反應條件:94 ℃預變性4 min,94 ℃變性30 s,50 ℃退火30 s,72 ℃延伸1 min,34個循環,72 ℃最后延伸8 min。PCR產物使用1.2%的瓊脂糖凝膠電泳檢測后經過純化,送武漢天一輝遠生物科技有限公司進行雙向測序,測序引物與PCR引物相同。
1.3 數據統計與分析
測序得到的每個個體序列進行組裝、校正,得到的一致序列后在GenBank中進行BLAST搜尋,確認為目的片段序列用于后續數據分析。使用ClustalX 2軟件進行多重比對并輔以人工校對。使用DnaSP 5.10軟件對單倍型數、單倍型多樣性、核苷酸多樣性、多態位點、平均核苷酸差異數等遺傳多樣性參數進行計算[13];使用MEGA 6.0軟件計算群體間的遺傳距離[14],使用NETWORK 5.001軟件構建單倍型之間的進化關系圖。使用ARLEQUIN 3.0軟件中的AMOVA方法分析群體間的遺傳結構和遺傳變異[15]。先將的10個群體劃分為1個組群,以驗證種群間是否具有顯著的遺傳分化,然后按不同河流將的不同群體劃分為4個組群,驗證是否存在顯著的地理結構;采用遺傳分化參數評價種群間的遺傳分化程度;采用Fu′sFS檢驗和Tajima′sD進行中性檢驗,結合核苷酸錯配分布圖推斷群體歷史是否發生近期擴張。利用MIGRATE-n v 3.0[16]計算群體間遷移率。
2 結 果
本試驗得到長度為1145 bp的RAG2基因片段,137條序列共檢測到41個單倍型,其中13個為共享單倍型體,其余為群體特有單倍型,變異位點13個,約占全序列的1.13%;得到Cytb的基因片段長度為1110 bp,152條序列共檢測到27個單倍型,其中有8個單倍型為群體所共享,其余為群體特有單倍型,變異位點14個,約占全序列的2.43%。
遺傳多樣性分析結果見表1,RAG2基因所有樣本平均單倍型多樣性為0.940±0.011,核苷酸多樣性為0.00246±0.00057;Cytb基因所有樣本平均單倍型多樣性為0.915±0.011,核苷酸多樣性為0.00461±0.00060。在所有獨立種群中,RAG2分析結果為灌河群體的單倍型多樣性與核苷酸多樣性均為最低,分別為0.645±0.106、0.00090±0.00013;Cytb的分析結果顯示,小潢河群體的單倍型多樣性與核苷酸多樣性為所有群體中最低。
表1 淮河源區10個群體代碼、樣本量及遺傳多樣性

表1 淮河源區10個群體代碼、樣本量及遺傳多樣性
群體站位經緯度基因樣本數量單倍型數目單倍型多樣性核苷酸多樣性多態位點譚家河N 31°53'51.50″E 113°58'15.60″RAG219140.965±0.0280.00390±0.0003910Cytb2060.800±0.0540.00356±0.000459南灣N 32°06'31.70″E 114°00'16.30″RAG217110.882±0.0720.00168±0.000174Cytb2050.826±0.0360.00336±0.000458高店鄉N 32°12'42.90″E 114°23'19.70″RAG2881.000±0.0630.00299±0.000296Cytb940.750±0.1120.00345±0.000477九龍河N 31°56'45.06″E 114°30'21.97″RAG215100.943±0.0400.00221±0.000175Cytb1760.721±0.0870.00344±0.000369小潢河N 31°59'19.90″E 114°55'18.30″RAG21240.803±0.0630.00094±0.000092Cytb1120.509±0.1010.00229±0.000455潑陂河N 31°46'14.29″E 114°52'34.88″RAG2840.786±0.1130.00100±0.000132Cytb1250.833±0.0690.00351±0.000448鄔橋N 32°13'52.60″E 115°05'58.00″RAG21770.868±0.0500.00123±0.000123Cytb1850.641±0.0970.00323±0.000488李渡口N 32°21'17.70″E 115°15'19.40″RAG21440.901±0.0460.00180±0.000134Cytb1750.750±0.0690.00299±0.000427九曲河N 31°38'29.26″E 115°19'58.67″RAG21490.934±0.0450.00177±0.000134Cytb1560.800±0.0710.00400±0.000349灌河N 31°53'51.50″E 113°58'15.60″RAG21340.645±0.1060.00090±0.000132Cytb1340.654±0.1060.00379±0.000448總計RAG2137410.940±0.0110.00246±0.0005713Cytb163270.906±0.0120.00452±0.0005914
基于核基因RAG2序列與線粒體基因Cytb序列對淮河源區10個群體進行了AMOVA分析(表2),RAG2的分析結果顯示在總遺傳變異中,組內種群間的遺傳變異占24.64%,Cytb的結果略高于RAG2,組內種群間的遺傳變異占總遺傳變異的26.14%。按不同水系將10個群體劃分為4個組群進行多層次的AMOVA分析,RAG2的分析結果顯示,淮河水系各群體組間的遺傳變異為0.09%,Cytb的分析結果顯示,各群體組間的遺傳變異為2.97%,均不具有顯著性差異,遺傳變異主要來自于群體內。
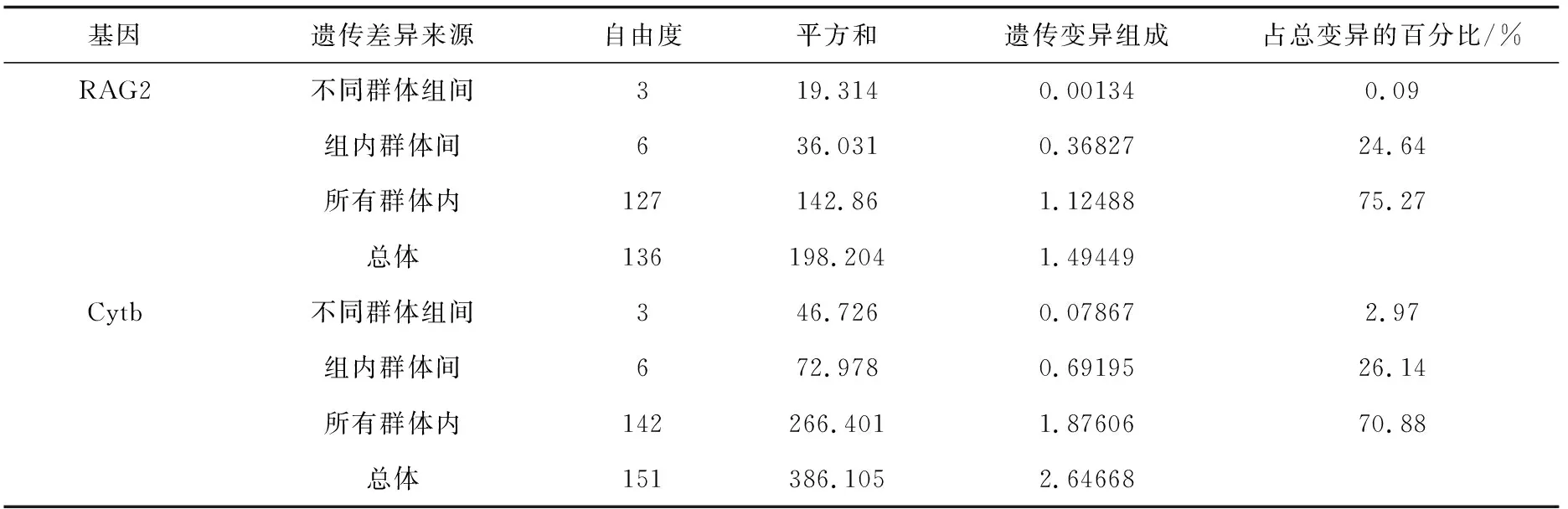
表2 基于RAG2、Cytb序列AMOVA分析
基于核基因RAG2序列與線粒體基因Cytb序列,利用MIGRATE軟件分別計算了群體間的遷移率(表3,表4)。RAG2基因的分析結果表明,群體間的遷移率為0.21(潑陂河→九曲河)~24.02(灌河→南灣),南灣群體與其他群體之間具有最高的單向遷入率(198.36)和最低的單向遷出率(44.2),而九曲河群體與其他群體之間具有最低的單向遷入率(2.22)和最高的單向遷出率(61.82)。Cytb基因的分析結果表明,群體間的遷移率為0.09(譚家河→鄔橋)~6.20(李渡口→高店鄉),各群體單向遷入率和單向遷出率具有不對稱性,其中高店鄉群體具有最高的單向遷入率,為40.00,遷入率最低的李渡口群體,為1.95,單向遷出率最高的是灌河群體,為10.47,遷出率最低的是九龍河群體,為5.05。
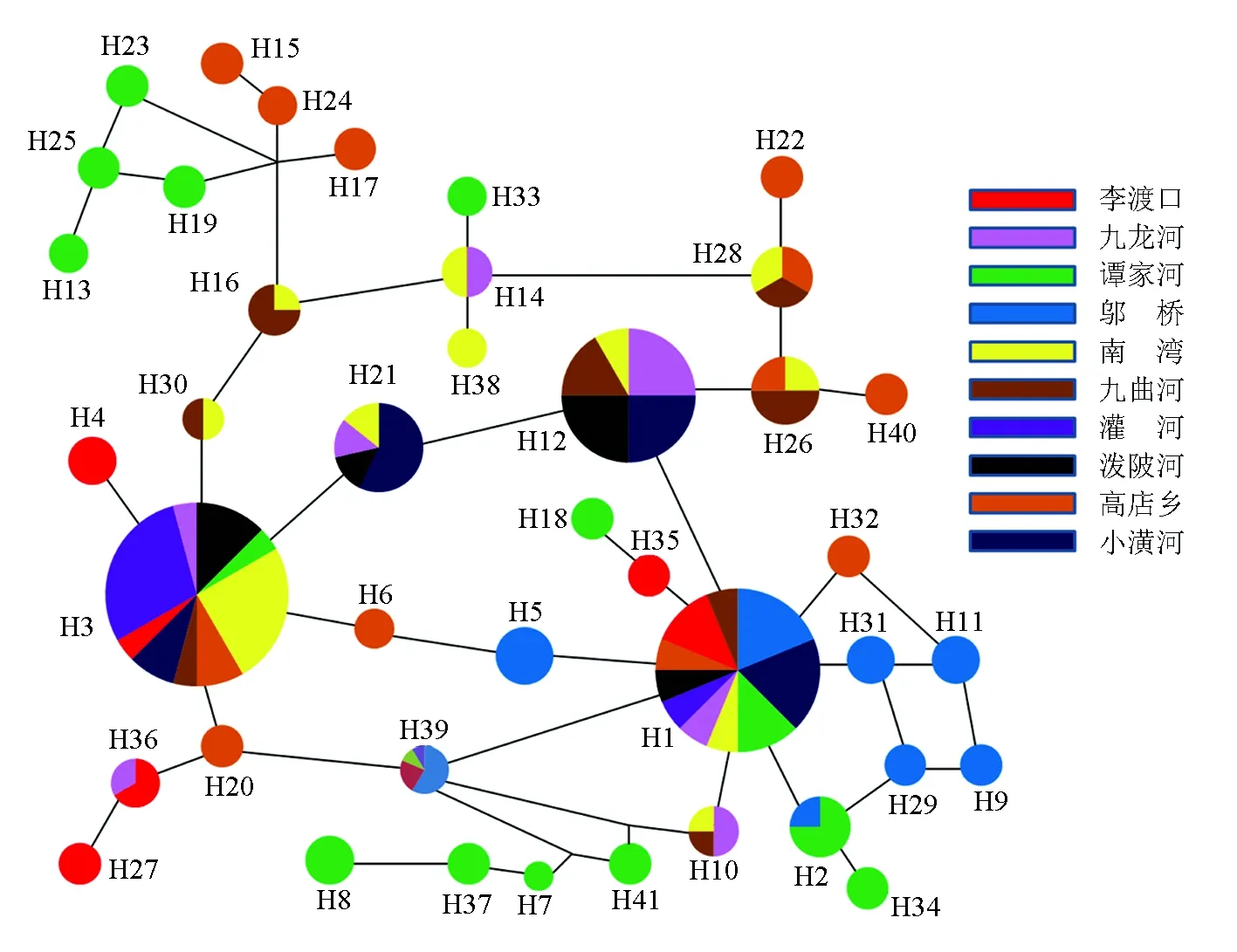
圖2 基于RAG2序列構建的單倍型網絡分析
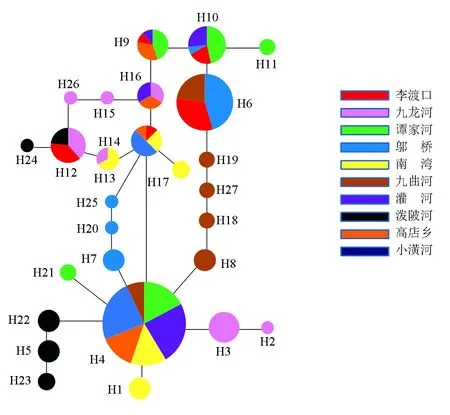
圖3 基于Cytb序列構建的單倍型網絡分析
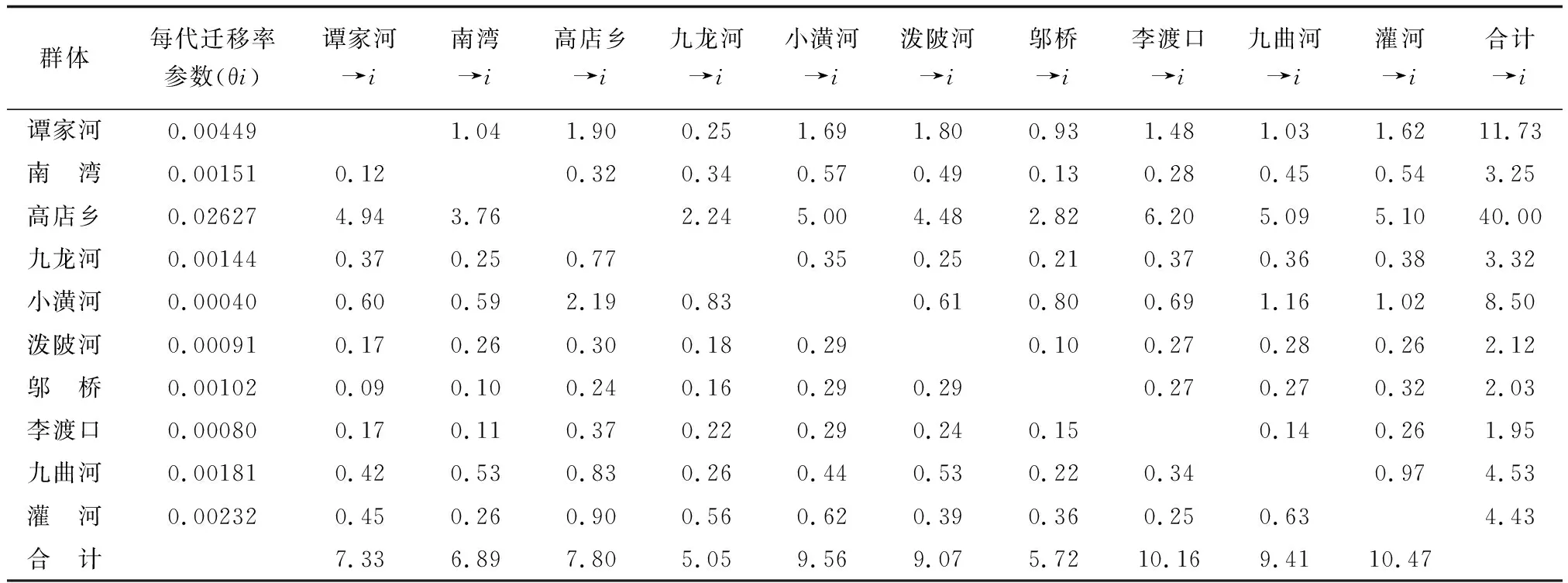
群體 每代遷移率參數(θi)譚家河→i南灣→i高店鄉→i九龍河→i小潢河→i潑陂河 →i鄔橋→i李渡口→i九曲河→i灌河→i合計→i譚家河0.004491.041.900.251.691.800.931.481.031.6211.73南 灣0.001510.120.320.340.570.490.130.280.450.543.25高店鄉0.026274.943.762.245.004.482.826.205.095.1040.00九龍河0.001440.370.250.770.350.250.210.370.360.383.32小潢河0.000400.600.592.190.830.610.800.691.161.028.50潑陂河0.000910.170.260.300.180.290.100.270.280.262.12鄔 橋0.001020.090.100.240.160.290.290.270.270.322.03李渡口0.000800.170.110.370.220.290.240.150.140.261.95九曲河0.001810.420.530.830.260.440.530.220.340.974.53灌 河0.002320.450.260.900.560.620.390.360.250.634.43合 計7.336.897.805.059.569.075.7210.169.4110.47
注:譚家河→i表示譚家河群體與其他群體間的單向遷入率,“i”表示種群,下同.
表4 基于RAG2基因計算10 個群體中的一個群體到其他群體的遷移率

表4 基于RAG2基因計算10 個群體中的一個群體到其他群體的遷移率
群體每代遷移率參數(θi)譚家河→i南灣→i高店鄉→i九龍河→i小潢河→i潑陂河→i鄔橋→i李渡口→i九曲河→i灌河→i合計→i譚家河0.005843.042.632.602.622.842.812.632.732.7824.68南 灣0.0484122.3822.9220.8921.7819.4422.2422.8121.8824.02198.36高店鄉0.01376.06.86.46.65.66.77.36.37.759.4九龍河0.000590.270.310.380.390.270.290.340.290.382.92小潢河0.003511.671.832.101.921.551.752.051.782.1016.75潑陂河0.000580.320.310.280.300.290.300.290.290.272.65鄔 橋0.001260.570.620.820.610.600.550.620.600.635.64李渡口0.016947.489.288.458.088.036.817.717.5110.3673.71九曲河0.000450.270.240.230.240.250.210.240.250.292.22灌 河0.0444819.8521.8121.2119.9620.1718.3819.6422.9820.39184.40合 計58.8444.259.061.0060.7455.6561.7059.2361.8248.55
基于RAG2基因序列與Cytb基因序列的Tajima′sD和Fu′sFs檢驗結果見表5,RAG2基因的Tajima′sD檢驗結果均為正值。Fu′sFS統計結果檢驗,將淮河流域的作為一個整體分析時,Fu′sFS為顯著負值(P<0.001)。核苷酸錯配分布圖為單峰,與Fu′sFs檢驗結果一致。表明群體近期可能經歷過快速擴張。用ARLEQUIN軟件計算得知擴張參數τ為2.17680,其擴張時間約為0.059百萬年。Cytb基因的中性檢驗(表7)和核苷酸錯配分布圖顯示群體未經歷過近期歷史種群擴張。
表5 淮河源區10個群體的Tajima′D和Fu′Fs中性檢驗結果
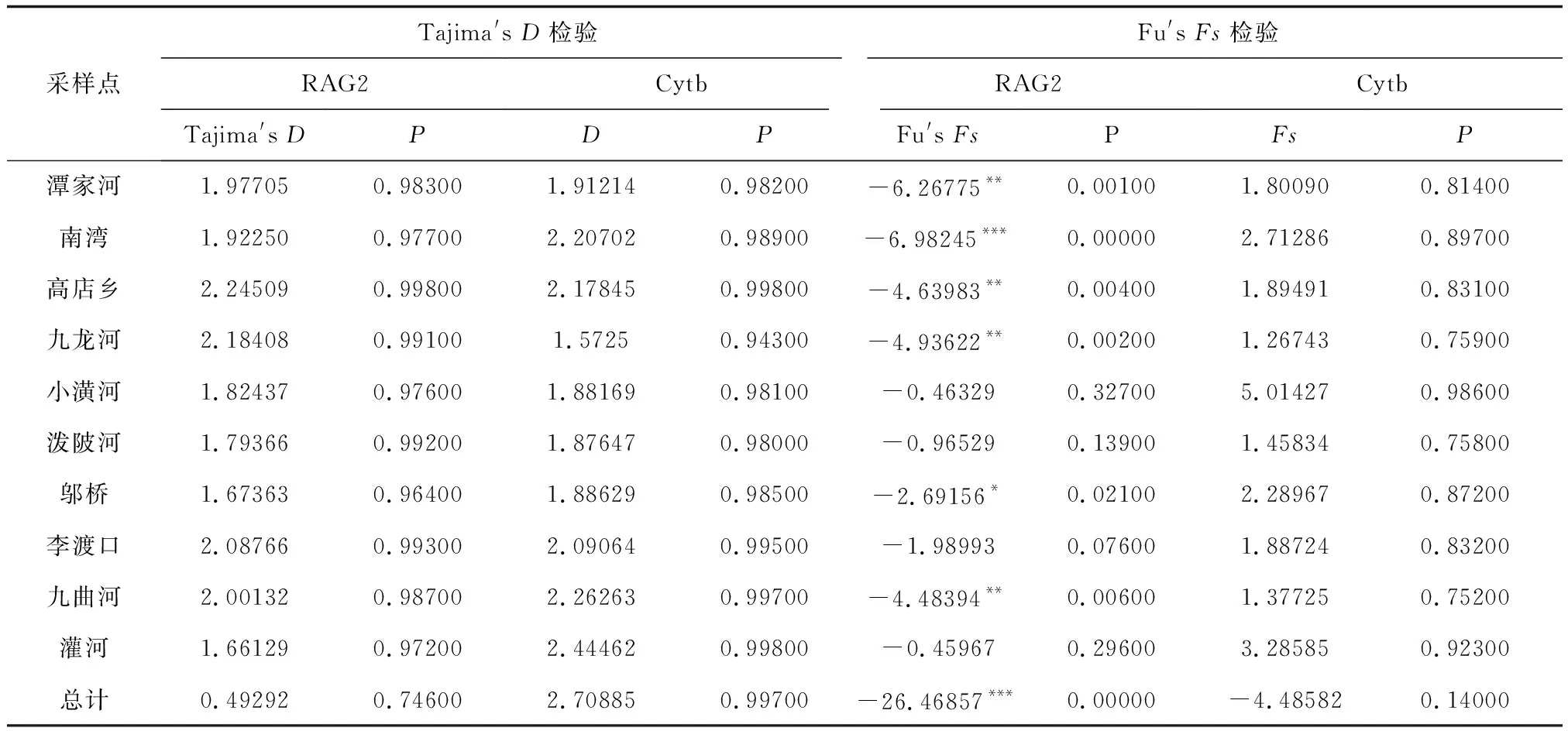
表5 淮河源區10個群體的Tajima′D和Fu′Fs中性檢驗結果
采樣點Tajima's D檢驗Fu's Fs檢驗RAG2CytbRAG2CytbTajima's DPDPFu's FsPFsP潭家河1.977050.983001.912140.98200-6.26775**0.001001.800900.81400南灣1.922500.977002.207020.98900-6.98245***0.000002.712860.89700高店鄉2.245090.998002.178450.99800-4.63983**0.004001.894910.83100九龍河2.184080.991001.57250.94300-4.93622**0.002001.267430.75900小潢河1.824370.976001.881690.98100-0.463290.327005.014270.98600潑陂河1.793660.992001.876470.98000-0.965290.139001.458340.75800鄔橋1.673630.964001.886290.98500-2.69156*0.021002.289670.87200李渡口2.087660.993002.090640.99500-1.989930.076001.887240.83200九曲河2.001320.987002.262630.99700-4.48394**0.006001.377250.75200灌河1.661290.972002.444620.99800-0.459670.296003.285850.92300總計0.492920.746002.708850.99700-26.46857***0.00000-4.485820.14000
注: *P<0.05, **P<0.01, ***P<0.001.
3 討 論
3.1 淮河源區種群遺傳多樣性分析
種群遺傳多樣性的大小主要是由種內遺傳變異的高低與遺傳變異的分布格局共同作用的結果,物種的遺傳多樣性越高其抵抗不良環境的能力就越強[17]。物種遺傳多樣的降低,主要與物種數量下降有關,如過度捕撈、環境污染等[18]。單倍型多樣性與核苷酸多樣性是衡量物種多樣性的重要指標[19]。
本研究結合核基因RAG2與線粒體基因Cytb對淮河源區10個地理群體的進行了遺傳多樣性分析,結果發現10個群體呈現出較高的單倍型多樣性大于0.50和較低的核苷酸多樣性小于0.01的模式。與其他鯉科魚類相比,明顯低于東方墨頭魚(Garraorientalis)、紋唇魚(Osteochilussalsburyi)、眼華鳊(Sinibramamacrops)、雅羅魚(Leuciscusleuciscusbaicalensis),但是略高于一些鯉科經濟魚類,如貢山裂腹魚(Schizothoraxgongshanensis)、青魚(Mylopharyngodonpiceus)、長鰭吻(Rhinogobioventralis)[12,20-24]。本研究中淮河源區出現這種單倍型多樣性高、核苷酸多樣性低的模式,可能是種群受到瓶頸效應后迅速擴張的結果。與長江流域遺傳多樣性(單倍型多樣性為0.9789、核苷酸多樣性為0.03289)相比[5],淮河源區遺傳多樣性明顯偏低。一方面可能與人為干擾、過度捕撈、棲息環境變化等因素有關。陳德蔭[25]在對野生鯪魚(Cirrhinusmolitorella)遺傳多樣性的研究中發現,由于過度捕撈、環境污染等因素的影響,導致鯪魚野生資源不斷下降,遺傳多樣性降低。另一方面也可能與水環境的穩定性有關,淮河水系與長江水系相比,流量、流域面積均小于長江水系。因此,相比長江水系,淮河流域水環境穩定性較差,魚類資源更容易受到環境變化的影響從而導致遺傳多樣性降低[26]。
本研究中兩種分子標記的研究結果基本一致,灌河、小潢河兩群體均呈現出低的單倍型多樣性小于0.50及低的核苷酸多樣性小于0.005的模式,表明兩群體在歷史上可能經歷過嚴重的瓶頸效應或是正在發生瓶頸效應。過度捕撈導致種群數量減少或是種群間不能進行正常的基因交流均會引發遺傳漂變導致遺傳多樣性降低。Kang等[27]研究認為,水庫大壩建設阻礙了魚類種群的擴散,限制了種群間的基因交流,易產生瓶頸效應和遺傳漂變,從而改變種群內和種群間的遺傳多樣性與種群遺傳結構。灌河支流發達,水流湍急,水能資源豐富,共修建大小水庫17座。水庫大壩的建設阻礙了灌河群體與其他群體之間的基因交流,導致遺傳多樣性降低。Ginson等[18]認為,過度捕撈造成種群數量減少,加劇群體間的遺傳漂變,導致種群遺傳多樣性降低。小潢河是途徑光山縣的一條重要河流,當地的漁民經常捕獲作為野雜魚出售,故小潢河群體較其他群體更易受到過度捕撈和環境變化的影響。
線粒體分子標記和核基因分子標記各自有各自的優勢,但研究者更加傾向于將線粒體基因與核基因聯合起來研究魚類的遺傳進化[28]。所以本研究中核基因RAG2和線粒體基因Cytb所揭示的淮河源區遺傳多樣性水平的不同并不矛盾,這是由于各自本身的特點決定的。因此,本研究中的灌河群體與小潢河群體受到人為因素的影響較大,需要加強對該地區魚類資源的保護。
3.2 淮河源區種群遺傳分化分析
遺傳分化系數是衡量群體間遺傳分化的一個重要指標[29],當遺傳分化系數大于0.25時,表示群體間的遺傳分化很大[30]。本研究中對淮河源區10個群體間的遺傳分化分析結果顯示,核基因RAG2群體間遺傳分化系數大于0.25的占40%,線粒體基因Cytb群體間遺傳分化系數大于0.25的占56%。核基因RAG2群體間的遺傳分化沒有線粒體Cytb基因明顯,原因一方面是核基因RAG2在脊椎動物進化過程中具有高度保守性,受時間和空間的嚴格調控[11];另一方面是淮河水系形成于中更新世,晚更新世淮河源區干流出現,群體間的分化時間不夠長,導致種群間存在特異性的基因未固定下來[31]。魏學文[32]基于核基因分子標記對中國大耳菊頭蝠復合體地理學的研究中也有類似的發現。
遺傳分化與基因流之間呈負相關關系,種群間的遺傳分化系數越大其基因流就越小[33]。本研究中兩種分子標記分析群體間的遺傳分化結果一致,鄔橋、李渡口群體與其他群體間的遺傳分化較大,遺傳分化系數大于0.25,貝葉斯推斷種群間的遷移率值大小與種群間的遺傳分化程度基本一致。鄔橋、李渡口群體均有著較高的遷出率和較低的遷入率,貝葉斯推斷種群間的遷移率值的大小直接反應了種群間基因流的大小。分析認為,鄔橋水庫大壩的建設阻礙了該群體與其他群體之間的基因交流,所以產生了較大的遺傳分化。單紅[34]對三峽水庫南方鲇(Silurussoldatovimeridionalis)的遺傳多樣性的研究中也有類似發現,以葛洲壩為界,上游和下游各自為一支,出現了明顯的遺傳分化。李渡口群體位于支流與干流的交匯處,水流湍急,加上地理隔離較為明顯,很少有機會與其他群體之間產生基因交流。同時人類的過度捕撈、環境污染等因素的影響導致河流的生境片段化加速了該群體與其他群體之間的遺傳分化。RAG2與Cytb的AMOVA分析結果顯示,雖然種群間存在著一定的遺傳變異,但大部分的遺傳變異存在于種群內。結合單倍型之間的進化關系,人為因素的影響對魚類種群的遺傳結構沒有產生足夠的影響。因此,群體間較高的基因流與較低的遺傳分化致使淮河源區群體間未形成明顯的群體遺傳結構和譜系格局。
3.3 淮河源區條群體歷史的動態研究
本研究中兩種分子標記的中性檢驗結果與核苷酸錯配分布圖的結果不一致。核基因RAG2的分析結果顯示,淮河源區群體近期發生過種群擴張現象,但Cytb的分析結果與RAG2結果不一致。根據RAG2基因序列間的分歧速率計算得知,群體的擴張時間大約為0.059百萬年,為晚更新世時期。這與徐丹丹[35]提出的長江中上游及珠江水系的鲇魚在晚更新世時期(0.04~0.05 百萬年)發生過種群擴張時間類似。晚更新世,淮河源區正在經歷第四紀冰期最后一次的構造運動,使該區域內的河流改道頻繁,逐漸塑造了現代淮河源區的地形地貌特征和水系構成[36]。為上層魚類,水系的變遷可能加劇了其種群間的遺傳分化。更新世冰期由于氣候寒冷,河流積水大量減少,魚類資源大量減少[37],淮河源區的可能當時也經歷了瓶頸效應,種群數量急劇減少。進入全新世,隨著氣候逐漸變暖,河流積水增加,甚至某些河流發生泛洪,種群快速繁衍,群體迅速擴張。

