甘藍型油菜脯氨酸降解途徑關鍵基因的進化分析
2018-10-11 06:31:18王翠平陳建偉
江蘇農業科學 2018年17期
王翠平, 陳建偉
(寧夏林業研究院種苗生物工程國家重點實驗室,寧夏銀川 750004
在環境脅迫如干旱、鹽、重金屬、紫外線等條件下,高等植物可以在短時間內通過迅速大量地積累脯氨酸等滲透調節物質對自身起到保護作用,以此來增強植物對滲透脅迫的抵抗能力[1-5]。在脅迫條件下,脯氨酸可以作為兼容性滲透調節物質、蛋白質和亞細胞結構、活性氧清除劑和氧化還原平衡劑[6-7]。近年來,有研究發現脯氨酸參與了開花及胚胎發育過程[8-11]。高等植物中,吡咯啉-5-羧酸合成酶(P5CS)是脯氨酸合成途徑的關鍵酶[8-9],而脯氨酸脫氫酶(proline dehydrogenase,PDH)是脯氨酸降解途徑的關鍵酶[10]。在擬南芥中,PDH定位于線粒體,由2個基因編碼(AtPDH1,At3g30775;AtPDH2,At5g30870),脯氨酸處理會誘導PDH1表達,而脅迫處理抑制PDH1表達[11]。
植物進化過程中多倍化是重要的進化動力,并且許多作物經歷了多倍化過程,多倍體能夠為新表型的出現提供原材料,具有進化上的優勢[12]。多倍化之后由于染色體重組和基因重排,有一些基因表達模式會發生變化,甚至會發生丟失[13-14]。蕓薹屬(Brassica)植物是研究由于多倍化引起的基因缺失、沉默和偏向表達的非常好的模式植物體系,禹氏三角模型能夠很好地解釋蕓薹屬植物的進化關系[15]。甘藍型油菜(Bassicanapus)是最新形成的異源四倍體,來源于2個二倍體白菜(B.rapa)和甘藍(B.oleracea)種間雜交,并且經歷了染色體加倍的過程[16]。而白菜、甘藍和擬南芥起源于共同的祖先,大概在1 450萬~2 040萬年之前這個祖先分化為擬南芥和白菜、甘藍的祖先[17]。比較物理圖譜結果表明,白菜和甘藍的祖先經歷了三倍化和基因重排的過程[18]。且白菜基因組測序數據也表明,白菜中大多數基因具有2~3個同源基因[19]。那么,推測異源四倍體油菜中同源基因拷貝數應該更多。有報道稱,油菜中基因拷貝數目可能由于進化過程中基因丟失的原因而減少[20]。
脅迫條件下脯氨酸積累是一個非常保守且有效的過程,然而迄今為止仍不清楚四倍體中脯氨酸代謝相關基因的進化模式及其與二倍體祖先脯氨酸代謝相關基因的關系。之前研究發現,油菜中含有6個脯氨酸合成途徑關鍵基因P5CS1,即該基因沒有發生丟失,并且在不同器官中與脅迫誘導情況下不同來源的P5CS1基因表達模式不同[21]。本研究集中探討了甘藍型油菜中脯氨酸降解代謝相關基因PDH1和PDH2的進化命運,包括二倍體親本進化上的來源、與二倍體親本物種的基因序列差異和重復基因是否發生丟失。研究結果能夠加深多倍化對脯氨酸代謝相關基因影響的理解,為多倍體表型和重復基因調控的關系提供理論依據。
1 基因序列分析
白菜和甘藍的相應基因序列由中國農業科學院蔬菜花卉研究所王曉武研究員提供,并且通過數據庫檢索和比對(http://www.brassica.info/; http://www.ocri-genomics.org/bolbase/index.html)。油菜基因序列來自于開放的油菜基因組序列網站(http://www.genoscope.cns.fr/brassicanapus/)。核酸序列分析應用DNAMAN軟件(Lynnon Biosoft,Vaudreuil,Quebec,Canada)。通過對同源基因對應位置的部分基因組DNA進行比對和序列分析,應用MEGA4軟件中鄰位相連法構建進化樹[22]。
2 結果與分析
2.1 油菜中脯氨酸合成代謝相關重復基因
2.1.1 白菜、甘藍、油菜中的PDH1基因概況 序列分析結果表明,白菜中存在3個PDH1的同源基因(http://www.brassica.info/)(ChrA2,Bra020731;ChrA6,Bra025421;ChrA9,Bra036196),甘藍中存在3個PDH1基因(http://www.ocri-genomics.org/bolbase/index.html)(Bol033129;Bol043056;Bol032297),甘藍型油菜中存在5個PDH1基因(http://www.genoscope.cns.fr/brassicanapus/) (ChrC7,NC_027773.1;ChrC5,NC_027771.1;ChrC5,NC_027771.1;ChrA9,NC_027765.1;unplaced genomic scaffold,NW_013650408.1)。根據系統命名法則[23],將白菜的3個PDH1同源基因分別命名為BraA.PDH1.a、BraA.PDH1.b、BraA.PDH1.c;將甘藍的3個PDH1基因分別命名為BolC.PDH1.a、BolC.PDH1.b、BolC.PDH1.c;將甘藍型油菜的5個PDH1基因分別命名為BnaA.PDH1.a、BnaA.PDH1.b、BnaC.PDH1.c、BnaC.PDH1.d、BnaC.PDH1.e。與擬南芥相同,蕓薹屬植物的PDH1基因也具有4個外顯子和3個內含子,除第1外顯子比擬南芥第1外顯子少3個堿基以外,其他外顯子堿基數目一致(表1)。蕓薹屬植物白菜、甘藍和甘藍型油菜的基因組序列(從ATG到終止密碼子)長度為2 201~2 671 bp,然而編碼區(CDS)長度一致,均為1 497 bp,所編碼的蛋白質都含有498個氨基酸。

表1 蕓薹屬PDH1基因結構對比
2.1.2 白菜、甘藍和甘藍型油菜中PDH2基因概況 基因組測序結果表明,白菜中存在1個PDH2基因(Bra028202),位于白菜A4號染色體上,命名為BraA.PDH2.a;甘藍中存在1個PDH2基因,命名為BolC.PDH2.a;甘藍型油菜中存在2個PDH2基因,分別命名為BnaA.PDH2.a、BnaC.PDH2.b。蕓薹屬植物PDH2基因cDNA編碼區長度為1 431 bp,含有4個外顯子3個內含子,和擬南芥一致(表2)。

表2 蕓薹屬PDH2基因結構對比
2.2 脯氨酸合成酶基因的同源性和進化關系
2.2.1PDH1基因的同源性和進化關系 為研究甘藍型油菜中的5個PDH1基因的二倍體祖先在甘藍型油菜染色體加倍過程中來自于白菜基因組還是甘藍基因組,將5個基因分別與白菜及甘藍的PDH1基因的基因組序列及編碼區序列做兩兩序列比對,結果如表2所示。同時,將甘藍型油菜、白菜、甘藍和擬南芥的同源PDH1基因的基因組DNA序列作多重比對,構建該基因的系統發生樹(圖1)。
序列相似性分析結果(表3)表明,BnaA.PDH1.a和BraA.PDH1.a、BnaA.PDH1.b和BraA.PDH1.c、BnaC.PDH1.c和BolC.PDH1.a、BnaC.PDH1.d和BolC.PDH1.b、BnaC.PDH1.e和BolC.PDH1.c之間的基因組DNA相似性分別為96.34%、99.67%、99.78%、98.48%、91.62%,而它們之間編碼區相似性更高,分別為99.20%、99.67%、99.80%、99.20%、99.53%,無論是基因組序列還是編碼區序列均高于同系列其他比對。因此,推斷BnaA.PDH1.a、BnaA.PDH1.b、BnaC.PDH1.c、BnaC.PDH1.d和BnaC.PDH1.e可能分別來

源于BraA.PDH1.a、BraA.PDH1.c、BolC.PDH1.a、BolC.PDH1.b和BolC.PDH1.c。而白菜的BraA.PDH1.b在甘藍型油菜中發生丟失。
而基于相對應的基因組序列的進化關系分析表明,BnaA.PDH1.a、BnaA.PDH1.b、BnaC.PDH1.c、BnaC.PDH1.d、BnaC.PDH1.e分別為白菜BraA.PDH1.a、BraA.PDH1.c和甘藍BolC.PDH1.a、BolC.PDH1.b、BolC.PDH1.c的直向同源基因(圖1)。
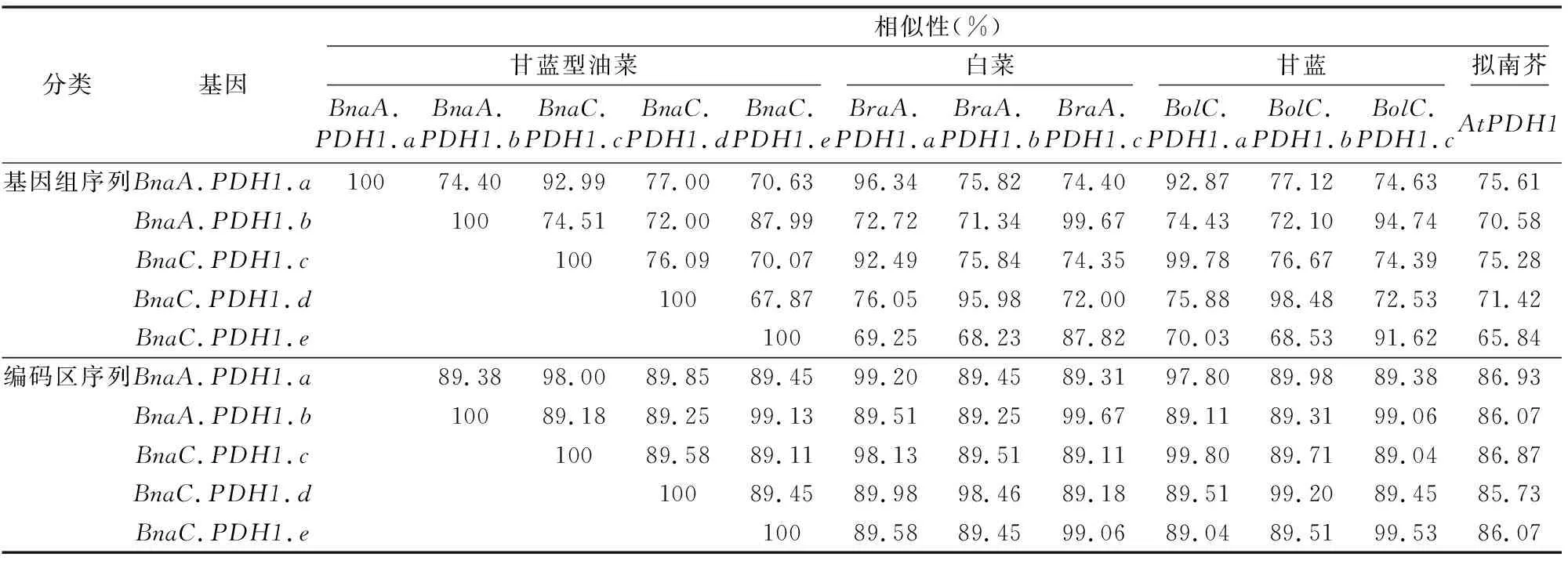
表3 甘藍型油菜PDH1基因的基因組序列和編碼區序列與二倍體親本白菜和甘藍PDH1基因序列相似性分析
2.2.2PDH2基因的同源性和進化關系 甘藍型油菜基因組中含有2個同源的PDH2基因,為了研究這2個基因的祖先來源,將2個基因分別與白菜及甘藍的PDH2基因基于基因組序列和編碼區序列做兩兩序列比對,比對結果如表4所示。同時,將甘藍型油菜、白菜、甘藍和擬南芥的所有已知序列的同源PDH2基因的基因組DNA序列作多重比對,構建該基因的系統發生樹(圖2)。
序列相似性分析結果表明,BnaA.PDH2.a與BraA.PDH2.a、BnaC.PDH2.b與BolC.PDH2.a基因組DNA序列相似性分別高達96.73%、99.67%(表4),而它們之間編碼區相似性更高,分別為99.86%、99.93%(表4),無論是基因組序列還是編碼區序列均高于同系列其他比對,推測BnaA.PDH2.a、BnaC.PDH2.b可能分別來源于BraA.PDH2.a、BolC.PDH2.a。
且基于相對應的基因組序列的進化關系分析也表明,BnaA.PDH2.a、BnaC.PDH2.b分別為白菜BraA.PDH2.a、甘藍BolC.PDH2.a的直向同源基因(圖2)。


表4 甘藍型油菜中PDH2基因的基因組片段與二倍體祖先中相對應的基因核苷酸序列相似性
3 討論
3.1 油菜的PDH1基因可能發生了1個拷貝的丟失
甘藍型油菜(B.napus)起源于白菜(B.rapa)和甘藍(B.oleracea)的雜交,并且經過了染色體加倍,而其祖先白菜和甘藍自身也經歷了基因組三倍化和重排的過程[16],這樣推測甘藍型油菜中每個基因同源基因的拷貝數最多可以達到6個。然而研究發現,由于基因丟失現象的存在,甘藍型油菜中同源基因的拷貝數平均為4個[20]。研究表明,甘藍型油菜中存在3個ALCATRAZ(ALC)基因[24]、3個GPAT4(sn-glycerol-3-phosphate acyltransferase 4)基因,然而存在的6個PSY(Phytoene Synthase)基因中有3個來源于白菜,另外3個來源于甘藍[25],而筆者之前的研究發現,甘藍型油菜中存在6個P5CS1基因[21]。本研究中,甘藍型油菜中存在5個PDH1基因,而其祖先白菜和甘藍中各存在3個PDH1基因,進化過程中甘藍型油菜可能發生了PDH1基因1個拷貝的丟失,而根據序列比對和進化分析結果,推測來自于二倍體祖先白菜的PDH1基因的1個拷貝發生丟失。而甘藍型油菜中存在2個PDH2基因,其祖先白菜和甘藍中各存在1個PDH2基因,甘藍型油菜中PDH2基因沒有發生丟失。
3.2 油菜中脯氨酸代謝相關基因與二倍體親本來源基因高度同源
根據白菜和甘藍全基因組測序結果,白菜和甘藍中PDH1和PDH2同源基因拷貝數都分別為3、1,而甘藍型油菜中拷貝數為5、2。序列比對結果表明,甘藍型油菜脯氨酸代謝途徑相關基因均與其親本白菜和甘藍來源的基因高度同源,PDH1和PDH2同源基因和親本來源基因的基因組DNA同源性分別為91.62%~99.78%、96.73%~99.67%。說明這些基因在甘藍型油菜進化過程中高度保守。這與之前的甘藍型油菜基因克隆的相關研究報道一致,甘藍型油菜中克隆得到3個PISTILLATA基因,兩兩序列相似性為96.49%~98.72%[26]。脯氨酸合成途徑上不同基因拷貝數的不同或許說明了這些基因在進化上的重要程度。
4 結論
以甘藍型油菜、白菜和甘藍為試驗材料,研究了甘藍型油菜中脯氨酸代謝途徑同源基因的進化命運,探討了多倍化對同源基因進化的影響。主要結論如下:通過白菜、甘藍和甘藍型油菜的基因組數據庫搜索和序列比對,確定了脯氨酸降解途徑關鍵基因(PDH1、PDH2)在甘藍型油菜中均存在多個拷貝,序列比對結果表明,它們和二倍體親本白菜及甘藍的對應基因高度同源;和親本相比,甘藍型油菜的PDH1基因來自于二倍體祖先甘藍的1個拷貝發生了丟失,而PDH2基因沒有發生丟失。
