粒用高粱4個主要光合性狀數(shù)量遺傳分析
2018-10-11 06:31:24盧華雨李延玲王惠茹
江蘇農(nóng)業(yè)科學(xué) 2018年17期
關(guān)鍵詞:模型
盧華雨, 李延玲, 羅 峰, 王惠茹
(天津農(nóng)學(xué)院農(nóng)學(xué)與資源環(huán)學(xué)院,天津 300384
高粱,在我國又被稱為“蜀黎”,屬于禾本科高粱族高粱屬,主要分布在亞洲、非洲、美洲等干旱和半干旱地區(qū),在我國的北方地區(qū)也大量種植。在世界范圍內(nèi)來看,高粱是僅次于小麥、水稻、玉米、大麥之后第五大糧食作物,也是世界上最重要的禾本科作物之一[1]。高粱具有抗鹽堿、耐高溫、耐寒、耐貧瘠、抗旱、抗?jié)车戎T多優(yōu)良性狀[2]。高粱作為C4植物,在高溫、強光、干旱、低二氧化碳的情況下仍然具有很高的光合效率[3]。高粱在人類發(fā)展的進程中也起到過重要的作物,也被稱為“救命谷”。我國是高粱的主產(chǎn)國之一,高粱為我國解決國民的溫飽問題作出了突出的貢獻。我國的高粱總產(chǎn)量僅次于美國位于世界第二。在我國,高粱在糧食作物栽培中,很早就已經(jīng)實現(xiàn)了“三系配套”,充分利用了高粱的雜種優(yōu)勢以及高光合效率,使高粱的單產(chǎn)大幅度的提高。但是我們也要注意,隨著世界人口的急劇增加以及可耕種土地的不斷減少,糧食危機很有可能再次出現(xiàn)[4]。目前已知高粱的最高產(chǎn)量是21 t/hm2,但這也僅僅是理論產(chǎn)量的56%,所以高粱的增產(chǎn)潛力是非常大的[5]。目前對于高粱的高光效遺傳育種方面的研究比較少見,主要集中在改變高粱冠層密度對于光合的影響[6]、種植密度對于高粱群體光合能力以及產(chǎn)量的影響[7]和高粱的株型性狀對于光合能力的影響[8],但對于高光效基因的遺傳育種研究較少。在其他主要作物上對于高光效育種研究比較深入,如在水稻方面有利用超級稻的生長優(yōu)勢的光合特征來提高水稻的光合性能[9]、水稻高光效育種[10]以及將高光效的基因?qū)胨緩亩岣吖夂闲蔥11]等幾個方面;在小麥方面有利用高產(chǎn)品種篩選出高光效品種[12]、小麥不同生育期光合效率的變化[13]以及不同供水條件對于小麥光合速率的影響[14]等諸多方面;蔬菜方面對高光效育種也有涉及如油菜[15]、甘藍[16]等。經(jīng)過對前人的研究發(fā)現(xiàn),高粱的株型[17]以及種植密度[18]對光合速率的影響非常大,因此想要增加高粱的生物產(chǎn)量可以改良高粱的株型以選育高光效品種。本研究對高粱光合性狀進行數(shù)量遺傳分析,以期從高粱的光合性狀的方向入手,通過提高高粱的光合速率、增加高粱的光合產(chǎn)物以增加高粱的產(chǎn)量,并結(jié)合分子輔助育種的手段,對控制相關(guān)性狀的基因進行定位挖掘,加快高粱育種進程,縮小育種年限,培育出高產(chǎn)優(yōu)質(zhì)的高粱品種。
1 材料與方法
1.1 試驗材料
試驗首先于2016年8月在天津市靜海縣良種場進行,選擇諸多高粱品種種植,每個品種隨機使用CRIAS-2便攜式光合儀測量光合速率,最終選擇光合能力較弱的品系引-20(美國普渡大學(xué)引進)與光合能力較強的品種忻粱52(國內(nèi)材料)作為親本,配制雜交組合獲得F1。2017年5月在天津市靜海縣播種,得到F2代植株209株,作為本次試驗群體。
1.2 性狀調(diào)查
選擇光照充足的天氣,每天09:00—11:30使用CRIAS-2便攜式光合儀測量高粱上數(shù)第2張葉的凈光合速率(Pn)、蒸騰速率(Tr)、氣孔導(dǎo)度(Gs)、細胞間CO2濃度(Ci)。
1.3 數(shù)據(jù)分析
通過光合儀獲得基本數(shù)據(jù)后,首先使用Excel作基本處理,再使用SPSS 19.2進行相關(guān)性分析。利用南京農(nóng)業(yè)大學(xué)蓋鈞鎰教授提出的單個分離世代群體的遺傳模型方法[19]進行光合數(shù)據(jù)的數(shù)據(jù)分析,可以得到F2群體11個遺傳模型以及每1個模型的AIC值。通過對11個模型的比較和篩選,選出4個AIC值最小的模型,隨后進行適合性檢驗[20]。通過檢驗篩選選出最適合的模型,然后根據(jù)一階分布參數(shù)[21],估計出一階遺傳參數(shù),并根據(jù)此參數(shù)計算出二階參數(shù),從而計算出主基因的遺傳率[22]。
2 結(jié)果與分析
2.1 性狀間相關(guān)性分析
高粱的高產(chǎn)以及物質(zhì)積累受到多種性狀的影響。其中植株的光合能力在高粱產(chǎn)量以及物質(zhì)積累形成階段起著非常重要的作用。植株葉片的凈光合速率、蒸騰速率、氣孔導(dǎo)度、細胞間的二氧化碳濃度直接影響著葉片的光合能力以及光合產(chǎn)物的積累,影響高粱植株最終的產(chǎn)量和干物質(zhì)含量。因此,為了了解葉片光合各個性狀之間的關(guān)系,對4個性狀進行了相關(guān)性分析。由表1可以得到,凈光合速率與蒸騰速率、氣孔導(dǎo)度、細胞間隙CO2濃度之間都呈現(xiàn)極顯著正相關(guān),其中,相關(guān)程程度最高的是氣孔導(dǎo)度與細胞間隙CO2濃度,相關(guān)系數(shù)為0.970,其次是蒸騰速率與細胞間CO2濃度,相關(guān)系數(shù)為0.966,相對來看,性狀之間相關(guān)程度最小的是凈光合速率與氣孔導(dǎo)度,相關(guān)系數(shù)為0.834。因此,在改良葉片光合能力的過程中,綜合考慮4個性狀之間的關(guān)系,經(jīng)過相關(guān)性分析可以得出結(jié)論,蒸騰速率、氣孔導(dǎo)度以及細胞間隙CO2提高會明顯增加高粱的凈光合速率。

表1 F2群體光合性狀相關(guān)性分析
注:“**”表示在0.01水平上(兩側(cè))極顯著相關(guān)。
2.2 光合相關(guān)性狀表型分析
忻粱52×引-20組合F2代群體光合性狀特征值、各個性狀的測定值(表2)以及各光合性狀頻次分布圖(圖1)表明,F(xiàn)2代凈光合速率、蒸騰速率、氣孔導(dǎo)度、細胞間CO2濃度均表現(xiàn)出了明顯的單峰現(xiàn)象,符合數(shù)量遺傳的分布特征。
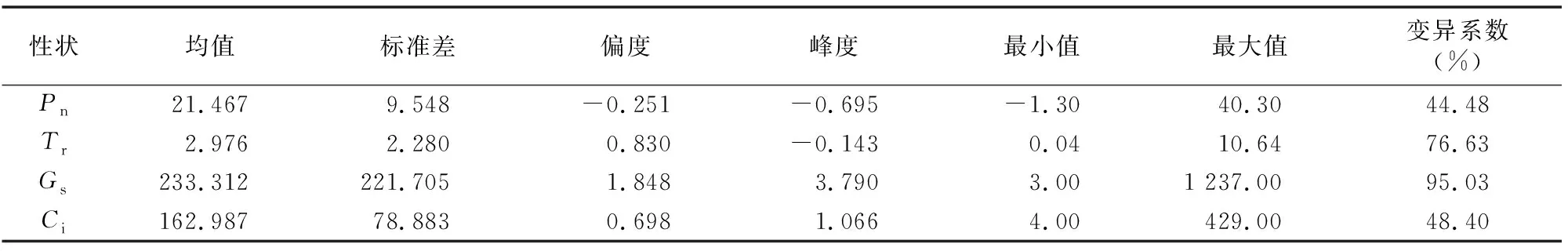
表2 忻粱52與引-20后代F2群體的光合性狀統(tǒng)計分析
2.3 光合性狀遺傳模型的選擇
利用主基因+多基因混合遺傳模型的F2單世代分析方法,對4個光合性狀進行遺傳分析(表3)。首先根據(jù)遺傳模型選擇的基本準則,即AIC值最小原則,每個性狀都要選擇AIC值最小的4個遺傳模型(表3中用*標注)作為遺傳分析備選模型。
2.4 光合性狀遺傳模型檢驗
利用U12、U22、U32(均勻性檢驗)、nW2(Smirnov檢驗)和Dn(Kolmogorov檢驗)對4個備選模型的F2世代進行適合性檢驗(表4),在4個模型中選擇出統(tǒng)計量達到顯著水平最小的1組模型作為最適模型。在凈光合速率4個備選模型中,模型B-2均勻性檢驗U12、U22、U32均未達到顯著水平(P>0.05),其他統(tǒng)計量達到顯著水平,并且模型B-2的AIC值最小。因此選擇模型B-2作為該群體凈光合速率最優(yōu)的遺傳模型,由此可以說明,凈光合速率的遺傳是受到2對主基因控制,并且主基因表現(xiàn)出加性-顯性的混合遺傳模型。
在蒸騰速率的4個備選模型中,模型B-2的U12、U22、U32(均勻性檢驗)、nW2(Smirnov檢驗)和Dn(Kolmogorov檢驗)均沒有達到顯著水平(P>0.05),并且模型B-2的AIC值最小。因此選擇模型B-2作為該群體蒸騰速率最優(yōu)的遺傳模型,由此可以說明,蒸騰速率的遺傳是受到2對主基因控制,并且主基因表現(xiàn)出加性-顯性的混合遺傳模型。
表3 凈光合速率、蒸騰速率、氣孔導(dǎo)度和細胞間隙CO2濃度遺傳模型的極大似然值和AIC值
在氣孔導(dǎo)度的4個備選模型中,模型B-1的U12、U22、U32(均勻性檢驗)、nW2(Smirnov檢驗)和Dn(Kolmogorov檢驗)均沒有達到顯著水平(P>0.05),并且模型B-1的AIC值最小。因此選擇模型B-1作為該群體氣孔導(dǎo)度最優(yōu)的遺傳模型,由此可以說明,蒸騰速率的遺傳是受到2對主基因控制,并且主基因表現(xiàn)出加性-顯性-上位性的混合遺傳模型。
在細胞間CO2濃度的4個備選模型中,模型B-1的U12、U22、U32(均勻性檢驗沒有達到顯著水平,P>0.05),nW2
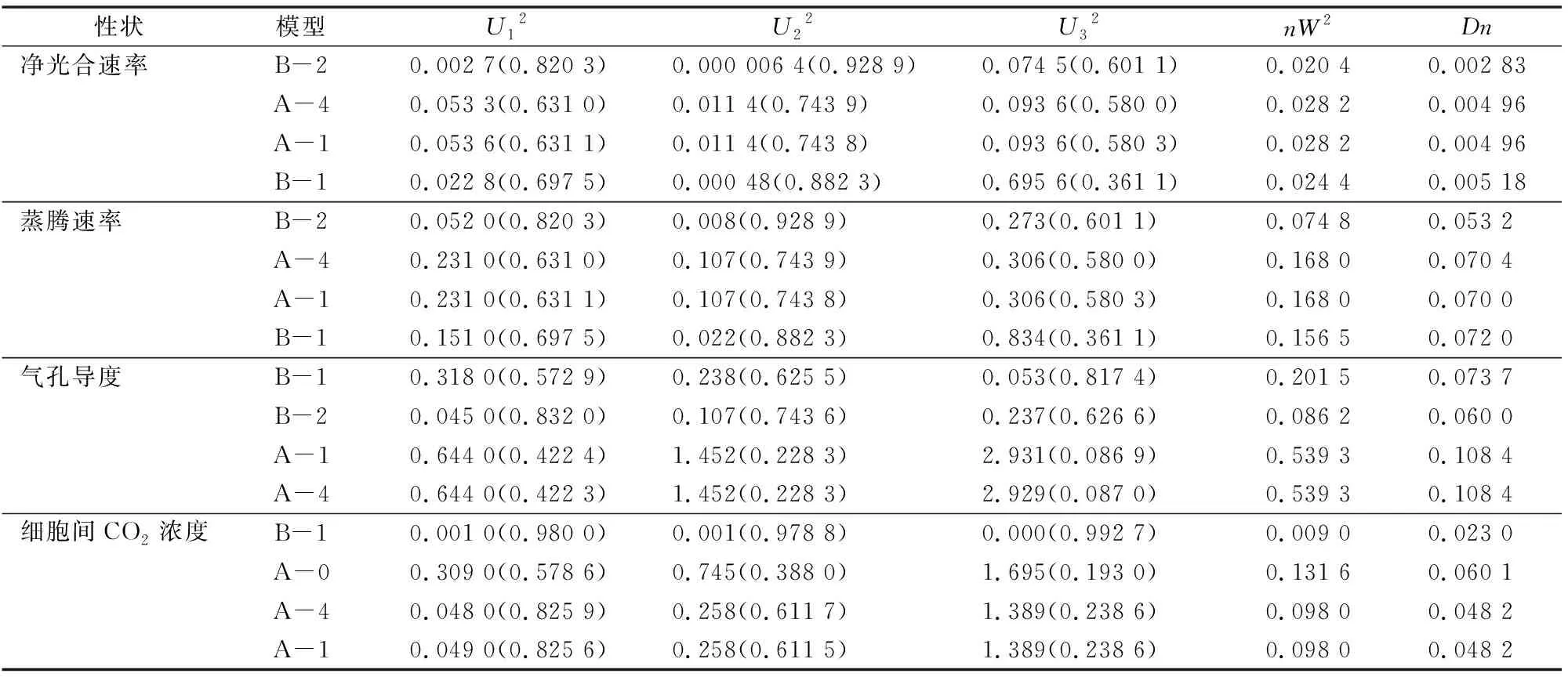
表4 光合各性狀遺傳模型分析檢驗
注:U12、U22、U32、nW2、Dn為適合性檢驗參數(shù),由主基因+多基因軟件計算得出。括號內(nèi)數(shù)字為適合性檢驗標準值。
(Smirnov檢驗)和Dn(Kolmogorov檢驗)達到顯著水平(P<0.05),并且模型B-1的并且模型B-1的AIC值最小。因此選擇模型B-1作為該群體氣孔導(dǎo)度最優(yōu)的遺傳模型,由此可以說明,細胞間CO2濃度的遺傳是受到2對主基因控制,并且主基因表現(xiàn)出加性-顯性-上位性的混合遺傳模型。
2.5 光合性狀的遺傳參數(shù)估計
根據(jù)之前確定了的遺傳最優(yōu)模型和IECM的估算方法,對F2群體的光合性狀進行遺傳參數(shù)估計(表5)。高粱的凈光合速率遺傳符合B-2遺傳模型,F(xiàn)2群體的遺傳分離符合加性-顯性的混合遺傳模型,2對主基因(AABB)的,加性效應(yīng)都表現(xiàn)出正值,2對主基因的加性效應(yīng)之和為11.75。2對主基因的顯性效應(yīng)都為正值,2對主基因的顯性效應(yīng)之和為1.748,凈光合速率的加性×加性的上位性為0.000 000 5。加性×顯性的上位性互作效應(yīng)值為-4.653,顯性×顯性上位性互作效應(yīng)值為-4.653。引-20與忻粱52雜交所得到的F2群體的凈光合速率的遺傳率為55.58%。說明控制高粱凈光合速率的相關(guān)基因狹義遺傳力較高,受環(huán)境影響較小,可以遺傳。高粱的蒸騰系數(shù)符合B-2遺傳模型,受到2對主基因控住,表現(xiàn)為加性-顯性混合遺傳模型,其中2對主基因的加性效應(yīng)值皆為正值,加性效應(yīng)值之和為3.307,2對主基因的顯性效應(yīng)值值為負值,顯性效應(yīng)值之和為-4.660。顯性效應(yīng)值明顯的小于加性效應(yīng)值,說明控制高粱蒸騰系數(shù)的2對主基因都以加性效應(yīng)為主,根據(jù)計算蒸騰系數(shù)的主基因遺傳率為85.04%。說明控制高粱蒸騰系數(shù)的相關(guān)基因狹義遺傳力很高,受環(huán)境影響小,可以穩(wěn)定的遺傳。
高粱的氣孔導(dǎo)度符合B-1遺傳模型,受到2對主基因控制,符合加性-顯性-上位性的遺傳模型,其中2對主基因的加性效應(yīng)值皆為正值,并且加性效應(yīng)值之和為384.953,2對主基因的顯性效應(yīng)值皆為負值,并且顯性效應(yīng)值之和為 -279.625,加性×加性上位性為192.176,加性×顯性的上位性互作效應(yīng)為-96.203,顯性×顯性的上位性互作效應(yīng)值為27.039。加性效應(yīng)明顯高于顯性效應(yīng),說明控制高粱氣孔導(dǎo)度的主基因以加性效應(yīng)為主,經(jīng)過計算得到高粱氣孔導(dǎo)度的遺傳率為79.94%。說明控制高粱氣孔導(dǎo)度的相關(guān)基因狹義遺傳力很高,受環(huán)境影響小,可以穩(wěn)定的遺傳。
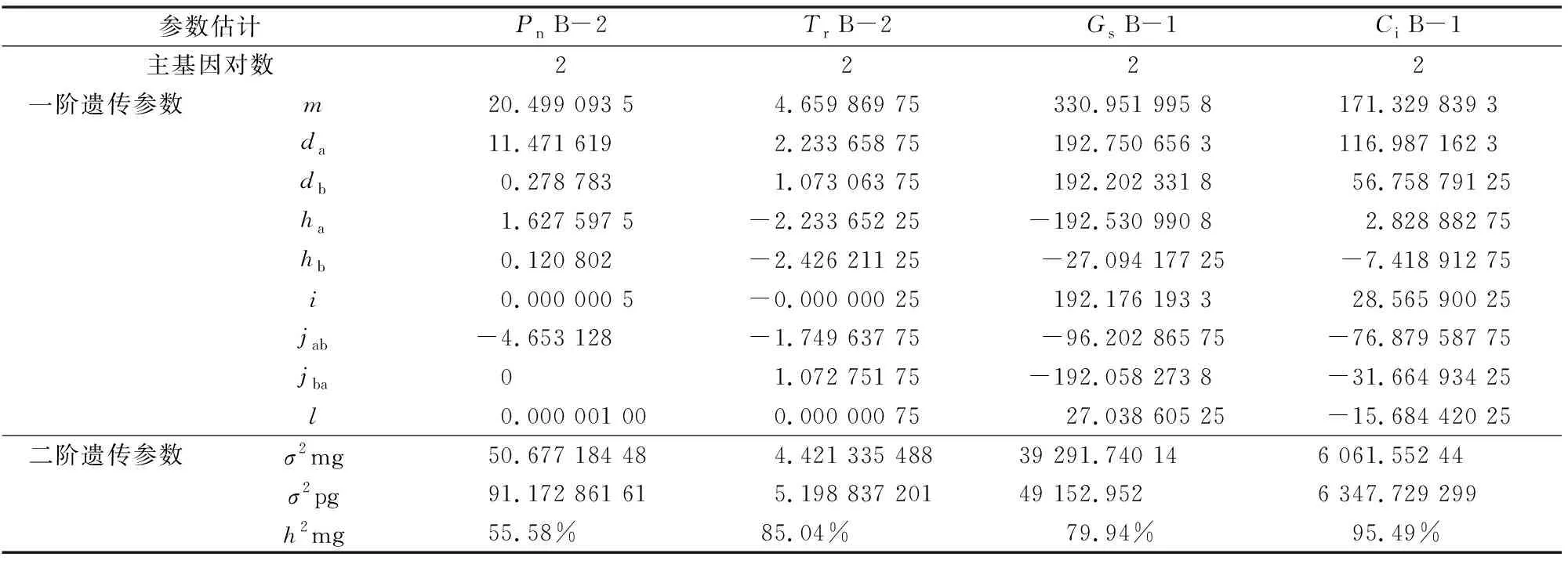
表5 高粱光合性狀遺傳參數(shù)估計
注:m為群體均方;da為第1對主基因的加性效應(yīng)值;db為第2對主基因的加性效應(yīng)值;ha為第1對主基因的顯性效應(yīng)值;hb為第2對主基因的顯性效應(yīng)值;i為2個主基因的加×加效應(yīng);jab為加性(a)×顯性(b)效應(yīng);jba為加性(b)×顯性(a)效應(yīng);l為2個主基因的顯×顯效應(yīng);σ2pg為多基因方程;σ2mg為主基因的方差;h2mg為主基因遺傳率。
高粱的細胞間CO2濃度的遺傳模型符合B-1模型,受到2對主基因控住,符合加性-顯性-上位性的遺傳模型,其中2對主基因的加性效應(yīng)值皆為正值,并且加性效應(yīng)值之和為173.746,2對主基因的顯性效應(yīng)值為1正1負,顯性效應(yīng)值之和為-4.59,加性×加性上位性的值為28.566,加性×顯性的上位性互作效應(yīng)為-76.880,顯性×顯性的上位性互作效應(yīng)值為-15.684,經(jīng)過計算,高粱細胞間CO2濃度的遺傳率為95.49%(表5)。說明控制高粱細胞間CO2濃度的相關(guān)基因狹義遺傳力很高,受環(huán)境影響小,可以穩(wěn)定的遺傳。
3 結(jié)論與討論
高粱的光合性狀是影響最終產(chǎn)量的重要因素,因此改良光合性狀對提高高粱的產(chǎn)量起著非常重要的作用。高光效育種在水稻上已經(jīng)取得重大進展,張建福等找到控制C4植物光合過程中固定CO2的關(guān)鍵酶丙酮酸磷酸雙激酶(PPDK),并將編碼玉米丙酮酸磷酸雙激酶(PPDK)基因的質(zhì)粒通過基因槍和農(nóng)桿菌介導(dǎo)轉(zhuǎn)化導(dǎo)入秈稻IR64中,在溫室條件下水稻的光合速率以及產(chǎn)量明顯高于對照組[23]。唐海濤等發(fā)現(xiàn)玉米雜交種單穗粒質(zhì)量最為相關(guān)的穗部光合性狀是穗位莖節(jié)長,其次是葉面積、葉長、光合效率和比葉重等,得出對玉米雜交種穗部產(chǎn)量性狀綜合影響較大的穗位光合性狀有穗位莖節(jié)長、葉長、比葉重等的重要結(jié)論[24]。其他一些學(xué)者在小麥[25]、甘藍[16]等農(nóng)作物上也取得了重大研究突破。
本研究以引-20與忻粱52雜交得到的F2群體為材料,對F2群體的光合性狀(包括凈光合速率、蒸騰系數(shù)、氣孔導(dǎo)度、細胞間CO2濃度4個性狀)進行了主基因+多基因的遺傳分析,結(jié)果表明,4個光合性狀都符合主基因+多基因的遺傳模型,存在著控制光合性狀的主效基因。其中凈光合速率受2對主基因控制,符合B-2模型,即加性-顯性的混合遺傳模型,主基因遺傳率為55.58%;蒸騰系數(shù)的遺傳受2對主基因的控制,符合B-2模型,即加性-顯性的混合遺傳模型,主基因遺傳率為85.04%;氣孔導(dǎo)度受2對主基因的控制,符合B-1模型,即加性-顯性-上位性的混合遺傳模型,主基因遺傳率為79.94%;細胞間CO2濃度受2對主基因控制,符合B-1模型,即加性-顯性-上位性的混合遺傳模型,主基因遺傳率為95.49%。因此,對高粱的光合性狀的改良,不僅能提高高粱的最終產(chǎn)量,還能提高高粱秸稈中干物質(zhì)含量[9],能夠推動高粱莖稈利用的多元化發(fā)展,并且有利于育種工作者在高粱育種過程中對光合性狀的選擇,了解控制光合性狀的基因效應(yīng)以及顯隱性關(guān)系,有助于進一步對控制光合性狀基因進行挖掘和篩選,大大減少了育種工作者的工作量,并提高了育種工作的成功率,在指導(dǎo)田間生產(chǎn)中具有重大的現(xiàn)實意義。
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網(wǎng)絡(luò)安全與數(shù)據(jù)管理(2022年1期)2022-08-29 03:15:20
導(dǎo)航定位學(xué)報(2022年4期)2022-08-15 08:27:00
中學(xué)生數(shù)理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數(shù)學(xué)備考)(2021年9期)2021-11-24 01:14:36
成都醫(yī)學(xué)院學(xué)報(2021年2期)2021-07-19 08:35:14
新世紀智能(數(shù)學(xué)備考)(2020年9期)2021-01-04 00:25:14
中學(xué)生數(shù)理化·七年級數(shù)學(xué)人教版(2020年10期)2020-11-26 08:24:50
數(shù)學(xué)物理學(xué)報(2020年2期)2020-06-02 11:29:24
光學(xué)精密工程(2016年6期)2016-11-07 09:07:19
