香菇原生質(zhì)體雜交后代的遺傳分析
2018-10-11 06:31:30劉俊杰
江蘇農(nóng)業(yè)科學(xué) 2018年17期
劉 娜, 張 敏, 宋 瑩, 劉俊杰
(遼寧省農(nóng)業(yè)科學(xué)院食用菌研究所,遼寧沈陽 110161
香菇(Lentinulaedodes)營養(yǎng)豐富,具有較高的保健價(jià)值,自古以來深受人們的喜愛[1]。中國是人工栽培香菇的發(fā)源地和世界香菇第一生產(chǎn)大國,目前,我國香菇年產(chǎn)量約為10萬t,占世界總產(chǎn)量的90%以上。香菇品質(zhì)的好壞與品種有很大關(guān)系,所以優(yōu)良香菇品種的選育尤為重要。
原生質(zhì)體雜交育種技術(shù)中的單核原生質(zhì)體是由雙核菌絲體中直接分離得來的,沒有經(jīng)過有性階段,一般認(rèn)為能更好地保持親本的性狀。利用這種原生質(zhì)體再生單核體,更容易定向選育優(yōu)良菌株[2]。同工酶分析作為分子遺傳標(biāo)記技術(shù)之一,不僅被廣泛應(yīng)用于動(dòng)植物的遺傳分析、生理生化研究,而且被作為化學(xué)分類的重要指標(biāo)應(yīng)用于真菌的分類鑒定中,特別是在屬內(nèi)種間及品種之間的分類鑒定中是非常有效的[3-4]。
在食用菌雜交育種中,親本的合理選配及雜種的準(zhǔn)確鑒定是至關(guān)重要的環(huán)節(jié)。多年來,育種工作者常常根據(jù)育種材料的生態(tài)型、生理特性及地理距離的差異選配親本。但是,這類差異與親本固有的遺傳差異往往并不完全一致。
本研究中的親本是基于酯酶同工酶技術(shù)、簡單重復(fù)序列區(qū)間(inter-simple sequence repeat,簡稱ISSR)分子標(biāo)記技術(shù),結(jié)合農(nóng)藝性狀選擇的4個(gè)菌株(168、931、荷香1號(hào)和野生香菇),制備原生質(zhì)體并單單雜交,采用酯酶同工酶技術(shù)對原生質(zhì)體單核化對稱雜交獲得的后代及其親本進(jìn)行遺傳分析,旨在鑒定雜交后代與親本的遺傳關(guān)系及雜交子的真實(shí)性。
1 材料與方法
1.1 試驗(yàn)材料
供試菌種及來源詳見表1。
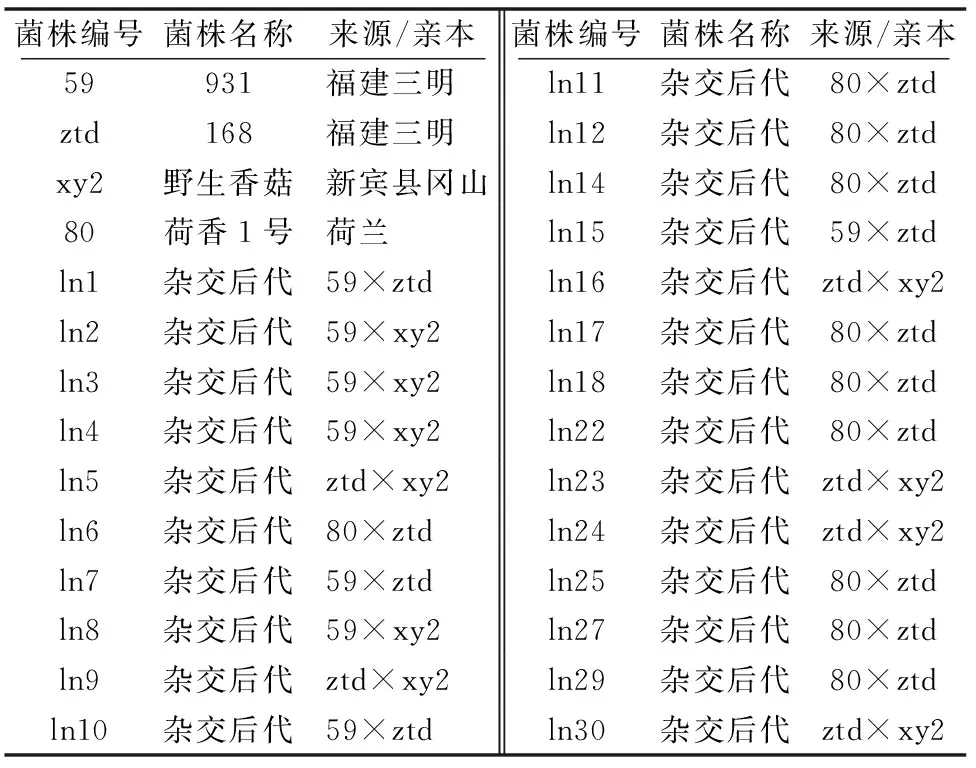
表1 供試香菇菌株及其來源
PDA綜合培養(yǎng)基配方:200 g馬鈴薯,20 g葡萄糖,20 g瓊脂,3 g磷酸二氫鉀,1.5 g硫酸鎂,3 g蛋白胨,1 000 mL水,pH值自然。
1.2 試驗(yàn)方法
1.2.1 雜交菌株獲得 獲得單核原生質(zhì)體雜交菌株的試驗(yàn)參照甘炳成等的方法[5]。
1.2.2 菌絲培養(yǎng) 將雜交親本及雜交后代菌株接于 25 mm×25 mm的試管內(nèi),每個(gè)菌株接10支,置于23 ℃恒溫培養(yǎng)15 d,刮取菌絲。
1.2.3 酯酶同工酶方法 采用垂直平板聚丙烯酰胺電泳法,收集供試菌株菌絲0.5 g/份,用液氮研磨,將樣品收集于 1.5 mL 離心管中,加入0.5 mL樣品提取液。將研磨后的樣品在4 ℃、12 000 r/min條件下離心20 min,取其上清液。將上清液、蔗糖溶液按體積比1 ∶1混合后作為電泳樣品。點(diǎn)樣量為40 μL,以溴酚蘭作為電泳指示劑,在4 ℃條件下電泳,初期電壓為每板150 V,當(dāng)溴酚蘭指示劑進(jìn)入分離膠后電壓升為每板250 V,待指示劑遷移至距末端1 cm左右處停止電泳,染色,保存?zhèn)溆谩?/p>
1.3 數(shù)據(jù)分析
聚類分析。相對遷移率(Rf)=X/Y。式中:X為從點(diǎn)樣孔下沿開始,酶帶在凝膠中的遷移距離;Y為從點(diǎn)樣孔下沿開始,指示劑在凝膠中的遷移距離。在測量遷移距離時(shí),以酶帶的中部位置為準(zhǔn)。依據(jù)酯酶酶譜,按照條帶的有無進(jìn)行標(biāo)記,在相同遷移率位置上,有條帶的記為1,無條帶的記為0。利用DPS數(shù)據(jù)處理軟件,分析得到供試菌株的聚類分析圖。
2 結(jié)果與分析
2.1 雜交親本與雜交后代酯酶同工酶圖譜
對酯酶同工酶圖譜經(jīng)過軟件Gel-Pro analyzer進(jìn)行條帶識(shí)別和人工審核的結(jié)果如下:由圖1-A、表2可知,親本59、xy2及其雜交后代共有遷移率各異的條帶13條。雜交后代ln3、ln4、ln8與親本有1條共同條帶,Rf為0.595,雜交后代有3條特異性條帶,Rf分別為0.351、0.402、0.571,說明雜交后代與親本之間有共源性,但雜交后代中又有屬于自己的特征酶帶,是有別于親本的新菌株。


表2 以59和xy2為親本的酯酶同工酶的相對遷移率
注:“—”表示沒有酶帶。下表同。
由圖1-B、表3可知,供試菌株親本ztd、59及其雜交后代的酯酶同工酶圖譜表現(xiàn)不同,酯酶同工酶酶譜帶數(shù)為5~10條,所有酶帶的Rf為0.171~0.804,6個(gè)菌株所共有的Rf為0.595、0.763,可以認(rèn)為是香菇的基礎(chǔ)酶譜帶,表明6個(gè)香菇品種之間有同源性,在主要代謝過程中具有相似的生理性狀。4個(gè)雜交后代在Rf為0.253、0.282、0.361、0.386、0.430、0.481、0.557、0.598、0.620、0.804的位置與親本存在差異,表明雜交后代雖然與親本菌株間有相同的酶帶,但是又有各自的特征酶帶,為新菌株。其中雜交后代中l(wèi)n15與其他雜交后代遺傳距離較遠(yuǎn),具有多條特征性條帶,說明ln15的變異幅度較大。
由圖1-C、表4可以看出,親本ztd、80與其雜交后代酶譜帶數(shù)和酶活性有較明顯的差異,其中共同的酶帶有2條,Rf分別為0.408、0.578。2個(gè)親本酶帶類型、數(shù)量、位置及寬度相差明顯,說明二者親緣關(guān)系較遠(yuǎn),為遠(yuǎn)源雜交[6];雜交后代酶帶類型、數(shù)量、位置及寬度相差較小,說明其親緣關(guān)系較近。Rf為0.170的是雜交后代共有而親本不具備的酶帶,可以看出,雜交后代有別于親本。
由圖1-D、表5可以看出,以ztd、xy2為親本,每個(gè)雜交后代與親本菌株有6~9條不等的酶帶。對供試菌株的遷移率分析表明,遷移率都在0.092~0.738之間。供試菌株在Rf為0.571、0.613這2處均有酶帶出現(xiàn),為雜交親本與后代共有的酯酶酶帶。雜交后代擁有的3條酯酶酶帶中,Rf為0.399、0.417的酶帶與親本ztd共有,Rf為0.595的酶帶與親本xy2共有,而Rf為0.455的酶帶為雜交后代區(qū)別于親本所特有的酶帶。

表3 以59和ztd為親本的酯酶同工酶相對遷移率
2.2 聚類分析圖
從圖2可以看出,931(59)和野生香菇(xy2)2個(gè)菌株遺傳距離較遠(yuǎn)。在相異系數(shù)為0.68時(shí)分為2類,第1類為ln3、ln4、ln8,第2類為59、xy2;雜交后代遺傳相異系數(shù)較小,在相異系數(shù)為0.51時(shí)聚為一類,但與2個(gè)親本的遺傳相異系數(shù)較大,在相異系數(shù)為0.85時(shí)聚為一類。由此可以看出,雜交后代是有別于2個(gè)親本的新菌株。
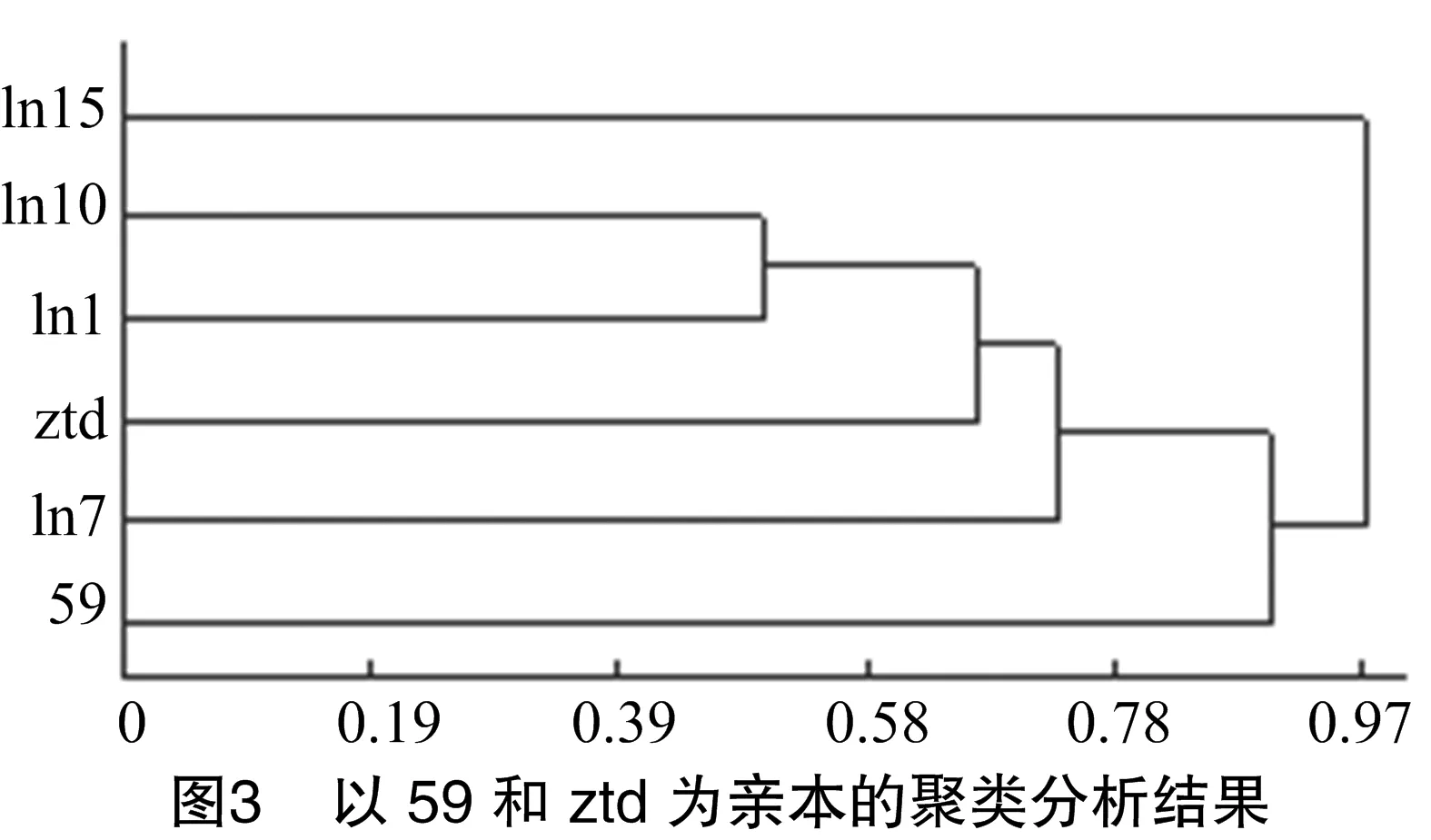
從圖3可以看出,以931(59)、168(ztd)為親本進(jìn)行雜交,931(59)和168(ztd)的親緣關(guān)系較遠(yuǎn),在相異系數(shù)為0.78時(shí)分為3類:第1類為ln15,第2類為ztd、ln1、ln7、ln10,第3類為59;雜交后代中l(wèi)n15與其他菌株的親緣關(guān)系較遠(yuǎn),其他幾個(gè)雜交后代親緣關(guān)系較近,在相異系數(shù)為0.73時(shí)雜交后代與親本ztd聚為一類,可以初步判斷,雜交后代遺傳性狀與ztd較為接近。在后期試驗(yàn)中發(fā)現(xiàn),ln1、ln7、ln10在生育期、菇體形態(tài)方面都接近親本ztd。
從以荷香1號(hào)(80)、168(ztd)為親本雜交后代的聚類分析結(jié)果可以看出,在遺傳相異系數(shù)為0.60時(shí),第1類為親本80,第2類為雜交后代與ztd。雜交后代中l(wèi)n18、ln22、ln25遺傳相異系數(shù)為0,ln11、ln12、ln14、ln17、ln27遺傳相異系數(shù)為0(圖4)。遺傳相異系數(shù)都為0的菌株說明它們具有共同的遺傳基因,為同一菌株。

表4 以80、ztd為親本的酯酶同工酶相對遷移率
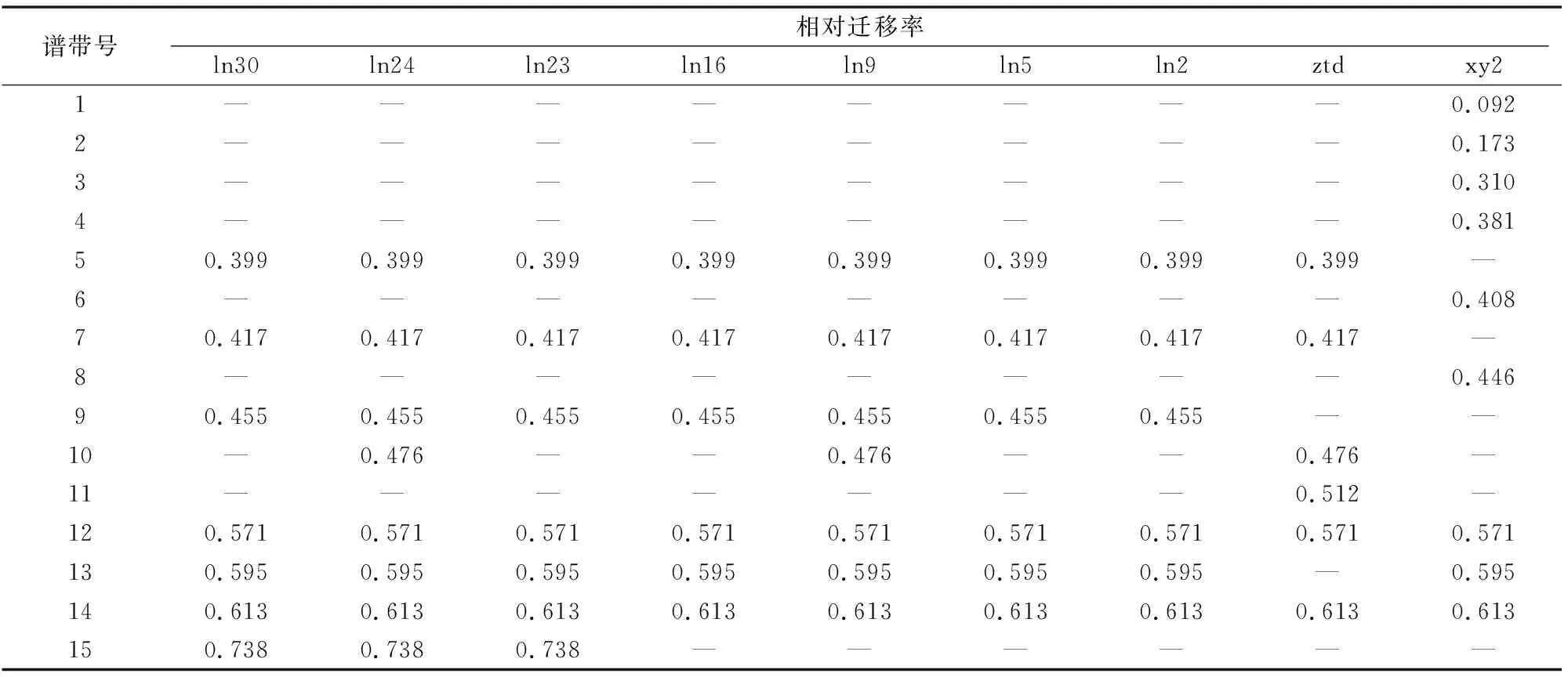
表5 以ztd和xy2為親本的酯酶同工酶相對遷移率


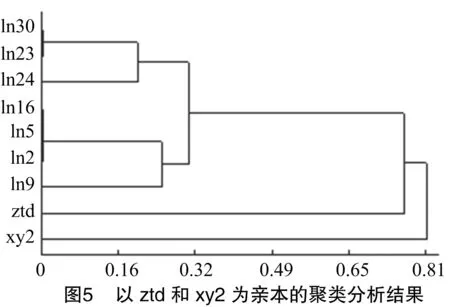
由以168(ztd)和野生香菇(xy2)為親本的聚類分析結(jié)果看出,xy2與其他菌株親緣關(guān)系較遠(yuǎn),單獨(dú)為一類;雜交后代中幾個(gè)菌株遺傳距離較近,在遺傳相異系數(shù)為0.32時(shí)聚為一類;ln23、ln30的遺傳相異系數(shù)為0,為同一菌株;ln2、ln5、ln16的遺傳相異系數(shù)為0,為同一菌株(圖5)。
3 結(jié)論與討論
本研究采用酯酶同工酶技術(shù),對雜交親本及其后代進(jìn)行遺傳差異研究,酯酶同工酶遺傳分析結(jié)果表明,供試菌株酯酶同工酶酶譜帶數(shù)為4~10條,遷移率在0.092~0.804之間,所有雜交后代與親本之間既有共同酶帶,又有屬于自己的特征酶帶,可初步認(rèn)為供試的24個(gè)雜交后代有別于親本。聚類分析結(jié)果表明,以931(59)、野生香菇(xy2)為親本的雜交群體,這2個(gè)親本菌株的遺傳距離較遠(yuǎn),在相異系數(shù)為0.68時(shí)分為2類,第1類為ln3、ln4、ln8,第2類為59和xy2;以931(59)、168(ztd)為親本進(jìn)行雜交,在遺傳相異系數(shù)為0.78時(shí)分為3類,第1類為ln15,第2類為ztd、ln1、ln7、ln10,第3類為59,雜交后代中l(wèi)n15與其他菌株親緣關(guān)系較遠(yuǎn);以荷香1號(hào)(80)、168(ztd)為親本,在遺傳相異系數(shù)為0.60時(shí)分為2類,第1類為親本80,第2類為雜交后代與ztd;以168(ztd)和野生香菇(xy2)為親本與雜交后代的聚類分析表明,野生菌株(xy2)與其他菌株親緣關(guān)系較遠(yuǎn),單獨(dú)為一類,雜交后代幾個(gè)菌株間遺傳距離較近。聚類分析結(jié)果也初步鑒定了雜交親本與后代之間的親緣關(guān)系,為以后的栽培試驗(yàn)提供了理論依據(jù)。供試的24個(gè)雜交后代均為真實(shí)雜交后代,結(jié)果與前期雜交后代親本拮抗呈陽性試驗(yàn)結(jié)果相符,說明酯酶同工酶技術(shù)可以作為鑒定雜交子真實(shí)性的標(biāo)準(zhǔn)之一。所有雜交后代中都具有特征酶帶,可作為判斷其他菌種是否為遼寧省農(nóng)業(yè)科學(xué)院食用菌研究所選育菌株的依據(jù)之一。從聚類分析圖可以初步看出雜交后代的遺傳距離趨近于哪個(gè)親本,至于農(nóng)藝性狀是否和聚類分析結(jié)果相符,還需要進(jìn)行栽培性狀比較試驗(yàn)。
中國香菇主栽品種遺傳背景相對狹窄,而野生資源存在豐富的遺傳多樣性,有意識(shí)地應(yīng)用野生種作為雜交親本,是一種有效擴(kuò)大栽培種遺傳多樣性的方法。為此,本研究將繼續(xù)利用野生資源進(jìn)行香菇新菌株選育,以保證香菇產(chǎn)業(yè)的可持續(xù)健康發(fā)展。
