綿羊Cytb基因生物信息學分析
2018-10-16 07:13:32臧勝芹陳曉勇孫洪新敦偉濤梁瑞圓
江蘇農業科學 2018年18期
臧勝芹,陳曉勇,楊 凌,孫洪新,敦偉濤,梁瑞圓
(1.河北省畜牧獸醫研究所,河北保定071000;2.河北工程大學生命科學與食品工程學院,河北邯鄲056038)
線粒體DNA(mitochondrial DNA,mtDNA)具有分子量小、進化速度快、母性遺傳等特點,因而mtDNA是研究分子進化的重要材料[1-2]。mtDNA的復制、轉錄及蛋白質合成均依賴于核基因編碼的各類蛋白質因子的調控[3]。細胞色素 b(cytochrome b,Cytb)是由mtDNA 13個多肽編碼基因之一,編碼產物為線粒體氧化磷酸化系統復合體Ⅲ組成之一,也是其中唯一由線粒體基因組編碼的蛋白質[4],是線粒體呼吸鏈上進行電子傳遞的細胞色素之一。細胞色素b的結構特征是都以血紅素b作為輔基而且并不通過卟啉環邊基與蛋白鏈共價結合。研究得比較多的只有細胞色素b5(Cytb5),它是一種膜蛋白質。根據晶體X射線分析推出,肽鏈組裝成一個桶狀構象把血紅素部分盛在“桶”中:幾片β片組成桶底,4個短的α螺旋(α1、α3、α4及 α5)構成四壁[5]。Cytb 基因廣泛應用于分子進化和系統發育研究,但有關綿羊Cytb基因編碼產物的研究很少。
為此,本研究通過測定湖羊Cytb基因CDS區序列,利用生物信息學方法,進行不同物種Cytb基因系統發育分析,對綿羊Cytb基因CDS區序列蛋白質的理化性質、二級結構及多參數預測、蛋白質跨膜結構和信號肽預測、亞細胞定位和蛋白質磷酸化、功能結構域、三級結構等進行分析,旨在為綿羊Cytb基因結構與功能研究提供參考。
1 材料與方法
1.1 Cytb基因序列來源
采集湖羊血液,提取DNA,參照文獻[6]合成引物擴增Cytb基因,將PCR產物測序獲得該基因序列。具體如下:上游引物:5'-GTCATCATCATTCTCACATGGAATC-3';下游引物:5'-CTCCTTCTCTGGTTTACAAGACCAG-3'。產物序列長度1 272 bp。由上海生工生物工程有限公司合成。PCR反應體系 30 μL:DNA 2.4 μL,10 × PCR Buffer 3 μL,dNTP(10 mmol/L)0.6 μL,上、下游引物(10 mmol/L)1.2 μL,Taq DNA Polymerase(5 U/μL)0.18 μL,ddH2O 21.42 μL。PCR反應條件:95℃預變性5 min;95℃變性20 s,退火30 s,72℃延伸90 s,35個循環;72℃延伸7 min。將擴增產物使用1.0%的瓊脂糖凝膠電泳檢測,120 V電泳35 min,使用凝膠成像儀檢測結果后,送上海生工生物工程有限公司進行雙向測序。并利用DNAman軟件對測得的湖羊Cytb基因進行翻譯獲得氨基酸序列。
此外,從NCBI網站的GenBank下載山羊、家牛、野豬、家犬、家馬、小家鼠、家貓、黑猩猩、大猩猩、人、原雞、斑馬魚等12個物種31條相似度較高的(一般為80%以上)Cytb基因CDS及其對應的氨基酸序列(表1)。
1.2 分析方法
使用 NCBI網站中的(http://www.ncbi.nlm.nih.gov/gorf/orfig.cgi)查找開放閱讀框功能,利用 BioEdit Sequence Alignment Editor對32條核苷酸序列和與之對應的32條氨基酸序列進行Fas格式轉換,利用NCBI網站Blast軟件進行核苷酸和氨基酸的同源性比對分析;利用Clustal X[7]和MEGA 6.0[8]軟件的鄰接法(neighboe - joiningmethod,NJ)構建系統發育樹;利用在線軟件(http://web.expasy.org/protparam/)以及DNAStar里的Protean分析蛋白質理化性質;利用JPred4在線軟件預測二級結構;利用DNAStar程序包里Protean軟件分析親水性、柔韌性、抗原指數和氨基酸表面可及性等參數;利用 TMHMM 在線軟件(http://www.cbs.dtu.dk/services/TMHMM/)進行蛋白質跨膜結構分析;利用SignalP在線軟件(http://www.cbs.dtu.dk/services/SignalP - 4.0/)進行蛋白質信號肽分析;利用 PSORT進行亞細胞定位分析;利用NetPhos 2.0 Server(http://www.cbs.dtu.dk/services/NetPhos/)對Cytb基因編碼蛋白質進行磷酸化位點預測;利用SWISSMODEL預測蛋白質的三級結構。

表1 12個物種的Cytb基因序列來源
2 結果與分析
2.1 綿羊Cytb基因開放閱讀框分析
通過ORF Finder分析表明,綿羊Cytb基因的ORF長度為1 140 bp,編碼379個氨基酸。
2.2 綿羊Cytb基因序列同源性及分子進化分析
通過NCBI進行同源性在線分析,湖羊、人、山羊、家牛、斑馬魚、家馬、家貓、原雞、大猩猩、小家鼠、黑猩猩、野豬和家犬的13個物種Cytb基因序列以及氨基酸序列進行比對,結果顯示,湖羊與山羊的相似性最高,其次是家牛,最后與原雞和斑馬魚一致性最低(表2)。系統發育樹顯示,湖羊和山羊親緣關系最為接近,其次是家牛,與原雞、斑馬魚等親緣關系較遠(圖1)。這與核苷酸一致性分析一致,符合物種進化規律。
2.3 綿羊Cytb基因編碼蛋白質結構分析
2.3.1 理化性質分析 綿羊Cytb基因編碼蛋白質由379個氨基酸殘基組成,其分子式為C3583H6028N1140O1451S324分子量為98.68 ku,理論等電點(pI)為5.0,說明該蛋白質為酸性蛋白質。其不穩定系數為64.20,說明該蛋白質為不穩定蛋白質,且預計在體外哺乳動物的網織紅細胞內半衰期為4.4 h,在酵母體內的半衰期>20 h,在大腸桿菌體內>10 h,脂溶指數為31.49,總平均疏水指數1.035,屬于疏水蛋白質。含有20種常見氨基酸殘基(圖2),其中Leu(13.98%)含量最高,其次Ile(11.87%)含量較高。帶負電荷的氨基酸殘基(Asp+Glu=0)等于帶正電荷的氨基酸殘基(Arg+Lys=0)。其摩爾消光系數為 88 850 L/(mol·cm),1A(280)=0.48 mg/mL,等電點是 7.52,pH 值為 7 時,帶電量為 1.78。

表2 綿羊Cytb基因核苷酸和氨基酸序列與其他動物的一致性分析


2.3.2 Cytb結構蛋白質二級結構及多參數分析 蛋白質二級結構是指蛋白質分子中多肽鏈本身的折疊方式,蛋白質分子的多肽鏈一般是部分卷曲盤旋成螺旋狀(α-螺旋結構),或折疊成片層狀(β-折疊結構),或以不規則卷曲結構存在于生物體內[9]。
Gamier_Robson方法是通過計算特定氨基酸殘基在特定結構內部的可能性來預測蛋白質的二級結構的。用該方法預測Cytb結構蛋白質二級結構,發現它有α螺旋形成,同時有許多β折疊存在,且分布比較均勻,在各β折疊單元之間存在長短不一的轉角(圖3)。該方法預測的結果還顯示,在該蛋白質的跨膜區有一些α螺旋結構形成。
Chou_Fasman方法是通過序列氨基酸殘基的晶體結構來預測二級結構,用該方法預測Cytb結構蛋白質二級結構,發現它有α螺旋結構形成,其位置分別在第His 8-Val 14、lys 172-Glu 202、Gln 312-Met 316和 Gly351-Trp379區段(圖3)。相應地,用該方法預測的β折疊和轉角結構就較少且主要處于該蛋白質的兩端。

組成Cytb蛋白質二級結構的α螺旋(HELIX,H)、β折疊(STRAND,E)、無 規 卷 曲 (LOOP,L) 的 比 例 為64.91 ∶0.79 ∶34.30(圖4)。

Cytb結構蛋白質的親水性分析:用Kyte_Doolittle方法對Cytb結構蛋白質的親水性進行了分析。結果顯示該蛋白質存在較少的親水性區域,但其中親水性較高的區域主要集中在:Glu 202-Thr 225、Leu 249-Glu 271和 His 308-Arg 318提示該區域暴露于表面的幾率較大,作為抗原表位的可能性也最大(圖5)。
Cytb結構蛋白質的表面可能性分析:在表面可能性較大的區域主要是 Ile 4-Met 11、His 32-Thr 60、Thr 203-Lys 227、Leu 249-Glu 271和Thr 309-Met 315區段,其他部位展示的可能性較小(圖5)。
Cytb結構蛋白質骨架區的柔韌性分析:含有一些的柔韌性區域,且分布比較均勻,提示該蛋白質肽段有一定的柔韌性,可以發生扭曲、折疊,能形成比較豐富的二級結構(圖5)。
Cytb結構蛋白質B細胞抗原表位的預測分析:通過聯合以上蛋白質結構預測方法分析Cytb結構蛋白質潛在的蛋白質抗原決定簇,結果顯示結構蛋白質含有一些抗原指數較高的區域,其中以羥基端的指數性最高,所跨區域也最大(Asp 248-Glu 271),提示該區段含有潛在優勢抗原表位,其他一些區段也可能含有一些潛在的抗原表位,如,Thr 203-Lys 227等,這些區域的抗原指數也比較高,蛋白質抗原指數較高的區域與其親水性較高區域的分布較為一致,這些區域可能包含潛在優勢的抗原表位(圖5)。

2.3.3 蛋白質跨膜結構和信號肽分析 從圖6可以看出,Cytb蛋白質共存在9個典型的跨膜螺旋區,氨基酸序號分別為33 -55、76-98、113-135、140-158、178 -200、229 -251、288-310、323-340和350-372。由此可推測,綿羊Cytb基因CDS區所編碼的蛋白質為跨膜蛋白質,提示它可能作為膜受體起作用,也可能是定位在膜上的錨定蛋白質或離子通道蛋白質。

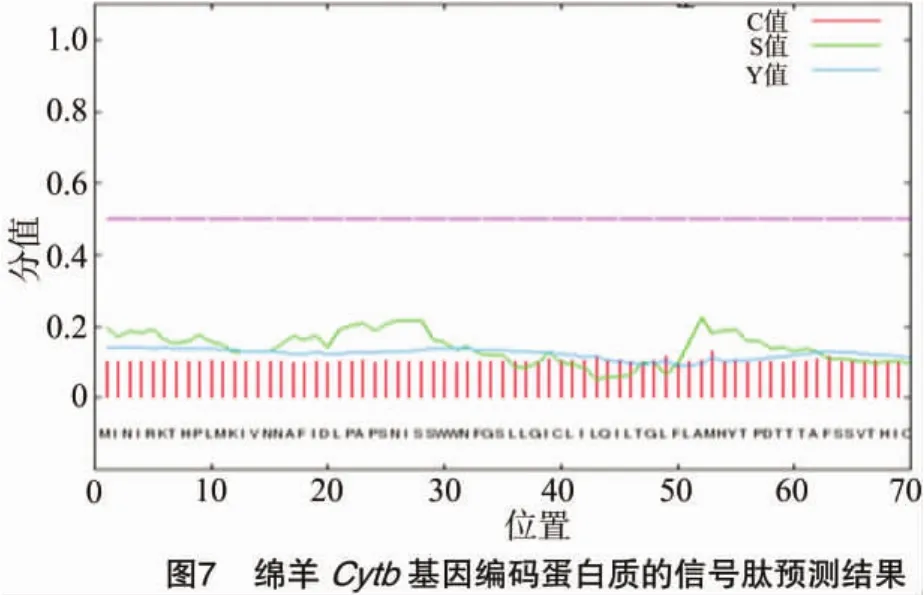
從圖7可以看出,Cytb蛋白質的C、Y和S值的計算結果不具備信號肽的要求,表明綿羊Cytb蛋白質不存在信號肽故該蛋白質為非分泌蛋白質。
2.3.4 亞細胞定位和蛋白質磷酸化分析 亞細胞定位可以明確某種蛋白質或表達產物在細胞內的具體存在部位。經亞細胞定位分析表明,該序列主要在內質網(77.8%)、細胞核(11.1%)、線粒體(11.1%)內發揮其生物學作用。蛋白質磷酸化是調節和控制蛋白質活力和功能的重要機制。該蛋白質經磷酸化分析顯示(圖8),綿羊Cytb蛋白質有較多的蘇氨酸(Thr)以及絲氨酸(Ser)磷酸化位點和相對較少的酪氨酸(Tyr)磷酸化位點。

2.3.5 高級結構預測與分析 利用SWISS-MODEL預測蛋白質的三級結構,結果(圖9)顯示,綿羊Cytb蛋白質序列與Blast數據庫中5j4z.53.A模板序列相似性高達61%,該蛋白質存在較多的扭曲,結構較為豐富,該結果同時也表明了綿羊Cytb由α-螺旋、β-折疊和無規則卷曲組成,與二級結構預測一致。而這些結構對其生物學功能的發揮有重要作用。

3 討論
在線粒體DNA(mtDNA)的13個蛋白質編碼基因中,Cytb的結構功能最為清楚[10-11]。Cytb主要存在于內膜磷脂中,在線粒體蛋白質編碼基因中有著十分重要的作用,在氧化磷酸化過程中作為電子傳遞的重要媒介,其自身可以在線粒體內進行轉錄和翻譯。Cytb基因的進化速度相對來說比較適中,通常被人們用來研究種內、近緣種間以及種間親緣關系,成為一種重要的區分手段[12]。在對不同物種同源性及系統發育分析可知,湖羊與山羊親緣關系最近,其次是與牛親緣關系最近,其中偶蹄目最先聚為1支,再與靈長類動物聚在一起,最后與原雞、斑馬魚等聚在一起。綿羊Cytb基因共編碼379個氨基酸。
常見的蛋白質二級結構有α-螺旋、β-折疊、轉角和無規則卷曲等。綿羊的Cytb基因編碼蛋白質以α-螺旋和無規則卷曲為主。蛋白質分子構象主要靠非共價鍵維持,蛋白質結構的穩定在很大程度上有賴于分子內的疏水作用,從各種因素的作用來看,疏水作用是非常重要的[13]。經分析綿羊Cytb基因編碼蛋白質為酸性、不穩定性疏水蛋白質。一般信號肽位于分泌蛋白的N端,通過判斷蛋白質是否含有信號肽部分,可以初步推測該蛋白質是否為分泌蛋白[14],結果分析表明,Cytb蛋白質既不存在信號肽,也不是分泌蛋白質。綿羊Cytb基因編碼蛋白質有9個跨膜結構域,這與劉安芳等對家鵝線粒體細胞色素b跨膜螺旋結構分析得出的結果[15]一致。Cytb主要在內質網、細胞核、線粒體內發揮其生物學作用,與線粒體DNA的各種功能和調節有著密切的關系。蛋白質磷酸化的表達直接影響蛋白質活力和功能的調節,是最重要的機制,綿羊Cytb蛋白質的蘇氨酸和絲氨酸磷酸化位點比較多,酪氨酸的磷酸化位點相對較少,蘇氨酸和絲氨酸磷酸化的主要作用是激活蛋白質的活力,結果表明該蛋白質酶活力較強[16]。
猜你喜歡
哲學評論(2021年2期)2021-08-22 01:53:34
民用飛機設計與研究(2020年4期)2021-01-21 09:15:02
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
電子制作(2018年18期)2018-11-14 01:48:24
山東工業技術(2016年15期)2016-12-01 05:31:22
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
現代企業(2015年9期)2015-02-28 18:56:50
中國中醫藥現代遠程教育(2014年11期)2014-08-08 13:23:44
終身教育研究(2014年5期)2014-02-28 01:23:06
