冰鮮鴿肉貯藏過程中的微生物菌群多樣性
2018-10-25 03:29:26蔡教英王小玉姚麗鋒游淑珠陳海光朱楚雯
肉類研究 2018年9期
蔡教英,王小玉,姚麗鋒,游淑珠,丁 琦,陳海光*,朱楚雯
(1.珠海檢驗檢疫技術中心,廣東 珠海 519015;2.仲愷農業工程學院輕工食品學院,廣東 廣州 510225)
“一鴿勝九雞”的俚語說明鴿子的營養成分非常豐富,比雞更勝一籌。粵港澳等地區素來有食用乳鴿的飲食習慣,烹調方法多種多樣,市場對鴿肉的需求也越來越大。但近年來,禽流感疫情嚴重影響了整個家禽產業的發展,傳統的活禽銷售模式對消費者及養殖戶都潛在巨大危害。隨著2014年以來廣東省倡導的“集中屠宰、冷鏈配送、生鮮上市”的家禽屠宰政策的實施,改變禽類食品供應模式,以冰鮮替代活禽供應,對建立防控人感染禽流感的長效機制和促進產業轉型升級均具有重要意義。
鴿肉營養物質豐富且水分活度高,這使得鴿肉在生產、貯藏、運輸及銷售等過程中易受微生物的污染,且在不同貯藏條件下,冰鮮肉的微生物多樣性存在一定差異[1]。冰鮮鴿是在乳鴿屠宰后將衛生檢驗合格的乳鴿胴體迅速冷卻至0~4 ℃,并在此溫度下對動物胴體進行加工、貯運和銷售[2]。冰鮮鴿肉在0~4 ℃低溫條件下貯藏,能夠有效抑制肉中微生物的生長和酶活性,降低細菌總數,減少有害毒素的產生,使冰鮮肉中的致病菌、生物毒素受到一定程度的抑制,延緩肉類腐敗變質[3]。目前,關于冰鮮肉類的腐敗微生物研究大部分集中在雞肉[4-6]、豬肉[7]、蝦[8]、牛肉[9]、羊肉、鴨肉、魚[10-11]、貝等。張莉等[12]采用轉錄間隔區序列(internal transcribed spacer,ITS)測序,大部分研究采用16S rDNA測序。孫彥雨等[13]應用16S rDNA的變性梯度凝膠電泳指紋圖譜技術對冰鮮雞肉的微生物多樣性進行研究。唐文靜等[14]使用16S rDNA分子鑒定對冷藏海鱸魚的優勢腐敗菌進行研究。岑璐伽等[15]使用聚合酶鏈式反應(polymerase chain reaction,PCR)-變性梯度凝膠電泳分析冷卻牦牛肉貯藏過程中的優勢菌。
近年來,隨著測序技術的發展,可以進行大規模平行測序,且能客觀反映樣本中DNA的豐度和鑒定出傳統培養條件下不可生長細菌的高通量測序技術應用越來越廣泛,而且較傳統測序法成本低廉[16]。與傳統方法相比,細菌的高通量測序技術在微生物群落研究方面的優勢越來越突出,目前已廣泛應用于醫學[17]、環境[18-20]、昆蟲學[21-23]、中藥[24-25]及植物[26-27]等多個方面。且近幾年來在食品領域的應用發展迅速,已廣泛應用于乳制品[15]、發酵類食品[16,28]、肉制品[4-7]、水產品[8,10-11]中的細菌群落多樣性研究。
本研究采用高通量測序技術對冰鮮鴿肉中的腐敗菌進行研究,分析不同增菌溫度下冰鮮鴿肉中微生物菌群的菌相構成情況、貯藏過程中各種微生物菌群數量的變化特點以及貯藏過程中肉質新鮮度的變化規律。為了提高DNA的得率,需對細菌進行增菌處理。目前的國家強制性標準中測定菌落總數時的培養溫度為(36±1) ℃,但是該條件下培養時的菌落總數測定對象只包括嗜溫性需氧微生物。對于低溫貯藏冷鮮肉來說,嗜冷菌也可能是優勢菌群,如果能對嗜冷菌和嗜溫菌進行同時檢測,綜合評價會更合理。因此,本研究選取4 ℃和37 ℃ 2 種增菌溫度,綜合分析得出冰鮮鴿肉的微生物消長規律,了解其菌相變化和品質變化的規律,以期為冰鮮鴿的保鮮控制技術提供理論基礎。
1 材料與方法
1.1 材料與試劑
冰鮮鴿肉由珠海市金同農副食品有限公司提供,隨機選取18 只當天屠宰的冰鮮鴿密封包裝后2 h內用冰盒運送至實驗室。在無菌環境下將鴿肉隨機分為9 組,每組2等份,分別收集每只冰鮮鴿的鴿肉作為樣品,每份25 g,經取樣袋密封包裝后放置于4 ℃冰箱貯藏。
Q5413細菌基因組DNA提取試劑盒 德國Tiangen公司;10855-021 LB肉湯 美國Termo Fisher Scientific公司;M0491L擴增聚合酶 美國NEB公司;P7589熒光定量染料、75510-019瓊脂糖凝膠試劑、AM9870瓊脂糖凝膠電泳緩沖液 美國Invitrogen公司;DL2000核酸Marker 日本TaKaRa公司。
1.2 儀器與設備
2100生物分析儀 美國Agilent公司;KB400恒溫培養箱 德國Binder公司;Minispin小型高速離心機德國Eppendorf公司;LogicA2生物安全柜 美國Labconco公司;Synergy2全波長酶標儀 美國Biotek公司。
1.3 方法
1.3.1 細菌總DNA提取
以購買當天為0 d,作為對照組,其余為實驗組。在無菌環境下,分別取貯藏不同時間(0、2、4、6、8、10、12、14、16 d)的冰鮮鴿肉25 g裝入拍打袋,加入225 mL無菌生理鹽水,用拍擊式均質器拍打后,移取1 mL菌液加入40 mL的LB培養基中,于4 ℃條件下增菌7 d,同時取1 mL菌液加入40 mL的LB培養基中,于37 ℃條件下增菌24 h。分別取1 mL增菌液至EP管內,采用Tangen細菌基因組DNA提取試劑盒按照相應操作步驟進行。
1.3.2 16S rDNA V3-V4區PCR擴增
引物序列:上游5’-A C T C C T A C G G G A GGCAGCA-3’,下游5’-GGACTACHVGGGTWTCTAAT-3’。對細菌16S rDNA基因V3-V4可變區進行PCR擴增。反應體系(25 μL):2×Mix 12.5 μL、引物(10 μmol/L)1 μL、DNA模板(30 ng)1 μL、ddH2O 10.5 μL。98 ℃預變性45 s;98 ℃變性15 s,57 ℃退火30 s,72 ℃延伸1 min,循環20 次;72 ℃末端延伸10 min,4 ℃條件下保溫。
1.3.3 測序文庫制備及數據分析
高通量測序文庫的構建和Ion Torrent PGM高通量測序的工作委托上海伯豪生物公司進行。按97%相似性標準進行操作分類單元(operational taxonomic unit,OTU)聚類,然后利用RDP-classifier及Silva數據庫進行物種注釋和分類學分析。基于OTU的分析結果,采用對樣本序列進行隨機抽樣的方法,計算常用的Alpha多樣性指數,包括超Ⅰ指數、Ace指數、香農指數(Shannon index)和辛普森指數(Simpson index),并作出相對應的稀釋曲線。超Ⅰ指數和Ace指數用于評價微生物菌群豐富度,其數值越高,表明群落物種豐富度越高;香農指數和辛普森指數反映物種多樣性,香農指數與群落多樣性呈正相關,辛普森指數則相反,其值越大,群落多樣性越低。通過多變量統計學方法的主成分分析(principal component analysis,PCA),可以直觀顯示不同樣品群落間物種組成的相似性及差異性[14]。
2 結果與分析
2.1 稀釋性曲線結果分析
為避免在高通量測序過程中由于引入錯誤或不可靠堿基等不可控因素造成的測序錯誤,減少對下游生物信息分析造成的不良影響,需要對測序得到的原始數據進行優化處理。通過利用QIIME軟件對測序所得原始序列數進行過濾,去除低質量序列片段、錯誤序列、連接后引物序列以及無法與數據庫中序列信息比對一致的序列后,得到最終的有效序列。本次測序所得到的有效序列均可以達到后續微生物多樣性分析的要求。稀釋性曲線是通過對序列進行隨機抽樣,以抽到的序列數與它們所代表的物種數來構建的曲線。根據曲線走勢可以比較測序樣本的物種豐富度,同時說明測序數據的合理性以及測序量能否涉及所有菌群。
18 個樣品經過測序后得到的下機序列總數為589 542 個,其中有效序列總數達420 154 個,每個樣品的有效序列百分比均達60%以上。說明本次測序所得到的有效序列可以達到后續微生物多樣性分析的要求。
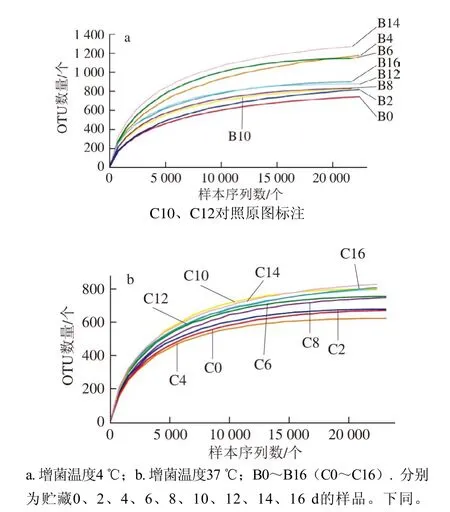
圖 1 不同增菌溫度下各樣品的稀釋性曲線Fig. 1 Rarefaction curves for each sample under different enrichment temperatures
利用R語言工具繪制曲線圖,由圖1可知,樣品在4 ℃增菌溫度下得到的OTU數量普遍比37 ℃多,說明在4 ℃增菌溫度下微生物的多樣性比37 ℃時更豐富。隨著測序量的不斷增加,每個樣品的稀釋性曲線都有趨向平緩的趨勢,說明本研究的檢測取樣合理,測序量基本反映出樣品中絕大部分的物種信息,本次實驗數據具有很大的實際價值。
2.2 不同增菌溫度下冰鮮鴿肉細菌群落的Alpha多樣性分析
根據97%相似性水平下的OTU信息,采用Alpha多樣性指數對樣品微生物物種的豐富度和多樣性進行評估。
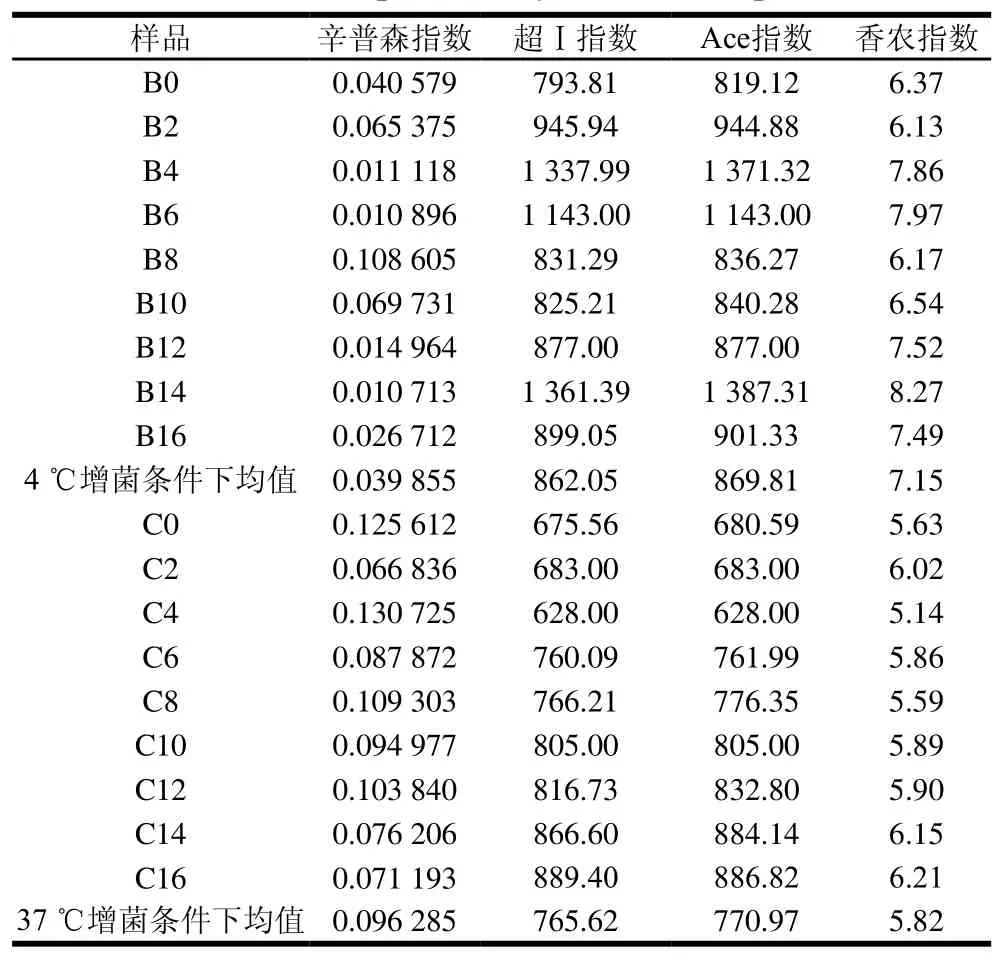
表 1 樣品的Alpha多樣性指數Table 1 Alpha diversity indexs for samples
由表1可知,樣品的辛普森指數、超Ⅰ指數、Ace指數及香農指數在4 ℃增菌溫度下的均值分別為0.039 855、862.05、869.81及7.15,在37 ℃增菌溫度下的均值分別為0.096 285、765.62、770.97及5.82,除辛普森指數外,4 ℃增菌溫度下的數值普遍高于37 ℃,表明在4 ℃增菌溫度下獲得的微生物群落多樣性與豐富度比37 ℃高。其中超Ⅰ指數和Ace指數最大的樣品為B14,其次為B4,最小的為樣品C4,說明4 ℃增菌溫度下貯藏14 d的冰鮮鴿肉物種豐富度達到最高。這可能是由于冰鮮鴿肉此時已腐敗,腐敗菌含量迅速增加,占主要優勢;而貯藏4 d時的物種豐富度也比較高,可能是由于此時冰鮮鴿肉中營養成分含量較高,細菌迅速生長繁殖,甚至達到細菌的對數生長期。香農指數最大的樣品為B14,最小的為C4。辛普森指數最小的樣品為B14,最大的為C4。上述結果表明,微生物群落多樣性最高的為4 ℃條件下貯藏14 d的冰鮮鴿肉樣品,最低的為37 ℃條件下藏貯4 d的冰鮮鴿肉樣品。
2.3 不同增菌溫度下細菌群落結構分析
采用RDP-classifier對各樣品中的物種數目進行物種注釋可以鑒定出冰鮮鴿肉貯藏過程中的細菌分屬于10 門、19 綱、28 目、39 科、53 屬。本研究就門、科分類學水平做具體分析。
2.3.1 基于門分類水平上的分析
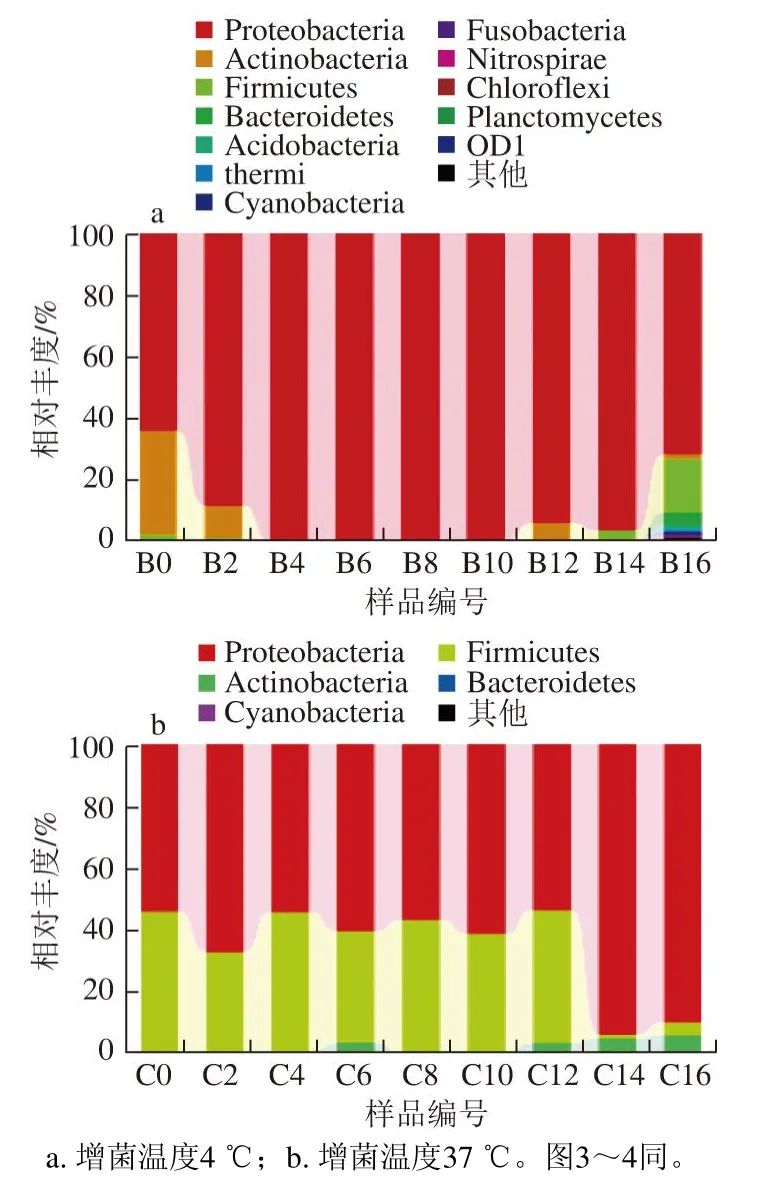
圖 2 基于門分類水平上的樣品菌群分布圖Fig. 2 Bacterial community distribution at the phylum level
由圖2可知,不同貯藏時間的冰鮮鴿肉微生物菌群在4 ℃和37 ℃增菌溫度下的菌相主要由變形菌門(Proteobacteria)、厚壁菌門(Firmicutes)和放線菌門(Actinobacteria)三大菌門組成。根據樣品在門水平上的菌群組成結果可以看出,4 ℃增菌溫度下變形菌門的總豐度為90.6%,放線菌門為5.8%,厚壁菌門為2.6%,37 ℃增菌溫度下變形菌門的豐度為66.1%,厚壁菌門為32.1%,放線菌門為1.8%。這說明不同貯藏時間的冰鮮鴿肉在不同增菌溫度下的微生物菌群組成均以變形菌門為優勢菌門,且4 ℃增菌溫度下尤為明顯。4 ℃增菌溫度下變形菌門的生長趨勢為由低到高,貯藏10 d時達到峰值,并在16 d時降至71.9%;放線菌門總體呈下降趨勢。而在37 ℃增菌溫度下,變形菌門的生長趨勢為由低到高,在14 d時達到峰值,厚壁菌門與變形菌門的生長呈相反的趨勢。除了這些主要菌門外,還存在一些豐度較低的擬桿菌門(Bacteroidetes)、酸桿菌門(Acidobacteria)、藍藻細菌門(Cyanobacteria)、梭桿菌門(Fusobacteria)、硝化螺旋菌門(Nitrospirae)、綠彎菌門(Chloroflexi)、浮霉菌門(Planctomycetes)以及其他未能分類的菌門,有的所占百分比不足0.01%,圖中顯示不明顯。這也說明冰鮮鴿肉在貯藏過程中的微生物多樣性豐富、豐度比較集中的特點。
2.3.2 基于科分類水平上的分析
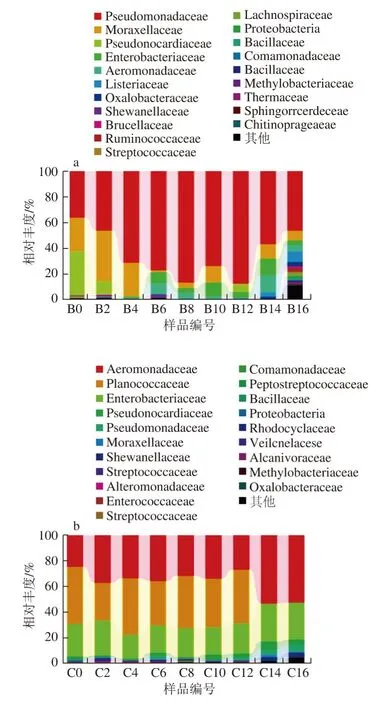
圖 3 基于科分類水平上的樣品菌群分布圖Fig. 3 Bacterial community distribution at the family level
由圖3可知,不同貯藏時間的冰鮮鴿肉在4 ℃和37 ℃增菌溫度下總共分離得到39 個不同的科。在門水平上占主要優勢的變形菌門(Proteobacteria)在科的分類水平上主要包括假單胞菌科(Pseudomonadaceae)、莫拉氏菌科(Moraxellaceae)、腸桿菌科(Enterobacteriaceae)、氣單胞菌科(Aeromonadaceae)、草酸桿菌科(Oxalobacteraceae)、希萬氏菌科(Shewanellaceae)、叢毛單胞菌科(Comamonadaceae)、甲基桿菌科(Methylobacteriaceae)、交替單胞菌科(Alteromonadaceae)、布魯氏菌科(Brucellaceae)、紅環菌科(Rhodocyclaceae)、食堿菌科(Alcanivoraceae)、柄桿菌科(Caulobacteraceae)及慢生根瘤菌科(Bradyrhizobiaceae)等。厚壁菌門(Firmicutes)在科水平上鑒定到的主要包括李斯特氏菌科(Listeriaceae)、疣微菌科(Ruminococcaceae)、鏈球菌科(Streptococcaceae)、毛螺菌科(Lachnospiraceae)、芽孢桿菌科(Bacillaceae)、消化鏈球菌科(Peptostreptococcaceae)、動球菌科(Planococcaceae)、腸球菌科(Enterococcaceae)及葡萄乳球菌科(Staphylococcaceae)等。放線菌門(Actinobacteria)在科水平上鑒定到的主要有假諾卡氏科(Pseudonocardiaceae)、微球菌科(Micrococcaceae)、微桿菌科(Microbacteriaceae)、棒狀桿菌科(Corynebacteriaceae)、紅椿菌科(Coriobacteriaceae)、段桿菌科(Brevibacteriaceae)及雙歧桿菌科(Brevibacteriaceae)等。
在4 ℃和37 ℃增菌溫度下,微生物菌群種類和豐度大小存在較大的差異性。在4 ℃增菌條件下,冰鮮鴿肉中的微生物菌群主要以假單胞菌科(Pseudomonadaceae)、莫拉氏菌科(Moraxellaceae)、假諾卡氏科(Pseudonocardiaceae)、腸桿菌科(Enterobacteriaceae)和氣單胞菌科(Aeromonadaceae)為主,其中假單胞菌科占比達64.6%,在4 ℃貯藏過程中呈先增長后下降的趨勢,貯藏16 d時達到峰值87.3%。假單胞菌科是冰鮮鴿肉貯藏過程中的主要優勢菌群,與目前大部分研究結果一致[15-16,29]。其次是莫拉氏菌科和假諾卡氏科,分別占14.5%和5.6%。在37 ℃增菌條件下,冰鮮鴿肉中的微生物菌群主要以氣單胞菌科(Aeromonadaceae)、動球菌科(Planococcaceae)、腸桿菌科(Enterobacteriaceae)、假單胞菌科(Pseudomonadaceae)和假諾卡氏科(Pseudonocardiaceae)為主,其中氣單胞菌科、動球菌科和腸桿菌科占比分別為36.7%、30.5%和24.4%。還有一部分菌群未能鑒定到具體菌科,有待進一步研究。
莫拉氏菌科(Moraxellaceae)和假單胞菌科(Pseudomonadaceae)在4 ℃增菌條件下的菌群豐度明顯高于37 ℃。莫拉氏菌科和假單胞菌科是典型嗜冷菌,是冰鮮鴿肉冷藏貯藏期間的優勢腐敗菌,這與原林等[30]的研究結果一致,說明莫拉氏菌科和假單胞菌科是冷凍肉制品、冷凍水產品等的優勢腐敗微生物。氣單胞菌科、動球菌科和腸桿菌科在37 ℃增菌條件下的菌群豐度明顯高于4 ℃,且37 ℃增菌條件下氣單胞菌科、假單胞菌科和假諾卡氏科所占的比例總體上呈增加趨勢,在貯藏16 d時達到最高,說明這幾類菌科是影響冰鮮鴿肉腐敗變質的優勢菌群。
綜合以上分析發現,低溫貯藏冰鮮鴿肉的主要菌群不僅包括嗜冷菌,還包括一些嗜溫菌。雖然樣本的貯藏條件為0~4 ℃,但是樣本中仍然存在一些嗜溫菌,生長溫度在30~37 ℃,如動桿菌科、腸桿菌科、氣單胞菌科等。因此冰鮮鴿肉在貯藏過程中一定要控制好溫度,溫度波動范圍太大會直接導致嗜溫菌等腐敗菌大量繁殖,促使冰鮮鴿肉腐敗。結合4 ℃低溫和37 ℃中溫2 種增菌溫度分析,可以較全面地反映冰鮮鴿肉在冷藏貯藏過程中污染的細菌群落結構的演替規律。
2.4 不同增菌溫度下冰鮮鴿肉細菌群落的PCA
依照不同貯藏時間將冰鮮鴿肉樣品分為9 組,每組由同一貯藏時間的4 ℃和37 ℃ 2 種增菌溫度的樣品組成。PCA圖中,橫軸表示第1主成分(PC1),縱軸表示第2主成分(PC2),圖中樣品之間位置越接近,表示樣品群落結構組成越相似。

圖 4 樣品細菌群落的PCAFig. 4 PCA of bacterial communities
由圖4可知,在4 ℃增菌條件下,PC1與PC2的貢獻率分別為68.48%和16.76%,在37 ℃增菌條件下,PC1與PC2的貢獻率分別為95.51%和2.78%,說明2 個主成分是解釋樣品微生物群落結構組成差異的主要因子。在4 ℃與37 ℃貯藏條件下,不同貯藏時間的各個樣品能明顯地分離,表明各個樣品間的微生物群落結構組成具有一定差異。在PC1方向上,4 ℃和37 ℃增菌條件下的樣品能夠顯著地分布在2 個區域,說明增菌溫度在PC1水平上對樣品菌群結構變化的影響顯著;在PC2方向上,不同貯藏時間的樣品區分明顯,說明貯藏時間在PC2水平上對樣品菌群結構變化的影響顯著。同一增菌溫度下,貯藏前期(0~4 d)的樣品與貯藏中期(6~10 d)、貯藏后期(10~16 d)的樣品之間分布距離較大,這主要是由于冰鮮鴿肉在貯藏前期仍處于鮮肉與次鮮肉狀態,而在貯藏中期開始加速腐敗,直到貯藏后期完全腐敗,進而導致冰鮮鴿肉群落結構組成發生較大變化。然而,2 種增菌溫度下,處于貯藏前期、中期、后期的各個樣品間距離較大,說明同一時期的樣品群落結構組成發生變化。
3 結 論
對冰鮮鴿肉在4 ℃和37 ℃ 2 種增菌溫度下的細菌群落多樣性進行研究,結果表明:在門水平上,4 ℃增菌溫度下變形菌門的豐度為90.6%,放線菌門為5.8%,厚壁菌門為2.6%;37 ℃增菌溫度下變形菌門的豐度為66.1%,厚壁菌門為32.1%,放線菌門為1.8%。其中,變形菌門從貯藏初期到貯藏后期的生長趨勢由低至高。在科水平上,在4 ℃增菌條件下,冰鮮鴿肉中的微生物菌群主要以假單胞菌科、莫拉氏菌科、假諾卡氏科、腸桿菌科和氣單胞菌科為主,其中假單胞菌科占比達64.6%,是冰鮮鴿肉貯藏過程中的主要優勢菌群,生長趨勢也是隨著時間的延長而增長。在37 ℃增菌條件下,冰鮮鴿肉中的微生物菌群主要以氣單胞菌科、動球菌科、腸桿菌科、假單胞菌科和假諾卡氏科為主,其中氣單胞菌科、動球菌科和腸桿菌科的占比分別為36.7%、30.5%和24.4%。貯藏前期,假單胞菌科、莫拉氏菌科為主要優勢菌科,但到后期莫拉氏菌科占比逐漸減少,而假諾卡氏科的占比逐漸增加。以上研究結果能夠幫助對冰鮮鴿肉低溫貯藏中的微生物菌群變化有較為全面的認識,要對冰鮮鴿肉進行全面質量管理的深入研究,基于門的水平,可以以變形菌門、厚壁菌門、擬桿菌門等優勢菌門為重點,基于科的水平,可以以假單胞菌科、莫拉氏菌科、假諾卡氏科、腸桿菌科和氣單胞菌科為重點。本研究結果對冰鮮鴿肉貯藏過程中微生物生長的抑制和延長冰鮮鴿肉貨架期方面的深入研究具有意義。
