甲狀旁腺素相關肽蛋白核定位序列及C末端缺失對小鼠下頜磨牙發育的影響??
2018-10-31 06:16:18顏成東蔣尚飛
重慶醫學 2018年29期
關鍵詞:小鼠
劉 洪,顏成東,蔣尚飛
(江蘇醫藥職業學院醫學技術學院,江蘇鹽城 224005)
甲狀旁腺素相關肽(parathyroid hormone related peptide,PTHrP)的結構和作用均和甲狀旁腺素(PTH)相類似[1]。PTHrP和PTH有相同的受體,并且PTHrP通過自分泌/旁分泌方式發揮多種生理功能[2]。PTHrP的N-末端(1~36)和PTH的功能基本相同;而PTHrP核定位序列(87~106)和C-末端共同通過細胞內分泌的方式達到促進細胞增殖的功能[3]。為了研究PTHrP核定位序列及C末端的功能,利用基因工程在PTHrP的第84位氨基酸序列后敲入一個終止密碼TGA,制備了不表達PTHrP核定位序列及C-末端的小鼠模型,稱為PTHrP Knock-in(PTHrP KI)小鼠。這種小鼠的表型有生存時間變短,以及骨骼、腦組織和皮膚發育不良等表現[4]。為了研究PTHrP核定位序列及C末端缺失對小鼠下頜磨牙發育是否存在影響,本研究利用出生后2周的PTHrP KI小鼠和野生型小鼠的下頜磨牙進行比較,研究PTHrP核定位序列及C末端缺失對小鼠下頜磨牙發育和礦化的影響。
1 材料與方法
1.1實驗動物 使用雄性PTHrP KI雜合子小鼠和雌性PTHrP KI雜合子小鼠交配(小鼠購自江蘇省實驗動物中心),獲得同窩的野生型(wild type,WT)小鼠和PTHrP KI純合子(PTHrP KI)仔鼠。仔鼠由母鼠喂養,母鼠使用普通飼料進行喂養。待小鼠生長至2周后取材,WT小鼠(WT組)和PTHrP KI小鼠(PTHrP KI組)分別取10只進行后續的影像學、組織學和分子生物學實驗。本次實驗經江蘇醫藥職業學院倫理委員會審核通過。
1.2 方法
1.2.1動物基因型鑒定 小鼠出生后剪取鼠尾,使用蛋白酶K進行消化,提取DNA后利用PCR進行目標片段擴增反應,產物使用BstEⅡ內切酶進行酶切反應后行瓊脂糖凝膠電泳,WT小鼠條帶為424 bp,PTHrP KI小鼠條帶為258 bp和166 bp[5]。
1.2.2取材 WT小鼠和PTHrP KI小鼠生長至2周后,頸椎脫臼處死,解剖分離左右兩側下頜骨,使用免疫電鏡固定液固定后行CT掃描,使用乙二胺四乙酸(EDTA)脫鈣后石蠟包埋,切片。
1.2.3影像學檢查 使用SkyScan 1072 scanner CT掃描儀對WT小鼠和PTHrP KI小鼠下頜骨進行掃描,通過CT檢查觀察,沿著下頜第一磨牙近中根管方向進行三維重建。
1.2.4蘇木精-伊紅(HE)染色 石蠟切片分別使用100%、75%和50%乙醇進行脫蠟,蘇木精染色8 min,伊紅染色2 min,分別使用50%、75%、100%乙醇脫水,二甲苯透明,中性樹膠封片。在光學顯微鏡下觀察放大100倍HE染色切片,測量兩種小鼠下頜第一磨牙長度,放大400倍HE染色切片測量兩種小鼠的下頜第一磨牙前期牙本質占總牙本質的比例。
1.2.5免疫組織化學檢查 石蠟切片使用牛睪丸透明質酸酶消化,Ⅰ型膠原(Collagen-Ⅰ)一抗為免抗鼠多克隆抗體(美國Santa Cruiz公司生產),二抗為生物素標記的羊抗兔二抗(美國Santa Cruiz公司生產),使用生物素-親和素-橋聯堿性磷酸酶酶標法(ABC-AP,美國Vector公司生產)孵育,堿性磷酸酶染色液染色后使用甲基綠復染。牙本質涎蛋白(dentin sialoprotein,DSP)、Ki-67和 p27一抗均為兔抗多克隆抗體(美國Santa Cruiz公司生產),二抗為辣根過氧化物酶標記的羊抗兔二抗(美國Santa Cruiz公司生產),鏈霉菌抗生物素蛋白-過氧化物酶連結法(Elite-AP,美國Vector公司生產)孵育后使用二氨基聯苯胺(DAB)染色液染色,DSP使用蘇木精復染。
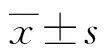
2 結 果
2.1影像學檢查 Micro-CT掃描三維重建結果顯示,PTHrP KI組小鼠的下頜第一磨牙牙根長度明顯小于WT組小鼠,見圖1。
2.2HE染色 WT組小鼠的下頜第一磨牙長度較PTHrP KI組小鼠長,差異有統計學意義(t=6.109,P<0.01);WT組小鼠的前期牙本質占總牙本質比例小于PTHrP KI組小鼠,差異有統計學意義(t=9.178,P<0.01),見圖2、表1。

A:WT組小鼠下頜骨(×100);B:PTHrP KI組小鼠下頜骨(×100);C:WT組小鼠下頜第一磨牙牙頸部(×400);D:PTHrP KI組小鼠下頜第一磨牙牙頸部(×400)圖2 兩組小鼠HE染色

表1 兩組小鼠下頜第一磨牙長度和前期牙本質占總牙本質比例比較

表2 兩組小鼠下頜第一磨牙Collagen-Ⅰ、DSP、Ki-67和p27陽性表達比較
2.3免疫組織化學染色 WT組小鼠Collagen-Ⅰ占下頜第一磨牙陽性比例、Ki-67陽性細胞比例和DSP占牙本質陽性比例均高于PTHrP KI組小鼠,p27陽性細胞比例低于PTHrP KI組小鼠,差異均有統計學意義(P<0.05),見圖3、表2。

A、B:Collagen-Ⅰ(×100);C、D:DSP(×400);E、F:p27(×400);G、H:Ki-67(×400)圖3 兩組小鼠磨牙中Collagen-I、DSP、Ki-67和p27陽性表達
3 討 論
在牙齒的發育、萌出和礦化過程中,多種鈣、磷調節激素如1,25(OH)2D3[6]、成纖維細胞生長因子-23(fibroblast growth factor-23,FGF-23)[7]的缺乏或者過表達都有可能干擾牙齒細胞外基質的分泌和礦化,最終導致牙齒發育異常。
本研究結果顯示,CT掃描后重建結果表明與WT小鼠比較,PTHrP KI小鼠下頜第一磨牙的長度縮短;通過對下頜第一磨牙進行HE染色、分析,結果發現PTHrP KI小鼠下頜第一磨牙的長度明顯縮短;進一步分析表明,PTHrP KI小鼠下頜第一磨牙前期牙本質的比例明顯高于WT小鼠。在牙本質的發育階段,成牙本質細胞分泌細胞外基質形成前期牙本質,前期牙本質在不斷礦化之后形成牙本質,如果前期牙本質的礦化過程受阻,則前期牙本質占總牙本質的比例就會明顯升高[6]。本研究HE染色結果表明,小鼠PTHrP核定位序列和C末端缺失可能導致了牙本質基質生成減少和礦化的異常。
應用免疫組織化學法進行分析,結果發現,PTHrP KI小鼠的Collagen-Ⅰ在磨牙區的陽性表達比例明顯小于WT小鼠。Collagen-Ⅰ是成牙本質細胞分泌的細胞外基質的主要成分,并且在牙本質礦化過程中能夠作為羥基磷灰石沉積的框架[8]。研究結果表明,小鼠PTHrP核定位序列和C-末端缺失導致了小鼠磨牙細胞外基質形成減少。PTHrP KI小鼠下頜第一磨牙牙本質中DSP的陽性面積明顯小于WT小鼠,DSP是由牙本質涎磷蛋白(dentin sialophosphoprotein,DSPP)基因轉錄后進行編輯合成,不僅能夠促進成牙本質細胞的分化,而且還能夠在牙本質的礦化過程中調節羥基磷灰石晶體的形成,而且能夠促進前期牙本質形成牙本質,因此,DSP也被認為是牙本質礦化的重要指標[9]。本研究結果表明,PTHrP核定位序列和C末端缺失導致了DSP合成減少,進而阻止了前期牙本質的礦化過程。免疫組織化學結果發現,PTHrP KI小鼠下頜第一磨牙牙髓和上皮根鞘處p27的陽性細胞比例明顯增加,而Ki-67的陽性細胞比例則有所降低。在有絲分裂期,Ki-67表達水平最高,因此可以把Ki-67作為細胞增殖狀態重要細胞因子[10-11]。p27是細胞周期蛋白依賴性激酶抑制因子(cyclin dependent kinase inhibitors,CDKI)家族成員,CDKI能夠和細胞周期蛋白-細胞周期蛋白依賴性激酶(CDK)復合物結合,抑制CDK的活性,從而阻斷細胞的增殖過程[12]。而p27是PTHrP的下游靶標,當PTHrP KI小鼠體內表達PTHrP蛋白中核定位序列和C-末端缺失,導致殘缺的PTHrP蛋白不能進入細胞核,喪失了對p27的調控抑制作用,從而導致了p27表達水平上調[13]。
PTHrP發揮促進細胞增殖、抑制細胞凋亡的作用是通過其蛋白的NLS和C-末端以細胞內分泌的方式進入細胞核后實現的,而PTHrP KI小鼠體內的PTHrP蛋白缺失了核定位序列和C-末端。體外研究表明,PTHrP缺失核定位序列和C-末端片段能調控細胞周期相關細胞因子的表達,從而使得細胞凋亡明顯增加[14]。研究發現,PTHrP在牙齒間充質細胞、牙乳頭、上皮根鞘等處均有表達[15],而PTHrP KI小鼠分泌的PTHrP蛋白因為缺失核定位序列和C-末端,失去了通過細胞內分泌方式進入細胞核來發揮促細胞增殖的作用,導致牙髓p27蛋白表達水平上調,Ki-67蛋白表達水平下降,導致牙髓間充質細胞的凋亡增加和向成牙本質細胞的分化減少,進而導致成牙本質細胞胞外基質分泌減少,并且進一步影響了牙本質的生成和礦化;此外,牙根的形成和上皮根鞘密切相關[16-17],同樣,PTHrP蛋白核定位序列和C-末端缺失還可能導致了小鼠磨牙上皮根鞘處的細胞凋亡增加,進而導致了PTHrP KI小鼠磨牙牙根發育障礙。
綜上所述,PTHrP核定位序列和C-末端缺失導致小鼠磨牙細胞增殖受阻,牙本質細胞外基質分泌減少,礦化障礙,最終導致了小鼠磨牙的發育異常。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
