水稻低溫敏感葉色突變體tcd32鑒定與遺傳分析
2018-11-09 08:34:04王妍斌林冬枝董彥君
上海師范大學學報·自然科學版 2018年5期
關鍵詞:水稻
王妍斌, 周 優, 林冬枝,2*, 董彥君,2*
(1.上海師范大學 生命與環境科學學院,上海 200234; 2.上海師范大學 遺傳研究所,上海 200234)
0 引 言
光合作用是影響水稻(OryzasativaL.)產量的關鍵因素之一,葉片是進行光合作用的主要器官,葉色深淺程度影響著光合作用的效率[1].葉色突變體是由葉綠素(CHL)合成和葉綠體發育相關基因沉默或失活引起的,直接或間接影響著葉綠素的合成和降解,改變葉綠素含量.大量研究表明,葉色突變體是研究植物光合作用機制、葉綠素生物合成途徑、葉綠體的結構功能、遺傳發育調控機理和農作物標記性狀等的理想材料[2-3].水稻葉色突變體有很多種,有一類突變體在不同溫度條件下呈現出不同的葉色表型,稱為水稻溫敏感葉色突變體,根據溫度對葉色表達的影響,將水稻溫敏感葉色突變體分為3種,即高溫表達型、低溫表達型和特殊溫敏感型[4-5].低溫表達型(水稻低溫敏感葉色)突變體是指水稻突變體的葉色表型在相對低的溫度條件下才會發生與正常葉色有差異的變化[6].目前報道有cde2[7],fan5[8],tsc1[9],tsl11[10],tcm11[6],tcm12[11],tws[12],lta1[13],1103(s)[14],ygl3[15],tsv3[16],tcd9[17],tcd5[18],tcd10[19],tcd11[20],OsGluRS[21].但其低溫誘導臨界溫度為20~28 ℃不等[22].高溫表達型(水稻高溫敏感葉色)突變體是指水稻突變體的葉色表型在相對高的溫度條件下會發生與正常葉色有差異的變化,與低溫條件下生長的水稻葉色表型無異(如tcm5[23]).鑒于溫度對溫敏感葉色水稻突變體葉綠體發育的影響,可以利用它們探究水稻的葉綠素合成、葉綠體發育,及光合溫度響應的分子機制.同時在水稻遺傳改良,高產育種中具有重要的潛在應用價值.
本研究通過60Coγ輻射水稻粳稻“嘉花1號”得到一個新的低溫敏感葉色突變體tcd32,對該突變體的葉色表型、光合色素含量、葉綠體顯微結構進行觀察和分析,并利用DNA分子標記對該突變基因進行了定位,為該基因的分子克隆及其作用機理的研究奠定基礎.
1 材料與方法
1.1 材 料
所用的低溫敏感葉色突變體tcd32是用粳稻“嘉花1號”干種子,經60Coγ射線輻照誘變所得,經多代在上海和海南自交繁育和選種,其農藝性狀已穩定.
1.2 方 法
1.2.1 苗期葉色表型觀察
將突變體tcd32與野生型(WT)“嘉花1號”的種子分別在30 ℃ 條件下催芽3 d后,將發芽的種子分別播種于20,25,30 ℃的光照培養箱(GXZ智能型,寧波江南儀器廠)中,每天光照12 h,光照照度為180 μmol·m-2·s-1.每隔2 d觀察1次幼苗的生長情況,觀察并記錄葉片的表型變化.
1.2.2 葉片光合色素含量的測定
苗期葉片中葉綠素a(CHLa),葉綠素b(CHLb)和類胡蘿卜素(CAR)含量的提取以及測定方法參考[24].分別選取在20,25,30 ℃條件下生長的突變體tcd32和野生型的三葉期幼苗的第三葉,剪取其葉片中間部分,稱取20 mg,剪碎后放入試管中,加入5 mL葉綠素提取液(體積比為:V乙醇∶V丙酮∶V水=5∶4∶1) 于試管中,密封后,輕輕搖勻,后置于室溫暗條件下處理18~24 h,直至葉片完全褪色.用METASH-UV5100分光光度計分別測定提取液對波長為470645663 nm的光的吸光度值,并分別計算出葉綠素a,葉綠素b和類胡蘿卜素的含量.實驗重復3次,計算其平均值.
1.2.3 葉片葉綠體顯微結構觀察
葉片取樣方法與上述1.2.2相同,然后將葉片沿葉脈方向剪成面積約1 mm2的正方形,放入2.0 mL的離心管中,之后立即放入2.5%(體積分數)的戊二醛固定液(體積比為:V25%戊二醛∶V0.2mol/L磷酸緩沖液∶
V無菌水=1∶4∶5),24 h后換新鮮的固定液,于4 ℃下保存,固定好后,進行清洗,再固定、洗滌、脫水與硬化,再經置換、浸漬、包埋、聚合、修塊、切片、染色等一系列處理過程后,用透射電鏡(Hitachi765型)進行觀察拍照.
1.2.4 遺傳分析與群體構建
利用培矮64S為母本,tcd32作為父本進行雜交,按常規種植加代收獲的F1代和F2代種子,作為遺傳研究和基因定位的群體.
1.2.5 水稻DNA提取
用十六烷基三甲基溴化銨(CTAB)法[25]提取親本突變體tcd32和培矮64S植株的DNA,F2代中用于定位的植株群體的DNA是用簡單、快速、便捷的Tris physiological saline(TPS)法[26]提取.
1.2.6 突變體基因定位
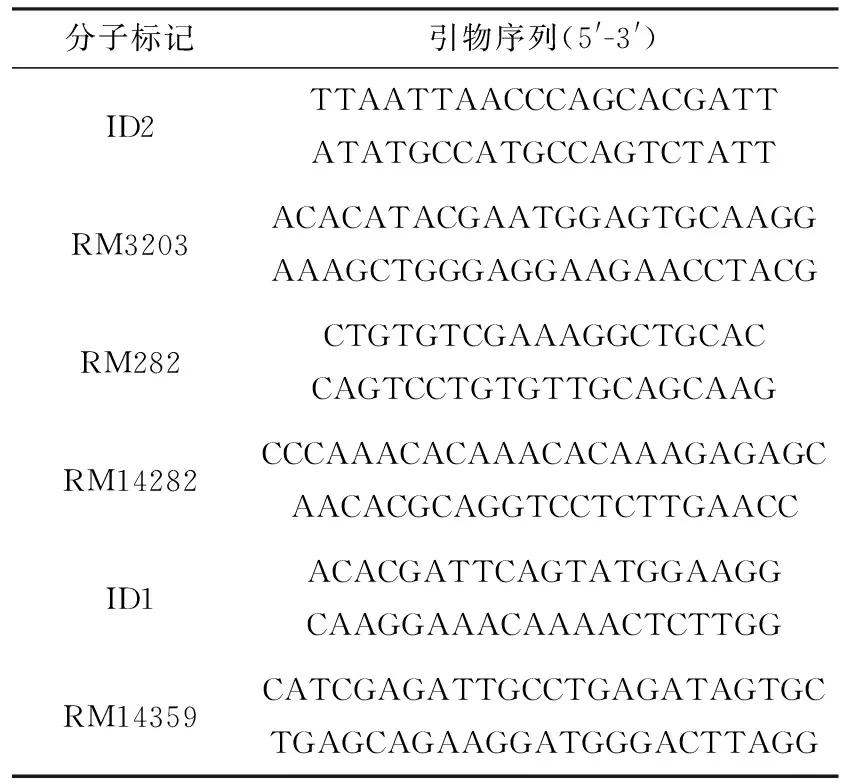
表1 用于基因定位的分子標記
利用本實驗室保存的水稻SSR和InDel引物,檢測突變體tcd32與培矮64S間的多態性.選取在染色體均勻分布且具有多態性的5~6對引物,對20 ℃低溫下生長的水稻中分離出的22個F2突變型株的DNA進行擴增,分析其帶型,確定突變基因所在染色體位置,并進一步擴大F2群體數進行基因定位.在目標區域內,根據美國國立生物技術信息中心(NCBI)公布的粳稻日本晴的全基因組序列和秈稻9311的全基因組序列間的差異InDel位點,利用Primer Premier5.0軟件設計引物(表1),進一步對突變基因進行定位.實驗DNA聚合酶鏈式反應(PCR)體系和PCR反應條件參照[26].PCR反應產物用2.0%~3.5%(質量分數)的瓊脂糖凝膠電泳檢測,經溴化乙錠染色后在紫外線凝膠成像儀上成像,并拍照記錄.用MapMaker/exp3.0軟件構建目的基因區域內的遺傳圖譜.
2 結果與分析
2.1 苗期突變體的表型

圖1 (a) 突變體tcd32和(b) 野生型“嘉花1號”在20,25,30 ℃條件下三葉期后生長過程
觀察突變體tcd32和野生型“嘉花1號”苗期植株葉色(圖1),結果顯示:tcd32突變體在20 ℃條件下,第一,二,三,四葉片均表現為白化表型,植株最終枯萎死亡;在25 ℃條件下,第一,二,三葉片表現為黃化表型,從第四葉期開始,葉片轉為黃綠色;但在30 ℃條件下,tcd32的植株表型與野生型沒有明顯差異.結果表明,tcd32的苗期葉色表型與生長溫度密切相關,具有低溫敏感葉色屬性.
2.2 突變體的光合色素含量變化
在3種不同溫度(20,25,30 ℃)條件下生長的突變體tcd32和野生型“嘉花1號”幼苗的三葉期葉片的光合色素測定結果(圖2)顯示:在20 ℃條件下,tcd32的光合色素含量(包括葉綠素a、葉綠素b以及類胡蘿卜素)與野生型相比顯著降低;在25 ℃條件下,tcd32的光合色素含量與野生型相比明顯降低;在30 ℃條件下,tcd32的光合色素含量與野生型的光合色素含量無明顯差異.此結果表明,苗期突變體光合色素含量與葉色表型相一致,具有溫敏屬性.
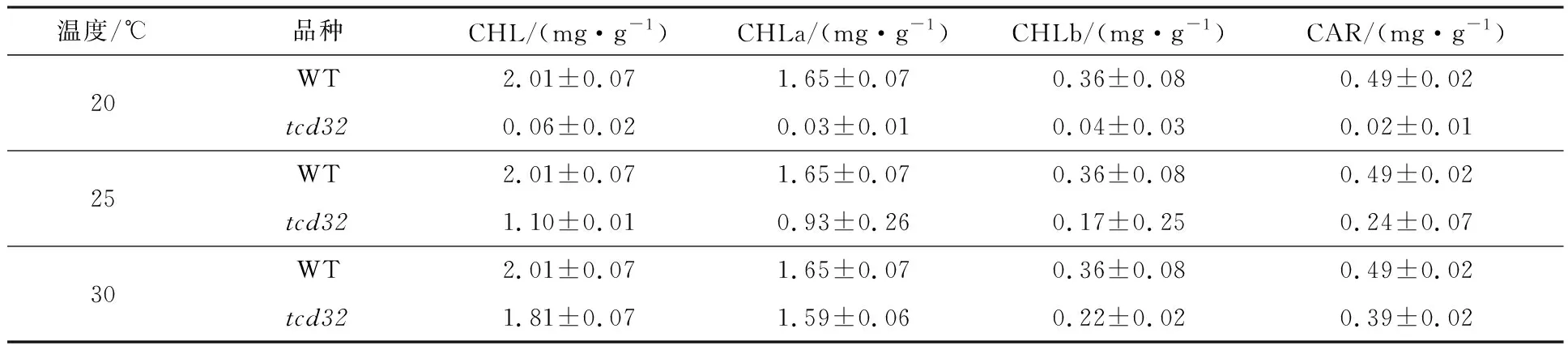
表2 突變體tcd32和野生型“嘉花1號”在不同溫度條件下的三葉期葉片光合色素含量
2.3 突變體葉綠體顯微結構
對在20 ℃和30 ℃下生長的突變體tcd32和野生型“嘉花1號”三葉期的第3葉中的葉綠體的顯微結構進行觀察(圖3),發現野生型“嘉花1號” 的葉綠體在高溫和低溫下都發育正常;而突變體tcd32在20 ℃下幾乎觀察不到一個完整的葉綠體結構,而在30 ℃下,突變體tcd32中的葉綠體又恢復正常發育,具有完整清晰的內囊體片層結構,與野生型相一致.這說明在低溫條件下該突變體tcd32的葉綠體發育分化受到嚴重影響,光合色素合成受阻,不能進行正常光合作用,進而產生突變表型.
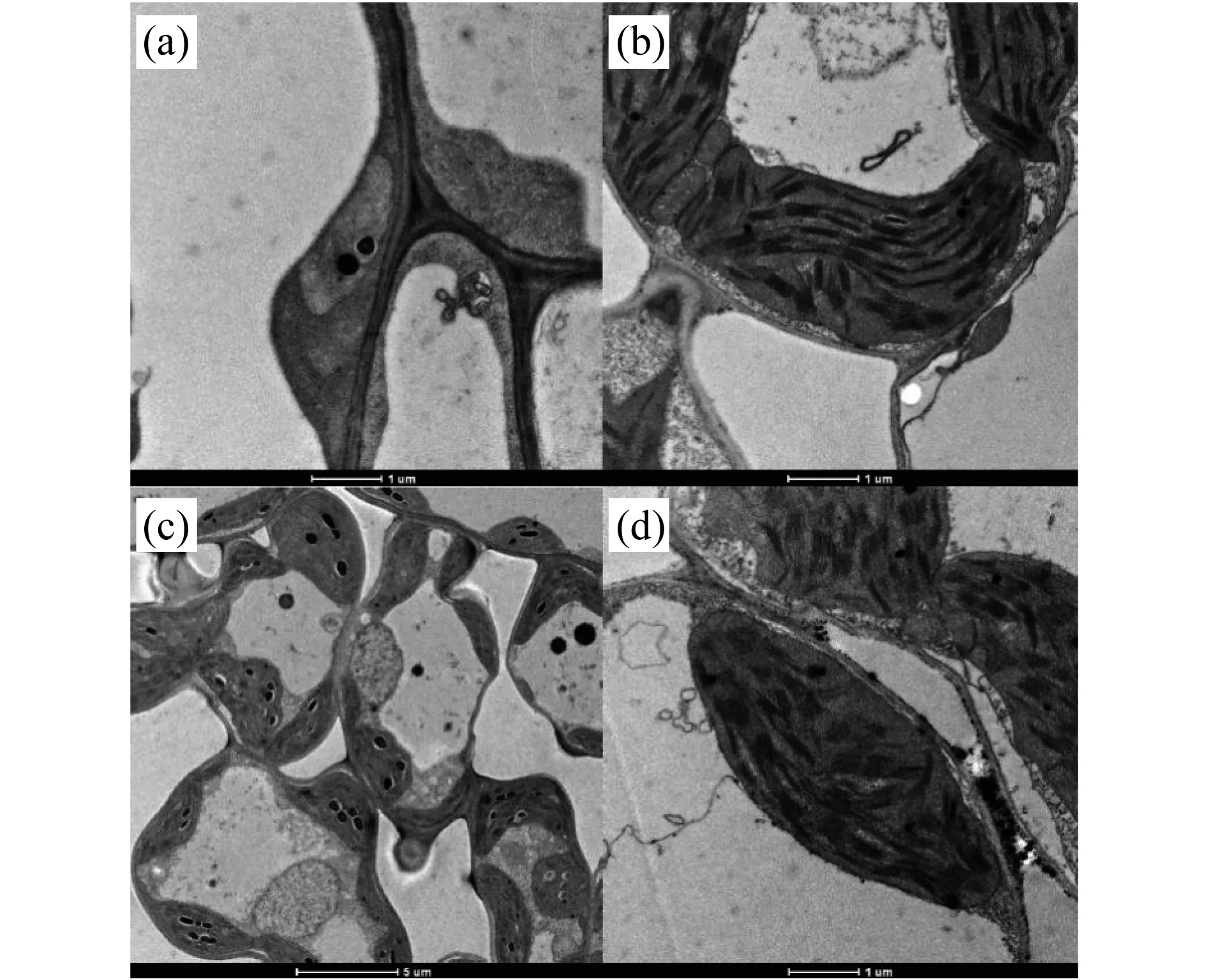
圖3 野生型“嘉花1號”和突變體tcd32植株在不同溫度條件下的葉綠體顯微結構.(a) 突變體tcd32,20 ℃;(b) 突變體tcd32,30 ℃;(c) WT,20 ℃;(d) WT,30 ℃
2.4 遺傳分析

2.5 基因定位
從F2代突變型白化苗中,選取22株進行連鎖分析,發現水稻第3號染色體上的分子標記RM14359有強烈的偏態擴增,如圖4(a)所示,表明tcd32位于第3染色體上,然后擴大F2群體至288株,將該基因tcd32定位在RM14359的上游,與其遺傳距離為9.2 cM,如圖4(b)所示,然后在RM14359的上游設計新的4個分子標記(ID1,ID2,RM14282,RM3203),將定位群體擴大至2887株,發現分子標記ID1和ID2均與突變基因tcd32表現共分離,而RM14282與tcd32基因的遺傳距離為1.3 cM,最終將tcd32基因鎖定在RM14282的上游的染色體頂端的537 kb內.
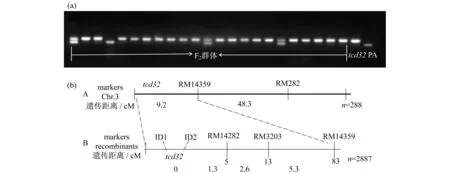
圖4 突變體在分子標記RM14359的(a) 電泳圖和(b) 遺傳定位圖
3 討 論
水稻是禾本科和單子葉植物的模式植物.隨著水稻基因組測序的完成,水稻的功能基因組學受到越來越多的關注[28].其中,葉色突變體是研究葉綠體發育與功能以及葉綠素代謝的理想材料.就不完全統計,缺乏葉綠素導致水稻葉色發生改變的突變體至少有297個,其中已定位到染色體上的突變基因有163個,在水稻12條染色體上均有分布.其中,第1,3,5,6,11染色體上居多,尤其第3染色體上多達38個,而在第12染色體上發現最少時只有4個[5,29-31].但在這些葉色突變體中,表現溫敏感的也有67個,其中低溫敏感型56個,高溫敏感型9個,特殊溫敏感型2個[5,29-30].分布在第1,3,4,9,11染色體上較多,在第3染色體上有13個,其中低溫敏感型有12個,高溫敏感型有1個(ST1[32]).而目前被克隆的低溫敏感型的葉色基因有:V1,V2,TCD5,TCD9,TCD10,TCD11.其中:V1基因編碼一個葉綠體蛋白NUS1[33];V2基因編碼一個調節葉綠體翻譯的新型鳥苷酸激酶(pt/mt GK)[34];TCD5基因編碼單加氧酶蛋白[18];TCD9基因編碼分子伴侶蛋白Cpn60α[17];TCD10編碼一個PPR蛋白[19];TCD11編碼核糖體S6蛋白[20],這些基因在低溫條件下對質體編碼基因的轉錄起調節作用,導致低溫條件下早期葉綠體發育異常.這表明水稻早期葉綠體發育過程中存在多種低溫響應的分子機制.另外,突變體tcd32表型與v1,v2,tcd5,tcd9,tcd10,tcd11類似,均表現為低溫敏感,即在低溫條件下葉片表現為黃化或白化,但在持續低溫條件下突變體tcd32死亡.在本研究中,將tcd32基因定位在第3染色體的頂端上的537 kb區間內,該區域包含89個候選基因,預測其中23個可能與葉綠體發育或葉綠素的合成相關,目前尚未被報道和克隆.今后,將在此基礎上繼續擴大定位群體,在染色體頂端設計更多新的分子標記.進一步對該突變體進行圖位克隆和功能分析,將有助于進一步了解水稻葉綠體發育分子作用機理.
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
