植物性狀研究的機遇與挑戰: 從器官到群落
2018-11-14 12:37:48何念鵬劉聰聰張佳慧于貴瑞
生態學報 2018年19期
何念鵬,劉聰聰,張佳慧,徐 麗,于貴瑞
1 中國科學院地理科學與資源研究所 生態系統網絡觀測與模擬重點實驗室, 北京 100101 2 中國科學院大學 資源與環境學院, 北京 100049
1 植物性狀及其研究進展
1.1 植物性狀及其主要研究方向
植物性狀(Plant trait)或植物功能性狀(Plant functional trait)是植物對外界環境長期響應與適應后所呈現出來的可量度的特征[1]。在實際研究過程中,人們更多關注與植物生長、繁殖、存活或與資源獲得、利用及其利用效率密切相關的性狀。植物是陸地生態系統初級生產力的主要參與者和貢獻者,因此大多數研究均圍繞植物性狀如何適應環境、優化資源利用或優化生產力而開展[2]。在指標上,更多選擇了能體現植物對環境適應并(或)影響植物生產力的特定屬性,如葉片形態特征(葉片大小、厚度、比葉面積和光合速率等)、根屬性(根大小、比根長和元素含量等)和種子大小與質量等[3-5]。
植物性狀研究可追溯到原始農業時期,人們通過對比發現與農作物產量和品質密切相關的性狀,再通過選育實現高產與優質。在生態學萌芽期開始,植物生態學家就開始了對自然群落的植物性狀的研究,重點關注植物如何通過特定性狀或多種性狀協同來實現群落結構穩定與生產力優化[4]。20世紀80年代后,植物性狀研究作為生物學、地學和環境科學交叉研究的紐帶,成為研究熱點。近年來,定量研究植物性狀(涉及碳收支、養分利用和水分經濟學)在不同尺度的空間變異規律、如何實現植物生產力優化是其中的重要研究內容,探討植被群落或植物多樣性對全球變化(氣候變化、氮沉降和酸沉降等)的響應與適應是重要的發展方向[5- 7]。本文對植物性狀的評述主要集中在陸地生態系統的自然群落性狀研究。
1.2 植物性狀研究的主要進展
1.2.1 種內和種間性狀變異及其空間格局
植物性狀種內與種間變異是當前最廣泛的研究內容。在控制實驗中,植物葉片形態(面積、厚度和比葉面積)、解剖結構(柵欄組織、海綿組織和表皮細胞)和氣孔都會隨著光照、溫度和CO2濃度變化而做出適應性的變化[8- 11]。在自然狀態下,羊草的葉片厚度、比葉面積、脯氨酸和K+/Na+隨著溫度和降水的變化而發生顯著的變化[12];擬南芥的個體大小和相對生長速率也呈現一定的緯度變異[13];光澤柳的比葉面積隨著土壤水分含量的增大而增大[14];3種櫟屬植物的葉片形態和化學組成與降水具有顯著的相關性[15]。與常綠植物相比,落葉植物的比葉面積更大[16];闊葉植物比針葉植物具有更高的氣孔密度[17];最大氣孔導度呈現出喬木>灌木>草本,落葉>常綠,闊葉>針葉[18]。Cornwell和Ackerly[14]發現植物性狀沿環境梯度呈現的規律主要來自于種間變異,而種內變異的影響較小。
已有研究圍繞植物葉片性狀(葉片大小、葉片厚度、比葉面積、氮含量(N)、磷含量(P)、光合速率和光響應飽和曲線)和根性狀(根分級特征、根大小、根長度、比根長、N、P、根瘤特征和菌根真菌特征),探討了植物性狀在不同生境的變異規律、特定物種(屬或科)根葉性狀沿環境梯度的變化規律[19-21]。Wang等[20]利用847種植物的實際調查數據,探討了葉面積大小、葉厚度、比葉面積和葉片干物質含量4個性狀沿3700 km的中國東部森林樣帶(NSTEC)的變化規律。Wang等[17]采用了壓痕法調查了NSTEC樣帶760種植物的氣孔性狀(氣孔密度和氣孔大小),首次探討了區域尺度葉片氣孔特征,并從物種-功能群-群落層次揭示了植物氣孔性狀沿緯度變異規律及其主要影響因素。Tian等[22]對NSTEC樣帶優勢喬木的葉片解剖結構性狀進行了調查分析(上表皮厚度、下表皮厚度、柵欄組織厚度和海綿組織厚度等),闡明了葉片解剖性狀的緯度格局與影響因素。上述研究基本形成了葉片常規形態性狀-氣孔性狀-解剖結構性狀的由表及里的指標體系。
近年來,圍繞葉片C、N、P含量的空間格局及其影響機制開展了較多研究。例如,Ordonez等[23]通過數據收集整合分析了全球尺度植物比葉面積、葉片N、P和N∶P的空間格局及其影響因素。He等[19]通過分析內蒙古、西藏和新疆3個地區213種草原植物C、N及其C∶N,發現3個地區間草原植物的C∶N是相對穩定的,氣候通過改變植物組成來影響草原植物葉N含量。Chen等[24]調查了中國14個森林386種木本植物葉片N、P含量及其N∶P比,并從氣候、土壤和植物生活型角度探討了其影響因素。Han等[25-26]通過收集文獻數據,探討了中國區域植物葉片N∶P化學計量特征以及葉片多元素的生物地理格局及其影響因素。
根性狀及其生物地理格局也是當前植物性狀研究的熱點之一,相關研究提高了人們對植物如何適應環境和養分利用策略的認識。Comas和Eissenstat[27]報道了25個共存物種的比根長特征。 Gordon和Jackson[28]針對植物細根元素含量(C、N和P)及C∶N∶P進行研究,揭示了植物根系在元素周轉中的作用。郭大立等圍繞中國森林植物根形態、根瘤和外生菌根等開展了系統的研究工作,較深入地揭示了植物對養分利用策略,并深入探討了中國從南到北優勢喬木細根形態的空間變異特征[29-32]。
在全球尺度上,種子質量隨著緯度的升高而降低,植物功能群和植被類型的變化是造成種子質量緯度變化的重要原因[33];Moles等[34]發現植物最大高度會隨著緯度升高而發生明顯的變化。
1.2.2 植物多種性狀協同與權衡
在自然環境中能長期生存并繁衍的植物,既需要具有適應環境變化的能力,還必須具備能獲得其生長和繁殖所必需光、水和養分的能力,是通過植物多種性狀協同與權衡來實現的。植物葉片和根系具有很多可測量的性狀,每個性狀都能不同程度的反映植物對環境的適應或者植物對資源的獲取能力。例如,比葉面積和葉片N含量等多種性狀都對植物光合速率具有較大的影響[12];同時,葉片氣孔密度和解剖結構等間接影響植物光合速率和水分利用效率[18,35-36]。類似地,細根是植物獲取養分的最重要器官;其中,根長度、比根長和根瘤特征與養分獲取能力密切相關,同時也與植物蒸騰能力等間接相關。植物不僅通過特定器官內部的多種性狀的協同(如比葉面積與N含量)[37];還通過葉-根之間16種元素的協同控制機制,來實現對環境的響應與適應[38]。Wang等[39]發現葉和根的形態特征趨異而元素含量趨同是植物葉-根協同的重要機制。雖然相關研究已經取得了長足進展,但由于植物性狀的多樣性及其所面臨環境的復雜性,還有許多問題仍待解決。
Wright等[16,40]利用全球葉片性狀數據庫提出了著名的葉經濟型譜(Leaf economic spectrum, LES);植物葉片性狀沿著一條連續變化的功能性狀組合譜有序排列, 其一端代表著比葉重小、含N量高、光合速率大、呼吸速率高、葉壽命短的“快速投資-收益型”策略;而另一端代表著壽命長、比葉重大、含N量低、光合速率和呼吸速率都偏低的“緩慢投資-收益型”策略。在此基礎上,Diaz等[6]利用4.6萬種成年植物高度、葉面積、葉N含量等6個指標,從物種水平驗證與發展了葉經濟型譜理論,并發現樹高-種子質量維度與葉經濟譜系維度相互獨立。LES的提出與發展,極大地提高了人們對植物通過性狀協同實現生產力優化機制的認識,尤其是為解釋碳氮元素在植物內固定、利用和流動提供了重要的理論依據。然而,關于LES的形成動力仍存在較大的爭議;一方面,Maire等[41]認為環境梯度引發植物資源權衡效應,植物通過自我調節來響應環境變化,因此LES主要受環境因子主導;另一方面,目前LES還多集中在資源獲取的維度,然而自然群落中影響植物光合作用的因素是多維的,未來還應考慮水分維度和溫度維度等[42]。
近期,根系經濟學譜系(Root economic spectrum, RES)和木材經濟學譜系(Wood economic spectrum, WES)也被提出。RES把根系的呼吸強度、比根長和N含量等歸為獲得性強的性狀(Acquisitive traits),而把根系干物質含量、根系直徑和木質素/N含量歸為更保守的性狀[43]。WES認為:較高的木質密度有利于抵擋害蟲和病原體的侵害,更加具有生物力學上的穩定性,從而提高了物種的壽命;較低的木質密度和較高的水分含量意味著更少的投資成本,因此物種有較高的樹高和直徑生長速度[44]。
1.2.3 植物性狀與功能的關系
自然群落中的植物性狀研究不應局限在特定器官、特定物種或特定科屬上,而應重點探討多種植物如何通過特定性狀或多種性狀協同,來實現群落結構穩定與功能優化機制。目前,植物性狀-功能研究大多是建立在器官水平,如葉片N含量與光合速率的關系、葉片比葉面積與光合速率關系、葉片葉綠素含量與光合速率的關系,這些個體或者物種水平得出的結論能否適用于到群落水平,還有待實測數據證實。近年來,如何將個體或者物種水平測定的性狀與群落結構和功能相結合,或從性狀角度解釋群落結構構建機制,是當前植物性狀研究的熱點與難點[45]。Kunstler等[46]通過分析全球14萬個樣地、300萬顆樹的最大生長速率、密度、高度和比葉面積數據,發現在區域和全球尺度上述性狀與競爭關系整體表現出較好的一致性,為預測森林物種間相互關系提供了美好的愿景;同時發現性狀的不相似性(dissimilarity)在局部群落對生產力影響方面的作用較小,但這種不相似性導致的權衡(Trade-off)確是影響物種間競爭和促進物種共存的重要基礎。Cadotte等[47]通過45個喬木物種15種性狀的分析,發現在物種水平葉片6種功能性狀與光合速率具有顯著的相關性。功能性狀的群落加權平均值一直被認為是獲得群落水平性狀的潛在的重要途徑,豐富度指數的加權方式最為常見,但這種加權方式忽略了自然群落中物種個體間的差異,基于物種生物量異速生長方程的群落生物量加權方式可很大程度克服上述不足。Wang等[20]通過結合生物量異速生長方程的群落生物量加權方式,探討了森林植物葉片大小、比葉面積等的緯度格局,發現氣候因素和土壤N含量具有重要影響。此外,氣孔性狀的群落生物量加權平均值與生態系統水分利用效率顯著正關性[18];葉片解剖結構的群落加權平均值與生態系統生產力和水分利用效率具有緊密的聯系[36]。
何念鵬等借助中國東部南北樣帶9個典型森林生態系統全面系統的調查數據(包括群落結構數據、樣地內1120多個物種葉-枝-干-根樣品的16種元素含量、葉片常規性狀-氣孔性狀-解剖結構性狀、葉片熱值-葉綠素-非結構性碳水化合物、1—5級細根形態性狀-元素含量等),借助群落結構數據+生物量異速生長方程+比葉面積數據,實現了部分植物性狀從器官-物種-功能群-群落的推導,并進一步建立了天然森林群落植物性狀與功能的聯系。一些研究結果揭示了天然森林群落中特定性狀與功能的定量關系,具有一定的理論和實踐意義[18,36]。然而,目前有關天然森林群落植物性狀與功能聯系的研究,仍集中在單一性狀,未來更應從多性狀協同角度探討大尺度下森林群落性狀與功能的關系。
1.2.4 植物性狀如何表征陸地植被對全球變化的響應與適應
結合陸地生態系統植被的結構和功能,定量分析植物性狀對全球變化(氣候變化、氮沉降和酸沉降等)的響應與適應是生態學近年來的研究熱點。為提高模型模擬精度,將部分重要的植物性狀納入模型正逐漸成為未來模型發展的重要方向[38,41]。然而,由于傳統的植物性狀研究大多在物種水平進行,缺乏群落尺度的數據,一定程度地制約著模型在大尺度上的發展和應用。
植物化學計量特征為科學家探討全球變化的效應提供了重要途徑。Elser等[48-49]通過整合分析不同湖泊和全球控制實驗(N或P添加),揭示了海洋和陸地生態系統中的磷限制性效應;并在此基礎上,定量評估未來N沉降增加情景下陸地生態系統凈初級生產力的變化。Yan等[50]通過整合分析全球淡水中植物N和P累積及其生態效應,發現在大氣沉降和人為富營養化情況下,淡水系統的P累積遠高于N累積,從而對淡水生態系統生產力形成巨大的負效應,需要加強治理力度使這些淡水系統盡快恢復到初始狀態。Zhu等[51]基于實測數據發現中國區域大氣沉降N∶P遠高于植被和土壤,并預測不平衡的N∶P輸入將進一步增強陸地生態系統磷元素的限制效應。此外,葉片N∶P比常用于確定不同地區的限制性元素,當N∶P < 14是為氮限制,當N∶P > 16時為磷限制。近年來,國內外學者從個體或群落尺度都開展了很多相關研究[25,52-53]。同時,隨著生態化學計量學的日益成熟,人們提出可用植物群落不同組分C∶N比值,結合N沉降通量來構建陸地生態系統固碳效應的理論框架[54];最近,Zhu等[55]基于中國區域氮沉降量以及NESCT樣帶9個典型森林生態系統不同植物組分C∶N比值,基于化學計量學基本理論評估了大氣氮沉降量對中國森林生態系統的固碳效應。
2 植物性狀研究面臨的機遇與挑戰
國內外學者已經在植物性狀研究領域開展了大量的研究工作,并取得了豐碩的成果。仔細分析上述4個主要進展的數據源,會發現在這些研究依然以優勢種為主體與重點,并未考慮天然森林群落結構與物種組成的復雜性。近年來,國內研究提出利用長期固定監測樣地開展系統的性狀調查工作(如中國生態系統研究網絡(CERN)、中國國家生態系統觀測研究網絡(CNERN)、中國森林生態系統定位觀測網絡(CTERN)、中國森林生物多樣性監測網絡(CForBio)),這此平臺為人們開展森林植物性狀、性狀-功能關系提供了非常理想的平臺,但近期其成果能尺度上推到森林群落的還很少[6,16]。總之,個體水平所獲得的性狀-功能的關系,在自然群落中是否適用還有待證實;同時,植物多種性狀協同(或權衡)是植物對環境響應與適應的重要途徑,但該思想在實際研究中很少被貫徹,未來可從如下幾個方面深入探索。
2.1 重視多種性狀協同或趨異規律的研究
以葉片為例,前期科研人員非常重視葉片常規性狀、元素含量、光合特性等研究[6,16,23]。但長期忽視葉片氣孔屬性(氣孔大小、氣孔密度和氣孔比面積)和葉片解剖屬性(上表皮厚度、下表皮厚度和柵欄組織厚度等)。未來的研究不僅要從各個性狀的種內、種間、區域變異分析,還更應該深入探討多種性狀的協同規律,從植物對環境的響應和適應角度分析。
2.2 以植物群落構造為基礎,開展系統性的植物性狀調查
在群落調查時,要盡可能覆蓋所有物種,并且每個物種盡可能涵蓋葉、枝、莖、根和種子等。目前,在包括比較權威的世界植物屬性數據庫(TRY數據庫),均是以收集公開發表數據為基礎而建立的,每個群落一般僅搜集一個或幾個物種的數據;由于未考慮群落結構,相關研究是將物種水平等同于群落水平[6,16,46]。此外,TRY數據庫的植物屬性數據,絕大多數集中在葉片,非常缺乏植物葉-枝-莖-根-種子的配套數據。
2.3 在群落尺度開展性狀-功能關系研究,突破性狀研究的難點
受測試技術與研究手段的限制,前期研究多是在個體水平探討性狀-功能的關系;在生態系統、區域或全球尺度上,一直存在性狀數據與功能數據空間不匹配的問題[45]。例如,植物性狀的測定大多在個體或者種群水平開展,而大尺度研究的功能更多在群落或生態系統水平進行,二者間存在空間尺度與量綱間的不匹配。因此,如何實現植物性狀從個體或者物種-功能群-群落-生態系統水平的尺度拓展,建立性狀-生態系統功能的定量聯系,是揭示生態系統對外界環境和資源變化響應與適應機制的基礎,也是改進和優化模型、提高其預測精度的重要條件。近期,國外學者正在嘗試建立植物性狀與生態系統功能之間的關系[18,36-37,56- 59]。總體來說,這方面的研究還非常少,且研究方法和主要結論仍需要進一步改進與證實。為了加快這一過程,我們認為需要發展新的概念體系、指標體系,真正實現植物性狀與宏觀生態學的聯系;通過與高新技術的結合,既拓展植物性狀研究的范疇,又使植物性狀研究能更好地服務于區域甚至全球生態環境問題的解決。
3 群落性狀或生態系統性狀的定義與內涵
當前,生態學研究正在快速向宏觀方向發展,關注的最基本尺度是群落或生態系統;另外,全球變化對陸地生態系統的影響評估也是以群落或生態系統結構與功能為重要對象。現階段大多數的植物性狀研究主要集中在植物器官或者個體水平,僅有少數研究將它們拓展到群落水平;在拓展過程中有3種方法:1)簡單算數平均法;2)相對生物量或多度加權平均法;3)生物量加權平均法。方法1和方法2所獲得的群落水平的植物性狀(如葉片氮含量g/kg或%),雖然可以有效地探討群落結構維持機制,由于無法實現對群落性狀在量綱上向單位土地面積轉換,使它很難與模型和遙感數據相匹配。而方法3可以獲得基于單位土地面積的群落性狀,在空間尺度匹配(或量綱匹配)的前提下實現個體水平測定的植物性狀數據與生態模型和遙感觀測相聯系,更好地探討區域尺度下自然生態系統結構和功能的關系及其對全球變化的響應與適應[36]。植物性狀研究的壁壘和宏觀生態學發展的現實需求,促使人們重新思考植物性狀研究的尺度問題;需要將植物性狀統一到群落尺度或單位土地面積尺度,才能與宏觀生態學研究的主要觀測手段緊密聯系起來(通量觀測、模型模擬和遙感觀測),推動相關學科發展。
群落性狀(Community trait)可定義為:在群落尺度能體現植物對環境適應性和生產力優化的性狀(其定義也適用于動物或土壤微生物等)。其核心內涵:1)群落性狀均被轉化為以單位土地面積為基數的性狀,如葉片大小、葉片生物量、葉片氣孔密度、葉片N和P含量等;2)任何群落性狀均是可測量或可推導,原則上均是采用群落生物量加權平均的方法進行推導的;3)群落性狀應能從不同層面反映植物對環境的適應或生產力的優化,即具有明確的生態學意義。群落性狀的提出和發展,一方面有利于將各種植物性狀在單位土地面積上標準化,便于深入探討不同性狀間的內部關系、協同或趨異規律,進而更好地從性狀角度揭示植物群落的構建與維持機制。另一方面,通過建立單位土地面積群落性狀,可解決長期以來植物性狀數據與宏觀生態學主要測試數據(遙感、通量觀測和模型等)空間尺度不匹配的問題,推動宏觀生態學的發展、更好地服務于區域生態環境問題的解決。從長遠看,只有充分利用各種高新技術的學科,才能獲得更快的發展;因此,群落性狀提出與發展將會成為相關領域新的生長點。為了更好地統籌自然生態系統中的植物、動物、微生物、土壤等要素,未來應進一步發展以單位面積為基礎的生態系統性狀(Ecosystem trait),生態系統性狀將在相對統一的空間尺度和量綱上獲得植物群落性狀、動物群落性狀、土壤微生物群落性狀、土壤屬性和氣候要素等,為深入研究不同尺度生物-非生物性狀間的關系奠定堅實基礎,并以群落性狀為核心連接宏觀生態學的主要研究途徑(遙感觀測、通量觀測和模型模擬),推動傳統性狀指標與宏觀生態學的應用相結合。
4 群落性狀指標體系初探
圖1給出了一個典型森林生態系統的性狀指標的基本框架圖,并展示了生態系統性狀、生態系統結構和生態系統功能的整體構架及其相互關系。在生態系統結構參數的協助下,可利用植物群落生物量加權法(非傳統的直接算術平均法和相對生物量加權平均法),將器官水平測定的植物性狀參數推導至群落水平,從而在同一尺度建立生態系統內植被-大氣間的功能參數(總初級生產力、凈初級生產力和蒸騰速率等)和土壤-大氣界面的功能參數的定量關系,構建群落性狀-功能間的橋梁。
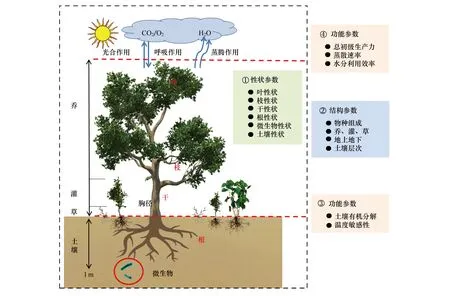
圖1 森林群落性狀與功能的基本框架圖Fig.1 Logical picture of community traits and functioning in natural forest圖中植物群落性狀參數均是通過群落結構參數,利用生物量加權法將器官水平測定的性狀推導到整個群落,并以單位土地面積為基礎參數
按圖1的框架,建立生態系統性狀-結構-功能的基本指標體系、測試途徑和推導方法(表1)。以葉片性狀為例,從葉片形態性狀(葉形狀、葉大小和比葉面積等)、氣孔形狀(氣孔大小、氣孔密度、氣孔比面積等)、解剖結構性狀(上表皮厚度、柵欄組織厚度、下表皮厚度等)和多元素含量性狀(C、N、P含量等)著手。在系統測試的基礎上,再利用比葉面積+異速生長方程+群落結構數據推導獲得各種基于單位土地面積的群落性狀。目前建議的指標體系,會在未來發展過程中逐步增加與完善,例如,此次沒有包括葉片光合速率,理由如下:1)葉片光合速率受溫度和水分影響非常大,在短時間內都存在較大的差異;2)葉片光合速率在群落或生態系統水平常用總初級生產力、凈初級生產力等指標來替代,并可通過通量觀測系統測定。在實際操作過程中,個體或器官水平測定的光合速率難以推導到自然群落水平;雖然植物最大光合速率是一個相對穩定的指標,但自然界中由于物種冗余、相互競爭等因素,植物最大光合速率與群落凈初級生產力的關系非常復雜,不同地點和不同生態系統類型間存在顯著變異。
植物性狀要實現從器官-群落的推導,理論上需要滿足兩個基本條件:1)調查應覆蓋植物群落出現的所有物種;2)詳細的植被群落結構調查數據。在實際操作過程中,針對每個群落需要開展詳細的調查,這是通過文獻搜集數據難以實現尺度上推的重要原因。網絡化長期定監測樣地(CERN、CNERN、CTERN和CForBio)為開展系統性植物性狀測定提供了較為理想的平臺,伴隨著“群落性狀”概念的提出和推導方法的逐步成熟,將有助于建立傳統性狀研究與宏觀生態學的橋梁。何念鵬等利用熱帶-寒溫帶9個典型森林生態系統詳細的調查數據,并嘗試性地利用生物量加權法將葉片氣孔從器官推導至群落水平,并發現群落水平的氣孔性狀呈現出明顯的緯度格局,且與生態系統總初級生產力和水分利用效率顯著相關[18,35-36],提供了天然森林群落性狀與功能聯系的直接證據。Zhang等[53]利用該方法獲得了中國東部8個森林葉-枝-干-根和土壤的C、N、P含量的加權平均值,并發現其結果與前期收集數據(數據直接平均法)所獲得結果差異顯著。過去幾年基于數據直接平均法或生物量相對加權法的研究較多,但真正基于群落生物量加權法的研究還僅在少數幾個站點實現;尤其是在如何實現由點至面拓展、天然群落性狀-功能定量評估、群落性狀與快速發展的宏觀生態學觀測技術的聯系等方面,還有待深入研究。
5 群落性狀的應用前景與展望
(基于單位土地面積的)群落性狀作為一個新概念,其指標體系和應用將會是一個逐漸深入和拓展的過程。隨著調查數據的逐漸豐富,它在如下幾方面具有較好的應用前景:1)群落性狀將有利于更好地探討自然群落的性狀與功能間的定量。在傳統研究中,大多數研究都是在物種水平測試植物性狀并探討性狀與功能的關系,并將這種物種水平的定量關系直接應用于自然群落(物種水平=群落水平);在復雜的自然群落中,這種簡單替換的定量關系的可信度還有待實測數據的進一步證實。2)群落性狀將有利于揭示全球變化對生態系統結構和功能的影響。由于群落性狀能與通量觀測、近地面遙感和衛星遙感的觀測數據在單位土地面積上匹配,使性狀研究可更好地結合或服務于相關的新技術,即推動新技術的發展,又為性狀研究提供新的生長點。當前,全球變化的生態效應多是在生態系統水平進行評估的;在利用N∶P比變化來探討養分限制性或未來氮磷沉降不對稱的生態效應時,如果能用群落水平的N∶P比(群落內不同物種長期適應與權衡的整體表現形式),其評估結果應該可以更準確。3)為生態模型提供更多和更準確的關鍵參數,提高模型模擬精度。以C∶N比為例,由于群落水平數據難以獲得,大多數生態模型均使用個別物種數據或少數物種算數平均會值來替代,如能提供更精確的群落水平C∶N,應該可以提高相關模型的預測精度。4)群落性狀的提出能促使更多植被性狀納入生態模型,為開發新一代模型奠定堅實基礎[60]。目前,許多學者已經意識到納入更多性狀參數是提高預測精度的有效措施,隨著群落性狀數據的逐步增多,未來一定會有更多性狀參數被納入模型以提高模型精度。
如果未來能進一步發展生態系統性狀,以單位土地面積為基礎,建立植物群落性狀、動物群落性狀、土壤微生物群落性狀、土壤屬性和氣候特征的聯系,將有利于從新視角探討生態系統內部生物和非生物間的有機聯系。在實際調查過程中,許多學者在特定地點或多個地點,都開展了系統的群落結構與性狀調查,但由于傳統概念與個人理解的差異,這類數據或群落性狀很少被公布并積累下來,這是性狀研究的巨大損失。鑒于此,本文探討了群落性狀的定義及其潛在應用,希望能引起國內同行對性狀研究概念與方法的思考,推動性狀研究從物種水平向群落水平的跨越,甚至拓展到區域或全球水平。同時,希望新的概念體系能逐步將傳統性狀與高速發展的新技術(定量遙感、通量觀測、生態模型)相結合,更好地服務于區域生態環境問題的解決,拓展性狀研究的新生長點。
猜你喜歡
英語世界(2023年10期)2023-11-17 09:18:18
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
