基于群體Parrondo博弈的根分枝建模方法
2018-11-22 09:37:54李松陽高繼勛劉曉東余文奇
計算機應用 2018年10期
李松陽,高繼勛,王 淼,劉曉東,余文奇
(河南工程學院 計算機學院, 鄭州 451191)(*通信作者電子郵箱lsy_cqu@cqu.edu.cn)
0 引言
根系是植物連接土壤與地上部分之間實現物質交換的重要橋梁,是植物吸收水分、養分資源和響應土壤環境變化的重要器官[1]。植物根系結構十分復雜,且埋藏在不透明介質中,難以觀測,對根系進行定量化研究十分困難,因此建立根系結構模型,采用計算機模擬重現根發育過程是用于根系研究非常有用的工具。由于根系的分枝過程對根系生長發育有重要影響,其可塑性導致復雜根系的產生[2],因此在根系結構模型中,如何對根系的分枝過程建模是一個重要的問題。目前國外主流的根系三維結構模型包括RootMap、RootTyp、SimRoot、SpacSys和RootBox等[3]。在RootMap模型中,通過定義分枝密度,頂端不分枝持續時間或分枝延遲時間等參數實現對根系分枝過程的模擬[4-5]。在RootTyp模型中,通過定義分枝間隔控制根原基的產生,經過根原基的發育延遲時間后根原基發展為分枝[6]。在SimRoot模型中,通過定義經驗的分枝頻率以及生長角度實現對根系分枝過程的仿真[7]。在隨后的OpenSimRoot模型中,依然通過參數分枝頻率對根系分枝進行建模[8]。在SpacSys模型中,通過參數分枝位置實現對根系分枝過程的仿真,其中分枝位置由頂端不分枝長度、基端不分枝長度以及分枝間隔所決定[9]。在RootBox以及隨后的CRootBox模型中,通過預定義分枝距離、分枝數目、頂端不分枝長度,以及基端不分枝長度等從參數決定根系分枝產生的位置[10-11]。在國內,張吳平等[12]定義了一個表征根個體周期性生長發育的基本單元(根元)。當根元的生長年齡大于1時,根元上的側根原基可以發育形成分枝。金明現等[13]依據參數距頂端長度實現側根位置的模擬。通過以上描述可以看出,現有的國內外主流根系結構模型中認為根分枝過程具有向頂的趨勢,是一個向頂序列,并且沒有考慮根原基對根分枝的作用。雖然根原基是順序出現的,是一個向頂序列。但根原基發展成為根分枝是一個受到環境以及根原基本身影響的可調制過程,并不完全是順序的。研究表明,雖然根原基的產生具有向頂序列的特性,但根原基發展為側根的過程并不是一個嚴格的向頂序列[14]。為了彌補上述方法的不足,充分考慮環境以及根原基對根分枝形成的影響,本文提出一種基于群體Parrondo博弈的根分枝仿真方法,用于異質根系生長環境下根分枝可塑性建模。該方法通過構建根原基為個體的根原基群體,采用Parrondo博弈實現根原基群體間的交互,依據群體交互結果,實現根原基個體中生長素更新,從而對根分枝過程實現建模,為根系建模與仿真研究提供模型支撐和分析手段。
1 模型描述
Parrondo博弈認為:對于給定的兩個博弈,如果每個博弈輸的概率都大于贏的概率,那么當這兩個博弈隨機或周期進行時就可能產生一種獲勝的結果。同時生物個體具有兩重屬性——種群屬性和自然屬性,種群(社會)屬性決定了同種個體之間存在競爭合作關系,自然屬性則揭示生存環境對個體存在的影響[15]。由于根系結構具有環境適應的特性,因此對于根系來說,各器官組織間的協調尤其重要。本文采用群體Parrondo博弈模型來模擬根系分枝過程,按照如下步驟進行:
步驟1 構建以根原基為個體的根原基群體。依據根自相似結構,劃分根為頂端不分枝區域n_la,基端不分枝區域n_lb,以及分枝區域lb;隨著根的伸長,當根長度大于頂端不分枝區n_la與基端不分枝區域n_lb長度之和時,依據根原基的序列產生方式及其發展窗口dw大小,對分枝區域lb進行順序分割,則分割區域i即為第i個根原基rpi,分割區域i+1為根原基rpi+1,其中根原基rpi與根原基rpi+1為鄰居根原基,最后,建立包含多個根原基個體rpi的根原基群體RS(如圖1所示)。

圖1 根原基群體示意圖Fig. 1 Schematic diagram of root primordial swarm
步驟2 獲取某個根原基個體rpi的鄰居根原基集合。遍歷根原基群體RS,對于其中的第i個根原基rpi,獲取其鄰居根原基集合rp_Neg={rpi-1,rpi+1}。如果rp_Neg不為空,遍歷rp_Neg,對于其中的第j個根原基rpj,計算rpi與rpj的交互結果;如果rp_Neg為空,則rpi不與其他根原基交互。
步驟3 計算根原基個體rpi感知環境因素影響因子Ei及環境變化強度g。以根原基rpi為旋轉軸,以r為旋轉半徑構建圓柱形區域,獲取當前時刻t時該區域內的環境因素總量Ei_upt(t)。設定根原基rpi最小資源需求量Ei_min,最優資源需求量下限Ei_opt1,最優資源需求量上限Ei_opt2,最大資源需求量Ei_max,計算當前時刻t時環境因素影響因子Ei(t)(如式(1)所示)以及環境變化強度g(如式(2)所示)。
Ei(t)=
(1)
(2)
其中:h為顯著性變化下限;Δt為生長間隔時間。
步驟4 采用Parrondo博弈模型計算rpi與rpj交互的輸贏。rpi與rpj隨機選擇以概率p′=p-ε*Ei進行A博弈或者以概率1-p′進行B博弈,ε為斜率,其取值為正小數。如果進行A博弈,依據當前時刻t時rpi與rpj生長素含量Ci(t)和Cj(t),初始生長素含量Ci_0和Cj_0以及生長時間Ti和Tj,計算當前時刻t時rpi與rpj的生長素收益Wi(t)(如式(3)所示)和Wj(t)(如式(4)所示);計算當前時刻t時rpi相對于rpj贏的概率pij(如式(5)所示)。
Wi(t)=Ci(t)-Ci_0
(3)
Wj(t)=Cj(t)-Cj_0
(4)
(5)
如果進行B博弈,當在時刻t時rpi生長素含量Ci(t)能夠被M整除時,計算環境影響的rpi贏的概率為p2′=p2-ε*Ei, 當在時刻t時rpi生長素含量Ci(t)不能夠被M整除時,計算環境影響的rpi贏的概率為p3′=p3-ε*Ei。其中p2、p3分別為兩種情況rpi贏的概率最大值。
步驟5 計算在時刻t+Δt時根原基rpi生長素含量和生長時間Ti(如式(6)所示)。如果進行A博弈,當rpi贏時,rpj支付a單位的生長素給根原基個體rpi;當rpi輸時,rpi支付a單位的生長素給根原基個體rpj。如果進行B博弈,如果rpi贏時,rpi增加a單位的生長素;如果rpi輸時,rpi減少a單位的生長素。設定根原基個體rpi博弈贏時符號sig=1,博弈輸時符號sig=-1,及其在生長間隔Δt中固定收益b,則計算在時刻t+Δt的rpi生長素含量Ci(t+Δt)(如式(7)所示)。
Ti+Δt=Ti+Δt
(6)
Ci(t+Δt)=Ci(t)+sig*a+b;b>a
(7)
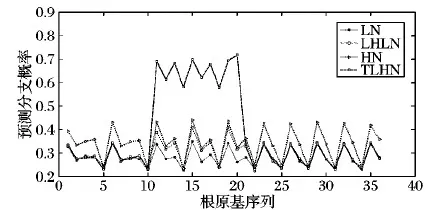
步驟6 根分枝預測。遍歷根原基群體RS,對于其中的第i個根原基個體rpi,獲取在時刻t時rpi生長素含量Ci(t)。設定rpi生長素需求最大值Ci_max及其發育時間Tlim。如果Ti 設定根系最大生長周期為20 d,根系依據生長函數(式(8))進行生長[11]。 λ(t)=k(1-exp(-(ρ/k)t)) (8) 其中:k為根最大長度;ρ為初始生長速度。當根長大于n_la與n_lb之和后,依據根原基發展窗口dw大小,按照向頂的方向順序對lb區域進行分割,從而構建根原基個體,形成根原基群體,進而通過根原基群體Parrondo博弈實現根分枝仿真。設dw=0.4,n_la=1.57,n_lb=0.07,k=26.9,ρ=2。根原基環境感知半徑r=0.01,最小資源需求量Ei_min=30,最優資源需求量下限Ei_opt1=130,最優資源需求量上限Ei_opt2=230,最大資源需求量Ei_max=330,h=0.05,初始生長素含量Ci_0=1和Cj_0=1,概率p=0.5,概率p2=0.15,概率p3=0.75,斜率ε=0.01,M=3,生長素支付a=1,固定收益b=2,生長時間間隔Δt=1,生長素需求最大值Ci_max=9,根原基發育時間Tlim=3。 為了驗證模型的有效性,以氮養分為例,設置了4種不同的根系生長環境:1)根系處于均勻低氮氮脅迫生長環境中(氮含量設置為90 μmol/L),記為符號LN。2)根系處于均勻高氮氮充足的生長環境中(氮含量設置為50 μmol/L),記為符號HN。3)首先根系生長處于均勻低氮氮脅迫生長環境中,經過一段時間后,部分根系處于均勻高氮氮充足的生長環境中(在仿真周期前15天,根原基分布于氮含量設置為50 μmol/L環境中;在仿真周期后5天,第10到第20個根原基分布于氮含量設置為90 μmol/L環境中),記為符號TLHN。4)根系處于異質的低氮氮脅迫,高氮氮充足和低氮氮脅迫的生長環境中(序號1~10個根原基分布于氮含量設置為50 μmol/L環境中,序號10~20個根原基分布于氮含量設置為90 μmol/L環境中,剩余根原基分布于氮含量設置為50 μmol/L環境中),記為符號LHLN。 圖2為在進行10 000次仿真后,群體Parrondo博弈下環境環境影響的根分枝預測圖示。在HN條件下比在LN條件下,根原基發展為根分枝的概率有明顯的提升,該仿真結果與Mounier等[16]實驗結果一致。在TLHN條件下,可以看出序號為10~20之間的根原基發展為根分枝的概率有明顯的提高。該仿真結果與Zhang等[17]觀察到的實驗結果一致。在LHLN條件下,可以看出在氮含量豐富的區域,根原基發展為根分枝的概率更高,意味著該區域會有較多的分枝。該仿真結果與Giehl等[18]的實驗結果一致。通過仿真實驗結果表明,本文方法得出的根分枝預測能夠實現對根原基轉化為根分枝過程的建模。 為了便于對比分析本文方法與RootMap等方法在根分枝建模上差異,假設在國外主流的RootMap、RootTyp、SimRoot等方法中分枝間隔大小等于根原基發展窗口dw。圖3給出了按照根分枝是向頂序列的思想在RootMap等方法中產生根分枝的示意圖。可以看出RootMap等方法在建模根分枝時沒有考慮環境資源分布不均以及環境資源分布隨時間變化對根分枝的影響,僅僅按照分枝間隔等參數,以概率1產生分枝。在該過程中資源分布對根分枝的影響,是通過資源分布影響根伸長,從而影響根分枝數目。但本文方法資源分布不僅僅通過影響根伸長,影響根分枝數目,還通過影響根原基發育為根分枝的過程,實現對根分枝的建模。 圖2 根分枝預測圖示Fig. 2 Root branching prediction diagram 圖3 RootMap等方法實現根分枝示意圖Fig. 3 Root branching by RootMap etc 通過對基于群體Parrondo博弈的根分枝模型的分析,發現Parrondo博弈的結果對根原基群體的交互結果有著重要的影響。因此Parrondo博弈中A博弈選擇概率p,B博弈中當生長素含量能夠被M整除時博弈概率p2以及當生長素含量不能夠被M整除時博弈的概率p3對根分枝有重要影響的參數(p2,p3的選擇要滿足負博弈的條件[19])。在LH條件下,通過設置不同的p、p2和p3值,分析參數調整對根分枝的影響(見圖4)。從圖4(a)可以看出,p值的選擇對根原基發展為根分枝過程有重要的影響。當p值較大時(p=0.55),說明A博弈被選擇概率較高。因此根原基間競爭加劇,從而造成根原基發展為分枝的概率有激烈的差異;當p值較小時(p=0.4),說明A博弈被選擇概率較低,從而造成根原基間競爭強度降低,合作意愿加強,根原基更傾向于從外部環境中獲取生長素,從而造成根原基發展為分枝的概率的差異相對較小。 在B博弈中,概率p2、p3的選擇對于根原基發展為根分枝的過程也有重要的影響(圖4(b)、(c))。直觀上來看p2、p3增加,表示根原基從外部獲取的生長素概率隨之增大,因此根原基發展成為根分枝的概率也會增加。從圖4(b)、(c)中可以看出,當p2=0.2或者p3=0.8時,根原基的分枝概率有很大的提高。當p2=0.1或者p3=0.7時,根原基的分枝概率相對較低。但p2表示B博弈中當生長素含量能夠被M整除時博弈概率,因此p2對少部分根原基發展為根分枝的過程影響劇烈。從圖4(b)可以看出根原基發展為根分枝的概率差異較大。概率p3表示B博弈中當生長素含量不能夠被M整除時博弈概率,因此p3對大部分根原基發展為根分枝的過程會有影響。從圖4(c)可以看出,當p3值增大后,大部分根原基發展為根分枝的概率會提高。 圖4 在LN條件下不同 p、 p2、 p3值對根分枝的影響Fig. 4 Effect on root branching by p, p2, p3 under LN condition 根系結構的適應性變化對植物的生存具有重要的要義。根分枝是復雜根系結構形成的重要過程。但根原基發展為根分枝的過程展示適應環境變化的可塑特性,并不是一個序列過程。但現有根系生長模型中,通常把根分枝的產生視為一個向頂的序列。本文基于根原基受控順序產生的特性,進一步把根原基視為智能個體,通過根原基個體對環境的感知以及根原基群間的競爭或者合作,通過根原基間的群體Parrondo博弈實現根原基的群體決策,從而實現根分枝可調制的預測,避免了傳統根系建模與仿真研究中根分枝產生方式的不足,從而能夠為植物建模與仿真整體研究提供模型支撐和分析手段,能推動植物建模與仿真應用于精確農業。2 仿真分析
2.1 仿真參數設置
2.2 仿真實驗設置
2.3 仿真結果分析
2.4 Parrondo博弈概率對根分枝影響的分析

3 結語
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
小讀者(2021年2期)2021-03-29 05:03:48
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
光學精密工程(2016年6期)2016-11-07 09:07:19
紅領巾·萌芽(2016年1期)2016-09-10 07:22:44
