漿細胞樣樹突狀細胞及TLR7 TLR9在BP皮損中的表達及臨床意義
2018-11-22 02:10:18朱寧韓睿程浩
浙江臨床醫學 2018年9期
朱寧 韓睿 程浩?
鮑溫樣丘疹病(BP)臨床表現為青壯年外生殖器部位多發色素性扁平丘疹,組織病理呈低度惡性原位鱗癌表現,本病的發生、發展及復發與人乳頭瘤病毒(HPV)尤其是與HPV16、18等高危型感染有關。漿細胞樣樹突狀細胞(pDC)作為機體重要的免疫效應細胞,通過其選擇性高表達的Toll樣受體7和9(TLR7和TLR9)識別病毒核酸,引起pDC活化、IFN-α和其他炎癥性細胞因子的分泌并激活其他類型免疫細胞,從而起到抗病毒感染的作用[1]。研究顯示[2]BP患者皮損處存在免疫抑制狀態,特別是局部細胞免疫受抑制,造成一系列免疫細胞和相關細胞因子的數量和功能異常有關,但具體機制尚未完全闡明。本文探討pDC和TLR7、TLR9在BP病毒免疫方面發揮的作用,現報道如下。
1 臨床資料
1.1 一般資料 選取2008年7月至2013年12月浙江大學醫學院附屬邵逸夫醫院36例BP患者的皮損蠟塊,均符合BP臨床診斷并經組織病理確診。其中男26例,女10例;年齡21~64歲,平均年齡(33.06±10.38)歲。病程1個月~3年,平均(7.33±8.00)個月。男性皮損部位包括龜頭、冠狀溝、包皮、陰莖、陰囊;女性皮損部位包括會陰、肛周、陰道口、大小陰唇等。所有病例均排除自身免疫性疾病、病毒性肝炎、結核及糖尿病等可能影響機體免疫力的疾病。本研究經醫院和學校倫理委員會批準,患者簽署知情同意書。
1.2 主要試劑 鼠抗人CD123單克隆抗體、兔抗人TLR7多克隆抗體、兔抗人TLR9多克隆抗體(均購自美國Santa Cruz公司),即用型UltraSensitivTM S-P免疫組化試劑盒、二抗(生物素標記的羊抗鼠、羊抗兔)、DAB酶底物顯色劑(均購自北京中杉金橋生物技術有限公司)。
1.3 方法 (1)CD123、TLR7、TLR9的免疫組化染色:分別取36例BP患者局部皮損和皮損旁正常皮膚組織,標本經10%中性緩沖福爾馬林固定,石蠟包埋,連續切片厚度為4 μm。采用免疫組化鏈霉素抗生物素蛋白-過氧化酶法(SP法),按照試劑盒說明書操作,常規脫蠟,水化,抗原修復,依次滴加過氧化物酶阻斷劑,封閉血清,分別用鼠抗人CD123單克隆抗體、兔抗人TLR7多克隆抗體、兔抗人TLR9多克隆抗體進行免疫組化染色,以PBS代替一抗作為陰性對照,一抗工作液稀釋濃度為1:100,生物素標記的二抗,鏈霉素抗生物素-過氧化物酶溶液,DAB顯色,蘇木素復染并封片,光鏡下觀察并判定結果。(2)免疫組化染色結果判定:①CD123+pDC的表達及計數[3]:10×40倍光鏡下觀察,表皮、真皮內以細胞漿和/或細胞膜著色呈褐色并有樹突狀突起的細胞為CD1a+LC、CD123+pDC,每張切片隨機選取表皮、真皮乳頭層為主的5個視野計數CD1a+LC、CD123+pDC細胞數和細胞總數,CD1a+LC、CD123+pDC密度=5個視野CD1a+LC、CD123+pDC細胞總數/5個視野總細胞數。②TLR7、TLR9的表達及半定量計分[4]:10×40倍光鏡下觀察,以細胞漿和/或細胞膜有棕褐色到淡黃色顆粒著色者為TLR7、TLR9陽性細胞,表達強度采用基于染色強度和陽性細胞百分率的半定量計分法:染色強度計分(A):3分:棕褐色;2分:棕黃色;1分:淡黃色;0分:未著色;陽性細胞百分率計分(B):3分:>66%的表皮細胞著色;2分:33%~66%的表皮細胞著色;1分:<33%的表皮細胞著色,TLR7、TLR9的表達強度為:A×B。每張切片隨機選取5個視野進行陽性細胞觀察并予以半定量計分,取平均值作為10×40倍光鏡下每個視野內TLR7、TLR9表達半定量計分的平均結果。
1.4 統計學方法 采用SPSS 20.0統計軟件。計量資料以()表示,兩組樣本均數比較采用t檢驗,相關性分析采用Pearson相關法,以P<0.05為差異有統計學意義。
2 結果
2.1 BP皮損組織中CD123+pDC的分布與密度 CD123分子主要表達于真皮乳頭層的單核細胞,為棕褐色胞漿著色的樹突狀細胞,結果見圖1。BP皮損真皮乳頭層中CD123+pDC密度(0.351±0.107)比皮損旁正常皮膚真皮乳頭CD123+pDC密度(0.232±0.111)明顯增多(P<0.01),且體積較大,呈多角形,見表1。
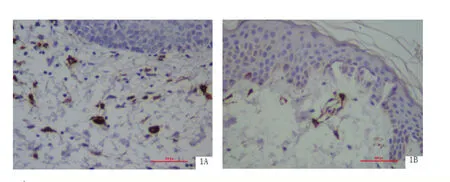
圖1 CD123+pDC在BP皮損組織和皮損旁正常皮膚組織中的分布與密度(SP×400)
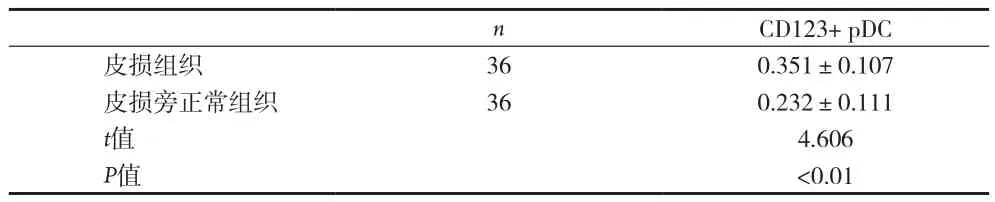
表1 BP皮損組織和皮損旁正常皮膚組織中CD123+pDC的表達比較
2.2 BP皮損組織中TLR7的表達 TLR7在真皮上部表達較強,主要位于胞漿。TLR7在BP皮損真皮內表達強度明顯高于皮損旁正常皮膚組織真皮內[(5.083±2.285)vs.(3.611±1.498),P<0.01],見圖 2、表2。
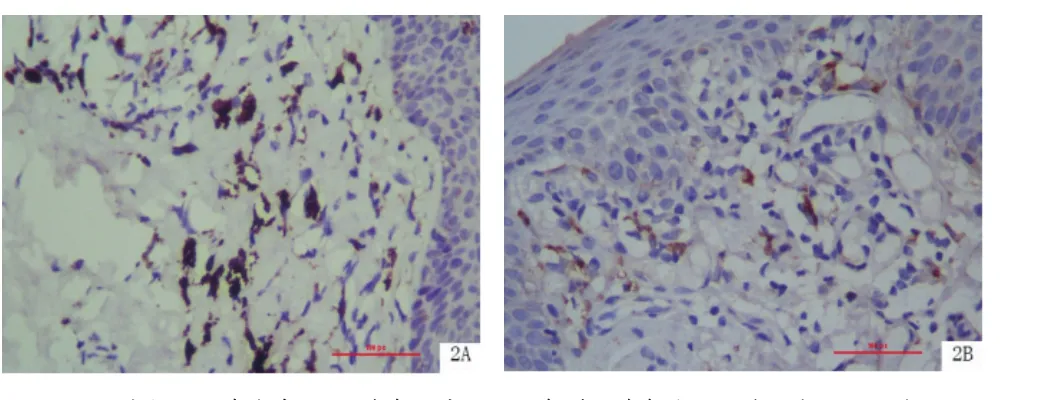
圖2 TLR7在BP皮損組織和皮損旁正常皮膚組織中的表達(SP×400)
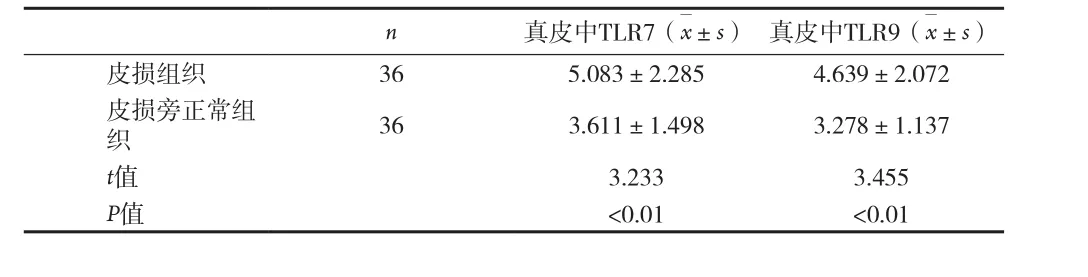
表2 BP皮損和皮損旁正常皮膚的真皮組織、表皮組織中TLR7、TLR9的表達比較
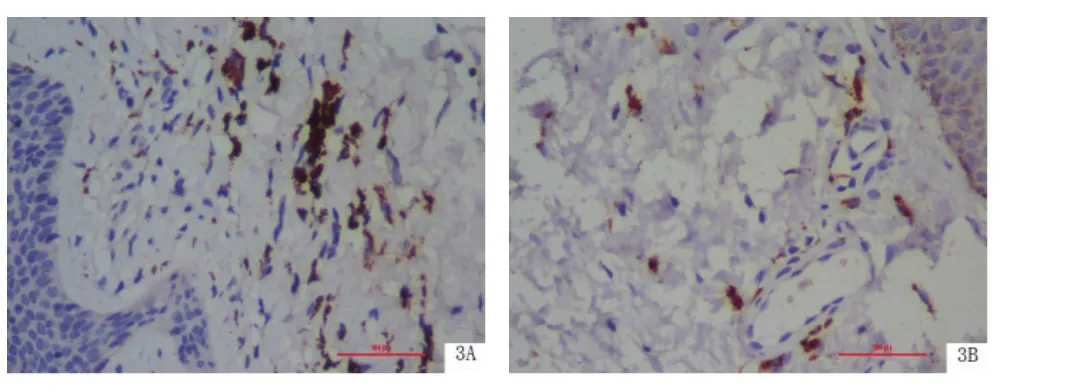
圖3 TLR9在BP皮損組織和皮損旁正常皮膚組織中的表達(SP×400)
2.3 BP皮損組織中TLR9的表達 TLR9主要定位于細胞的胞漿,以真皮上部為主(4.639±2.072),結果見圖3。TLR9在BP皮損真皮內表達強度明顯高于皮損旁正常皮膚組織真皮內[(3.278±1.137),P<0.01],見表2。
2.4 BP皮損CD123+pDC細胞密度與TLR7、TLR9表達的相關性分析 BP皮損組織中CD123+pDC細胞密度較高,皮損真皮中TLR7、TLR9高表達,皮損組織中CD123+pDC密度與皮損真皮中TLR7表達水平及與TLR9表達水平間均呈直線正相關關系(前者r=0.617,P=0.000;后者 r=0.505,P=0.002),見圖 4A、B。在BP皮損旁正常皮膚組織中CD123+pDC細胞密度較低,皮損旁正常皮膚組織真皮中TLR7、TLR9低表達,皮損旁正常皮膚組織中CD123+pDC密度與皮損旁正常皮膚組織真皮中TLR7及與TLR9之間均呈直線正相關關系(前者r=0.617,P=0.000;后者r=0.522,P=0.001)。
3 討論
pDC是機體免疫系統在抗病毒感染過程中主要產生IFN-α的細胞,能夠對多種病毒感染作出應答。人類較多病毒性疾病均存在不同程度的pDC數量和功能改變,但不同的研究結果存在一定的爭議。
目前國內外關于HPV感染相關的疾病中pDC的免疫作用和地位研究報道甚少,特別在與高危型HPV16、18感染關系密切的BP的發病機制中pDC的作用地位還未見報道。Zhu Xiaoxia等[3]報道尖銳濕疣患者皮損中CD2AP+pDC、CD123+pDC的密度和陽性率均比對照組邊緣正常組織高。Van Seters等[5]報道HPV(+)的外陰上皮內瘤變(VIN)患者冷凍切片中pDC的數量增加。Bontkes HJ等[6]報道83%的HPV感染相關宮頸癌患者的冷凍切片中有pDC存在,pDC表達的HPV16病毒衣殼VLP受體—CD49f,與HPV16 VLP共培養分泌IFN-α,提示HPV16 VLP可通過誘導pDC分泌IFN-α來發揮抗病毒的免疫功能。本研究結果發現CD123+pDC在BP皮損組織中的密度較皮損旁正常組織中增高,細胞胞體較大。因此推測pDC可能參與了BP皮損中抗HPV感染的免疫反應,但BP易反復不愈的臨床現狀提示皮損中pDC雖然密度增加但可能未能建立有效的免疫應答。
TLR作為一種天然免疫模式識別受體,通過與相關病原分子結合,引發一系列細胞間信號轉導、炎癥介質的釋放,在機體誘導與調節天然免疫以及獲得性免疫過程中發揮重要作用。pDC高表達TLR7和TLR9[7],TLR7主要識別病毒ssRNA,TLR9主要識別病毒CpG-ODN片段,可通過髓樣分化因子88(MyD88)依賴或非依賴途徑,活化核轉錄因子kappa B(NF-κB)或干擾素調節因子(IRF)家族(IRF-3、5、7),進而調節一系列炎癥性細胞因子,最終影響pDC產生大量的 IFN-α,從而產生抗病毒反應[8]。Daud II等[9]研究顯示在HPV16感染的宮頸上皮中,TLR7、TLR9的表達升高及相關細胞因子的誘導對于HPV16病毒的清除具有重要作用。Cannella F等[10]研究顯示TLR9在HPV感染的宮頸黏膜中表達升高,在HPV的持續感染者TLR9的表達更高,HPV 16可以影響TLR9的轉錄,TLR9的升高如未能使HPV得以清除有可能提示患者向宮頸癌發展的可能性大。本研究結果顯示,BP皮損真皮中TLR7和TLR9染色均較深,表達強度高于相應皮損旁正常真皮,與上述Daud II及Cannella F等研究報道的在HPV感染相關病變組織中TLR7、TLR9表達升高結果一致。推測HPV感染后局部皮損組織處于高水平免疫激活狀態,盡管TLR7和TLR9在參與機體識別HPV或在抗病毒免疫應答、抑制HPV免疫逃逸中發揮重要作用,但可能未能扭轉其他機制所致的局部免疫功能低下,而導致HPV感染呈持續狀態。
Pearson相關性分析顯示,BP皮損及皮損旁正常組織中,pDC細胞密度與真皮中TLR7表達強度之間、pDC細胞密度與真皮中TLR9表達強度之間均存在直線正相關,提示BP皮損真皮中TLR7和TLR9的表達升高可能與皮損局部pDC密度增高有關,由此推測BP皮損局部pDC未能發揮有效的細胞免疫功能可能不是因為pDC未能充分表達TLR7和TLR9的原因而造成,但具體原因尚需進一步研究闡明。
