人源CerS2重組腺病毒構(gòu)建及其對肝癌細(xì)胞HepG2細(xì)胞周期的作用
2018-11-28 02:46:50楊小理歐陽旭紅向加林王鳳學(xué)
重慶醫(yī)學(xué) 2018年31期
關(guān)鍵詞:肝癌
楊小理,錢 玥,歐陽旭紅,向加林,楊 艷,王鳳學(xué)
(1.遵義醫(yī)學(xué)院附屬醫(yī)院醫(yī)學(xué)檢驗(yàn)科,貴州遵義 563003;2.遵義醫(yī)學(xué)院檢驗(yàn)醫(yī)學(xué)院2015級,貴州遵義 563003; 3.貴州省遵義市第一人民醫(yī)院檢驗(yàn)科 563003)
神經(jīng)酰胺合酶(ceramide synthase,CerS)基因家族成員主要包括CerS1、CerS2、CerS4、CerS5、CerS6[1-3],各成員在真核生物中呈現(xiàn)出嚴(yán)格的保守性[4]。CerS2參與神經(jīng)酰胺的合成[5],神經(jīng)酰胺作為鞘磷脂信號途徑中重要的第二信使效應(yīng)分子,參與激活多種蛋白激酶和蛋白磷酸酶調(diào)節(jié)凋亡、增殖、分化和生長停滯等信號過程[6]。研究表明,CerS2在肝癌、乳腺癌、前列腺癌、膀胱癌細(xì)胞株中具有抑制腫瘤細(xì)胞增殖、侵襲的活性[7-10],且較多研究關(guān)注CerS2在腫瘤微環(huán)境的作用及機(jī)制探討。而CerS對動物來說是很重要的相關(guān)基因,其變異將導(dǎo)致嚴(yán)重的代謝紊亂,但CerS2在腫瘤與代謝中的作用卻鮮有報道。本研究通過構(gòu)建人源CerS2重組腺病毒,觀察其對肝癌細(xì)胞HepG2的轉(zhuǎn)染、介導(dǎo)CerS2基因的表達(dá)能力及對細(xì)胞周期的影響,為進(jìn)一步研究CerS2基因?qū)epG2細(xì)胞生長及肝糖、脂質(zhì)代謝可能的影響奠定基礎(chǔ)。
1 材料與方法
1.1材料與試劑 限制性內(nèi)切酶、感受態(tài)大腸埃希菌DH5α、引物合成、RT-PCR、QPCR試劑盒均購自大連寶生物Takara公司;HEK293購自上海中科院細(xì)胞庫,AdMax重組腺病毒表達(dá)載體購自加拿大Microbix公司;DMEM、胎牛血清、胰酶均購自Gibico公司;脂質(zhì)體2000購自Invitrogen公司;質(zhì)粒提取試劑盒、膠回收試劑盒、DNA純化試劑盒均購自O(shè)mega公司;全蛋白提取試劑盒購自凱基生物技術(shù)有限公司;蛋白Marker為Fermentas產(chǎn)品,內(nèi)參抗體為Anti-β-actin-HRP-conjugated,購自康成;Anti-Ceramide synthase 2抗體、Anti-p27 kip 1抗體購自Abcam公司。
1.2方法
1.2.1人源CerS2基因的擴(kuò)增 從Genebank檢索人源基因序列即NM_181746,送公司化學(xué)合成CDS區(qū)全長片段。取1 μL作為模板,用PrimerSTAR進(jìn)行目的片段PCR擴(kuò)增,上游引物:5′-CGT AGA ACG CAG ATC GAA TTC GCC ACC ATG CTC CAG ACC TTG TAT GA-3′;下游引物:5′-CCA TGG TGG CAA GCT TGT CAT TCT TAC GAT GGT TGT TAT T-3′。
1.2.2穿梭質(zhì)粒pDC315-CerS2-GFP的構(gòu)建及重組質(zhì)粒的篩選與鑒定 采用1.0%的瓊脂糖電泳PCR產(chǎn)物,對PCR產(chǎn)物進(jìn)行回收、純化,同時將pDC315-綠色熒光蛋白(GFP)、PCR產(chǎn)物線性化,并經(jīng)In-Fusion交換酶連接、轉(zhuǎn)化、選取陽性克隆經(jīng)PCR鑒定且送大連寶生物測序。
1.2.3重組腺病毒pDC315-CerS2的包裝 將穿梭質(zhì)粒pDC315-CerS2與Admax系統(tǒng)中pBHGloxpΔE1,3cre載體共轉(zhuǎn)染293細(xì)胞,觀察細(xì)胞狀態(tài),第14天左右出現(xiàn)細(xì)胞病變(cytopathic effect,CPE),待所有細(xì)胞都變圓并大部分開始脫落時,收集上清液和細(xì)胞,離心后保留細(xì)胞和2 mL上清液,用液氮和37 ℃水浴鍋反復(fù)凍融樣品3次,離心去沉淀。加入10% 無菌甘油,過濾后得到病毒毒種,保存在-80 ℃。
1.2.4Adv-CerS2-GFP的擴(kuò)增、純化與滴度鑒定 待293細(xì)胞接近長滿時,加入毒種1 mL,當(dāng)大部分細(xì)胞出現(xiàn)CPE且脫落時,收集病毒上清液,再次擴(kuò)增,重組腺病毒經(jīng)Adeno-X Virus Purification Kit純化試劑盒純化,保存在-80 ℃。采用終點(diǎn)稀釋法,計(jì)算病毒滴度。
1.2.5Adv-CerS2-GFP在肝癌細(xì)胞HepG2中的表達(dá)及亞細(xì)胞定位 在6孔板中細(xì)胞爬片,將Adv-GFP(空載組)、Adv-CerS2-GFP(實(shí)驗(yàn)組)分別感染HepG2細(xì)胞,并設(shè)HepG2空白對照組,感染48 h后,PBS洗滌3次,4%多聚甲醛固定30 min,并采用DAPI染核,激光掃描共聚焦顯微鏡觀察與拍照,藍(lán)光激發(fā)和發(fā)射波長分別為340 nm/488 nm,GFP激發(fā)和發(fā)射波長為488 nm/525 nm。
1.2.6RT-qPCR和Western blot法檢測各組細(xì)胞CerS2、p27的表達(dá) 感染48 h后采用Trizol分別提取各組細(xì)胞的總RNA,逆轉(zhuǎn)錄合成cDNA,采用SYBR GreenⅠ熒光染料RT-qPCR二步法檢測CerS2 mRNA表達(dá)水平,以β-actin基因作為內(nèi)參,用2-△△CT值表示基因相對表達(dá)量。所用引物β-actin上游:5′ -TTG CGT TAC ACC CTT TCT T-3′,下游:5′ -GTC ACC TTC ACC GTT CCA-3′;CerS2上游:5′-ATC GTC TTC GCC ATT GTT-3′,下游:5′-CGG TCA CTG CGT TCA TCT-3′;p27 kip1上游:5′-GGC TAA CTC TGA GGA CAC-3′,下游:5′-TTC TTC TGT TCT GTT GGC-3′。采用全蛋白提取試劑盒提取各組細(xì)胞總蛋白,BCA法蛋白定量,取蛋白30~50 μg行10%十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(SDS-PAGE)轉(zhuǎn)移至PVDF膜、封閉、一抗(分別為anti-CerS2 antibody、anti-p27 kip1 antibody)4 ℃孵育過夜、洗膜,二抗室溫孵育1~2 h,洗膜并顯色成像,比較條帶灰度值與灰度比。
1.2.7流式細(xì)胞術(shù)PI法檢測各組細(xì)胞周期的改變 腺病毒轉(zhuǎn)染48 h后,收集、調(diào)整細(xì)胞濃度約1×106個/mL,70%冷乙醇500 μL、4 ℃下固定2 h,100 μL RNase A 37 ℃水浴30 min,再加入400 μL PI染色混勻,4 ℃避光30 min;上機(jī)檢測,記錄激發(fā)波長488 nm處紅色熒光。
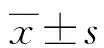
2 結(jié) 果
2.1CerS2目的基因擴(kuò)增的PCR產(chǎn)物及重組穿梭質(zhì)粒pDC315-CerS2-GFP的鑒定 將PCR擴(kuò)增得到的產(chǎn)物回收、經(jīng)EcoRⅠ與KpnⅠ雙酶切后取2 μL電泳鑒定,所得到的片段大小約為1 142 bp左右,與目的片段cDNA大小一致。重組穿梭質(zhì)粒pDC315-CerS2-GFP測序結(jié)果顯示,所克隆片段與NM_18746序列CDS區(qū)完全一致(圖1)。病毒滴度為1.0×109PFU/mL。

圖1 Lass2重組腺病毒測序結(jié)果(僅展示部分)
2.2Cers2亞細(xì)胞定位 激光掃描共聚焦結(jié)果顯示,在細(xì)胞核周圍呈現(xiàn)藍(lán)、綠色重疊,少量分布在細(xì)胞質(zhì)內(nèi),見圖2。

圖2 CerS2在HepG2肝癌細(xì)胞中的表達(dá)(亞細(xì)胞定位)
2.3各組HepG2肝癌細(xì)胞GerS2表達(dá)水平比較 對照組、空載組CerS2表達(dá)比較,差異無統(tǒng)計(jì)學(xué)意義(P>0.05);感染48 h的實(shí)驗(yàn)組比對照組CerS2 mRNA相對表達(dá)量增加了129.79倍(P<0.01),蛋白表達(dá)水平也明顯增加(P<0.01),見圖3。
2.4各組細(xì)胞周期的變化 流式細(xì)胞儀PI法結(jié)果顯示,與對照組、空載組比較,實(shí)驗(yàn)組細(xì)胞G0/G1期百分比明顯增加,G2/M、S期百分比均明顯減少(P<0.01),見表1、圖4。

a:P<0.05,與實(shí)驗(yàn)組比較
圖3各組HepG2肝癌細(xì)胞CerS2表達(dá)水平比較

圖4 各組細(xì)胞周期分析結(jié)果

表1 各組細(xì)胞周期百分比比較
a:P<0.01,與實(shí)驗(yàn)組比較
2.5各組HepG2細(xì)胞p27 kip1表達(dá)水平比較 實(shí)驗(yàn)組p27 kip1 mRNA表達(dá)水平較對照組、空載組明顯上調(diào)(P=0.026、0.019),且p27 kip1蛋白表達(dá)水平也明顯高于對照組及空載組(P<0.01),見圖5。

a:P<0.05,與實(shí)驗(yàn)組比較
3 討 論
CerS2基因是在肝癌相關(guān)基因大規(guī)模功能篩選時發(fā)現(xiàn)的與腫瘤轉(zhuǎn)移相關(guān)的新基因,研究表明其含保守的TLC功能域和特定的HOX功能域,HOX作為序列特異性DNA結(jié)合轉(zhuǎn)錄調(diào)節(jié)因子,在調(diào)控基因轉(zhuǎn)錄方面有著非常重要的作用,其對神經(jīng)酰胺的合成是必需的,早期研究發(fā)現(xiàn)HOX域與細(xì)胞周期調(diào)控密切關(guān)聯(lián),通過細(xì)胞周期影響細(xì)胞生長[6]。此外,CerS2在結(jié)構(gòu)上包含了一個看家基因的典型特征,這是該家族其它成員所沒有的,其在各類腫瘤中抗腫瘤的具體機(jī)制仍不清楚。
本研究應(yīng)用Admax系統(tǒng)構(gòu)建重組腺病毒表達(dá)載體,該包裝系統(tǒng)利用了重組酶系統(tǒng)(Cre-loxP和FLP-frt),操作簡單且重組效率高、獲得的病毒產(chǎn)率高、目的基因的表達(dá)水平高,采用的增強(qiáng)型綠色熒光強(qiáng)度較普通熒光強(qiáng)度提升且保留了熒光蛋白功能的同時能快速有效的對融合的目的基因進(jìn)行報告。經(jīng)酶切、測序鑒定,CerS2重組腺病毒構(gòu)建成功,并在HepG2肝癌細(xì)胞中形成高效過表達(dá),亞細(xì)胞定位結(jié)果提示CerS2融合蛋白在細(xì)胞核周圍表達(dá),少量在細(xì)胞質(zhì)。該重組腺病毒的構(gòu)建為本課題進(jìn)一步研究CerS2在腫瘤代謝中的作用及機(jī)制奠定了基礎(chǔ)。
幾乎所有腫瘤細(xì)胞的生物學(xué)特性都是由于細(xì)胞周期紊亂所導(dǎo)致的失控性生長,即腫瘤細(xì)胞的生長依賴于細(xì)胞周期的正常更替[11]。G1/S期是細(xì)胞周期的主要限制點(diǎn),其中細(xì)胞在G1期完成必要的生長和物質(zhì)準(zhǔn)備,在S期完成其遺傳物質(zhì)--染色體DNA的復(fù)制, G2期進(jìn)行必要的檢查及修復(fù),以確保DNA復(fù)制的準(zhǔn)確性,然后在M期完成遺傳物質(zhì)到子細(xì)胞中的均等分配,并使細(xì)胞一分為二,而G0期是指細(xì)胞處于阻滯的狀態(tài),是脫離細(xì)胞周期、暫時停止分裂的一個階段。本研究中流式細(xì)胞儀分析細(xì)胞周期各時相的百分比結(jié)果顯示CerS2基因抑制了肝癌細(xì)胞的生長,使細(xì)胞大量處于G0/G1期,阻滯肝癌細(xì)胞HepG2進(jìn)入下一階段的增殖及分裂。
為進(jìn)一步探討其可能的作用機(jī)制,本實(shí)驗(yàn)觀察了上調(diào)的CerS2對細(xì)胞周期調(diào)節(jié)蛋白p27表達(dá)的影響。p27蛋白是CIP/KIP家族成員之一,是細(xì)胞增殖一個主要的負(fù)調(diào)控因子,通過結(jié)合及調(diào)控各種細(xì)胞周期素(Cyclin)-細(xì)胞周期素依賴性激酶(CDK)包括Cyclin D(D1,D2,D3)-CDK4/CDK6、Cyclin E-CDK2復(fù)合物[11-12],從而調(diào)控細(xì)胞進(jìn)入G1期和S期[13],其調(diào)控腫瘤細(xì)胞周期的同時能抑制腫瘤細(xì)胞的分裂,是負(fù)性調(diào)控細(xì)胞周期的蛋白[14]。p27被認(rèn)為是人類多種腫瘤獨(dú)立的預(yù)后因子和未來可能的腫瘤治療靶點(diǎn)。有研究報道,CDK抑制結(jié)構(gòu)域位于p27蛋白的N-末端部分,該結(jié)構(gòu)域使細(xì)胞足以停滯于G0/G1期,但p27可能在不同的細(xì)胞與不同分子相互作用,在適當(dāng)?shù)臅r間到達(dá)適當(dāng)?shù)募?xì)胞內(nèi)位置發(fā)揮其生物學(xué)活性,呈現(xiàn)出對細(xì)胞周期不同的調(diào)控[15]。本研究結(jié)果提示上調(diào)的CerS2基因促進(jìn)p27蛋白表達(dá)增強(qiáng),這可能是肝癌細(xì)胞HepG2細(xì)胞周期G0/G1期阻滯的可能機(jī)制之一,但其具體的分子生物學(xué)機(jī)制如過表達(dá)的CerS2是通過何細(xì)胞調(diào)控因子或信號調(diào)控網(wǎng)絡(luò)導(dǎo)致p27上調(diào)及p27通過抑制何種Cyclin-CDK復(fù)合物導(dǎo)致細(xì)胞周期的改變等等尚待后續(xù)深入研究。
猜你喜歡
天津醫(yī)科大學(xué)學(xué)報(2019年3期)2019-08-13 06:53:08
中成藥(2016年8期)2016-05-17 06:08:14
癌癥進(jìn)展(2016年12期)2016-03-20 13:16:17
罕少疾病雜志(2016年5期)2016-03-11 16:34:44
吉林大學(xué)學(xué)報(醫(yī)學(xué)版)(2015年1期)2015-12-17 07:47:28
腫瘤預(yù)防與治療(2015年1期)2015-09-26 07:26:20
中國當(dāng)代醫(yī)藥(2015年16期)2015-03-01 02:03:11
中國醫(yī)藥導(dǎo)報(2015年26期)2015-02-28 22:07:59
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:34
肝膽胰外科雜志(2015年4期)2015-02-27 11:12:24
