核桃內果皮苯丙氨酸解氨酶基因克隆及表達分析
2018-11-30 06:43:36郭永翠張建良
西南農業學報 2018年10期
郭永翠,朱 玲,高 山,張建良,張 銳,2*
(1.塔里木大學植物科學學院, 新疆 阿拉爾 843300; 2.塔里木大學新疆生產建設兵團塔里木盆地生物資源保護利用重點實驗室, 新疆 阿拉爾 843300; 3.新疆伊犁州農業科學研究所, 新疆 伊寧 835000; 4.新疆生產建設兵團第一師三團, 新疆 阿拉爾 843300)
【研究意義】核桃(JuglansregiaL.)在中國栽培歷史悠久,因其富含蛋白質、脂肪、碳水化合物等人體所必需物質及鐵、鋅、錳等多種微量元素,對于延緩人體衰老、心血管的保健作用、預防慢性疾病等具有非凡意義,有著極佳的保健效果和藥用價值,在國內國際市場均具有巨大的消費潛力[1-4]。【前人研究進展】新疆核桃殼薄、早實豐產、出仁率高且種仁飽滿,其中紙皮核桃品質尤為突出,成為近年成為消費熱點[5]。‘紙皮’核桃殼厚小于1.0 mm (大多在0.5 ~0.8 mm),出仁率達65.0 %以上,種仁飽滿,內果皮發育完整,其中‘溫138’核桃是‘紙皮’核桃栽培園中發現的突變株,其平均出仁率高達83.2 %,產量極高,果殼極薄、極易脫皮,但殼面(內果皮發育不完整)開裂導致露仁,是適于核桃深加工的優質種質資源[6-7]。木質素(lignin)是核桃內果皮(硬殼)的主要成分且是內果皮中難分解的大分子有機物質,其沉積出現在次生加厚的細胞壁中以提高細胞壁的強度及硬度,對核桃內果皮發育影響較大[8-9]。苯丙氨酸解氨酶(phenylalanine ammonia-lyase,簡稱PAL)存在于一些菌類、藻類及所有綠色植物中,是參與木質素生物合成過程中苯丙烷類代謝途徑的首個關鍵酶,在多種植物的木質化組織中含有較高PAL活性,PAL基因對木質素的合成起重要作用[10-11]。目前已從辣椒(CapsicumannuumL.)[12]、當歸(Angelicasinensis)[13]、馬尾松(Pinusmassoniana)[14]、黃瓜(CucumissativusL.)[15]等多種植物中成功獲得了PAL基因并展開相關研究。【本研究切入點】經研究發現,植物中PAL基因對木質素的合成起重要作用,在轉基因煙草(NicotianatabacumL.)植株中抑制PAL的表達將導致木質素等相關次生代謝產物含量的下降,但植株生長出現不正常現象[16]。不同PAL基因在擬南芥(Arabidopsisthaliana)植株中各個部位的表達量不同,表明不同的PAL基因具有不同生物活性和表達部位[17];其在不同品種龍眼(EuphorialonganL.)葉片中的表達量存在顯著差異[18];且在獨一味(Lamiophlomisrotata)的不同組織中表達有差異,表達量由高到低分別是葉片、根部、葉柄、莖[19]。【擬解決的關鍵問題】目前關于‘溫138’核桃露仁現象及其與之相關的核桃內果皮硬化過程中木質素沉積的系統分子機理尚不明確,本研究以‘溫138’、‘紙皮’核桃為研究對象,對其內果皮中木質素相關PAL基因進行克隆及生物信息學分析,為運用基因工程改良核桃果實品質提供理論依據。
1 材料與方法
1.1 材料
供試材料露仁種質‘溫138’核桃及硬殼發育完整‘紙皮’核桃,均采自新疆維吾爾自治區溫宿縣核桃木本糧油林場(圖1)。在果實膨大期后期6月7日(約花后51 d)至硬核期7月26日(約花后100 d)采樣,采樣覆蓋整個核桃內果皮硬化期。將核桃內果皮剝離存于-80 ℃超低溫冰箱中。
植物總RNA提取采用E.Z.N.A Plant RNA特殊植物RNA提取試劑盒(Omega公司);cDNA第一鏈合成采用EasyScript One-Step gDNA Removal and cDNA Synthesis SuperMix(100rxns)試劑盒(全式金公司);膠回收采用Easy Pure?Quick Gel Extraction試劑盒(全式金公司);qRT-PCR檢測采用SYBR?Premix ExTaqTMGC(Perfect Real Time染料法)試劑盒(TaKaRa公司)。
1.2 核桃內果皮總RNA提取
利用植物總RNA提取試劑盒提取核桃內果皮中總RNA。用1 %瓊脂糖凝膠電泳方法檢測及鑒定PCR產物的濃度、純度和亮度。采用cNDA合成試劑盒反轉錄合成cDNA。
1.3 引物設計
根據核桃轉錄組數據庫和NCBI數據庫已發表的核桃PAL基因序列(登錄號:JX069977.1)設計引物,‘溫138’核桃引物PAL-F1:5′-CCCCTCCAAACAACCCAA-3′及PAL-R1:5′-CCTTCAGACAATCCAGCA-3′,‘紙皮’核桃引物PAL-F2:5′-ACCCAATCCAAGAAAGAA-3′及PAL-R1:5′-CCTTCAGACAATCCAGCA-3′。引物由上海生工生物技術有限責任公司合成。

A:‘溫138’核桃硬殼發育情況, B: ‘紙皮’核桃硬殼發育情況A: ‘Wen138’ walnuts shell development, B: ‘Zhipi’ walnuts shell development圖1 ‘溫138’和‘紙皮’核桃硬殼發育情況Fig.1 ‘Wen138’ walnuts and ‘Zhipi’ walnuts shell development
1.4 核桃內果皮PAL基因的克隆及測序
RT-PCR反應體系(20 μl)及條件為:模板3 μl,上、下游引物(10 μM)各0.5 μl,2×EasyTaq?PCR SuperMix 10 μl,最后加滅菌ddH2O6μl。擴增條件:94 ℃/3 min(預變性);94 ℃/45 s(變性),57 ℃/45 s(退火),72 ℃/2 min(延伸),31個循環;72 ℃/8 min(繼續延伸);4 ℃保存。用1 %瓊脂糖凝膠電泳檢測PCR產物并成像,后將擴增的目的條帶進行回收并連接到T載體,篩選出陽性克隆測序。
1.5 生物信息學分析
1.5.1 核苷酸、氨基酸序列分析 將陽性克隆的測序結果及推導氨基酸序列采用NCBI數據庫BLAST功能(https://blast.ncbi.nlm.nih.gov/Blast.cgi)進行在線PAL序列匹配度、同源性分析;ORF Finder (https://www.ncbi.nlm.nih.gov/orffinder/)檢測cDNA序列的開放閱讀框;PAL基因推導氨基酸序列采用Clustal X 1.08軟件進行多序列比對;PAL蛋白功能結構域利用NCBI數據庫Conserved Domains (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi)進行在線分析。
1.5.2 理化性質分析 運用ExPASy數據庫ProtParam功能(http://web.expasy.org/protparam/)分析核桃內果皮WJ-PAL蛋白及ZJ-PAL蛋白的相對分子質量、氨基酸數量及理論等電點等相關理化性質。
1.5.3 蛋白質二級結構及三維結構預測 運用EBI數據庫InterProScan功能(http://www.ebi.ac.uk/interpro/search/sequence-search)在線分析及預測核桃內果皮PAL蛋白保守區域、采用PredictProtein在線平臺(https://www.predictprotein.org/)分析PAL蛋白二級結構,選用在線同源服務軟件SWISS-MODEL (http://swissmo del.expasy.org/)預測PAL蛋白質三維結構。
1.5.4 系統進化樹 采用NCBI數據庫BLAST在線比對,選取與克隆基因相似性較高的植物,采用MAGA5.1軟件(Neighbor-Joining法)構建PAL基因與其它相似性高植物的系統進化樹。
1.6 PAL基因的表達分析
以18S為內參,采用實時熒光定量PCR (qRT-PCR)檢測PAL基因的相對表達量。根據已克隆的核桃內果皮木質素PAL基因序列,按照標準熒光定量PCR引物原則,分別設計‘溫138’、‘紙皮’核桃PAL基因熒光定量PCR引物,露仁種質‘溫138’核桃PCR引物PAL-WF:5′-AGGTGAAACGCATGGTGG-3′,PAL-WR:5′-CCTGTGATATGGTCAGGGTCTT-3′,硬殼完整種質‘紙皮’核桃PCR引物PAL-ZF:5′-CCGTTGTTGACGCTGTTTAT-3′,PAL-ZR:5′-GGGAG ACCCTGACCATTTC-3′,內參基因引物:NBQ-F:5′-AGTCGTAACAAGGTTTCCGTAGGT-3′;NBQ-R: 5′-GCTGGGCAGGTATCGACAAT-3′。以‘溫138’及‘紙皮’核桃8個時期的cDNA為模板,采用兩步法進行Real-Time PCR擴增,反應體系:cDNA模板1 μl,正、反引物(10 μM)各0.2 μl,SYBR?Premix ExTaqTMGC5 μl,最后加滅菌ddH2O至10 μl。擴增條件:95 ℃/30 s,95 ℃/5 s,40個循環;熔解曲線生成步驟:58 ℃/30 s,72 ℃/30 s,95 ℃/15 s,55 ℃/15 s;55 ℃/20 min;95 ℃/15 s。利用2-△△Ct方法[20]分別計算PAL基因在硬殼完整和露仁核桃內果皮發育過程中的表達量。
2 結果與分析
2.1 總RNA的提取及檢測
核桃內果皮中總RNA凝膠成像呈現較清晰完整的28S rRNA和18S rRNA條帶,5S rRNA條帶較模糊,表明所得的總RNA有較好的完整性,28S rRNA的亮度是18S rRNA的兩倍,測得核酸濃度A260/A280為1. 90~2. 20,說明提取的總RNA有較高的純度(圖2)。
2.2 核桃內果皮PAL基因的克隆
以‘溫138’和‘紙皮’核桃內果皮總RNA反轉錄后的cDNA為模板,通過所設計的引物(PAL-F1、PAL-R1和PAL-F2、PAL-R1)進行RT-PCR反應(圖3),得到露仁及硬殼完整核桃PAL基因序列,分別命名為WJ-PAL和ZJ-PAL(圖4)。WJ-PAL基因片段長度為1000 bp (含有600 bp的ORF),可編碼200個氨基酸,經分析發現其分子量為21.11 kD,理論等電點為8.44,WJ-PAL屬于堿性蛋白;ZL-PAL基因片段長度為1000 bp (含有690 bp的ORF),可編碼230個氨基酸,經分析發現其分子量為24.10 kD,理論等電點為7.44,ZJ-PAL屬于弱堿性蛋白。
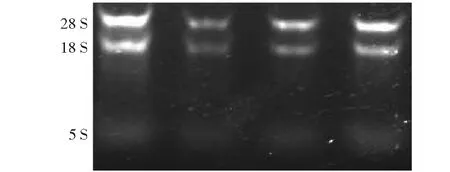
圖2 電泳檢測核桃內果皮總RNAFig.2 Electrophoresis of total RNA from walnut endocarp
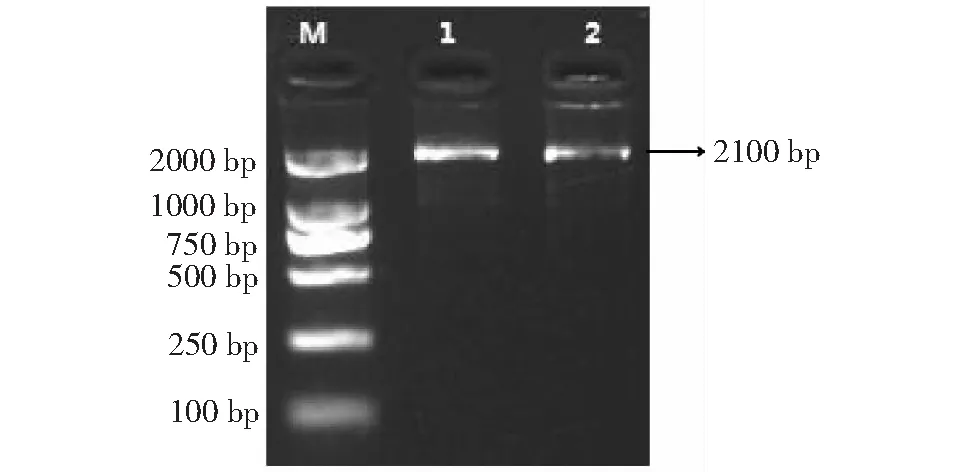
M: DL2000 DNA Marker; 1: ‘溫138’核桃; 2: ‘紙皮’核桃M: DL2000 DNA Marker; 1: ‘Wen138’ walnut; 2: ‘Zhipi’ walnut圖3 核桃內果皮PAL基因的保守區PCR產物Fig.3 PCR product of PAL gene conservative region in walnut endocarp
2.3 生物信息學分析
2.3.1 序列分析 將得到的測序結果與NCBI數據庫己知植物PAL基因進行序列同源性比對的結果(圖5)表明,WJ-PAL與ZJ-PAL核苷酸序列及氨基酸推導序列均與其它植物PAL基因序列有較高的匹配度。
將本研究獲得的2個基因進行相似性分析,結果顯示WJ-PAL和ZJ-PAL核苷酸序列與黑核桃、桃、日本杏等物種的PAL基因序列匹配度最高,其同源性均可達到77 %~96 %,其中均與黑核桃(AJ278454.1)的匹配度最高,WJ-PAL、ZJ-PAL與黑核桃同源性分別達到89 %、96 %。其相似性經NCBI數據庫BLASTp在線分析可知,WJ-PAL和ZL-PAL
與其它植物推導氨基酸序列相似性均在79 %以上。PAL推導氨基酸序列發現WJ-PAL與ZL-PAL均存在Phenylalanine ammonia-lyase and histidine ammonia-lyase (PAL-HAL)、PPK09367、huth及HutH 4個非特異性位點,并且含有phe_am_lyase、芳香族裂解酶的Lyase_aromatic及PAL (PLN02457) 3個功能結構域,WJ-PAL與ZL-PAL為多結構域蛋白,屬于裂解酶I (Lyase_I_Like)超家族(圖5)。
2.3.2 系統進化樹分析 將本研究獲得的兩個基因核苷酸序列進行多重序列比對(圖6),露仁種質‘溫138’克隆所得WJ-PAL基因和硬殼完整品種‘紙皮’克隆所得ZJ-PAL基因序列一致性顯示只有54.76 %。
經過在線比對,發現核桃WJ-PAL及ZJ-PAL基因與其它植物的PAL基因具有較高的相似性,選取與WJ-PAL及ZJ-PAL相似性較高的十九種植物構建核桃PAL基因與其它植物PAL基因的系統進化樹(圖7)。結果顯示,核桃與喬木類植物親緣關系較近,‘紙皮’核桃與黑核桃聚為一支,川楊、顫楊、胡楊、毛果楊等楊樹較好的聚為一支,鹽膚木與芒果這兩個漆樹科植物聚為同一支,蕓香科柑橘屬的檸檬與甜橙聚為同一支,葡萄科的葡萄與桑科的桑樹較好的聚為一支,并且自展值均達到100,其它植物也各自與相近的植物聚為一支。硬殼完整核桃‘紙皮’(ZJ-PAL)與黑核桃(Juglansnigra)聚在一起,卻與露仁核桃種質‘溫138’ (WJ-PAL)未能聚為同一支,這說明就PAL基因而言,硬殼完整核桃‘紙皮’與黑核桃聚為一支說明其親緣關系較近,而露仁種質‘溫138’核桃卻未與黑核桃及其它同源植物聚在一起說明有較遠的親緣關系。

A: WJ-PAL基因片段測序; B: ZJ-PAL基因片段測序A: Sequencing result of WJ-PAL gene fragment; B: Sequencing result of ZJ-PAL gene fragment圖4 核桃PAL基因片段測序Fig.4 Sequencing result of PAL gene fragment in walnut

A: WJ-PAL基因的結構域分析; B: ZJ-PAL基因的結構域分析A: Domain analysis of WJ-PAL gene; B: Domain analysis of ZJ-PAL gene圖5 PAL基因的結構域分析Fig.5 Domain analysis of PAL gene

圖6 WJ-PAL及ZJ-PAL基因多重序列比對Fig.6 Multiple sequence alignment of WJ-PAL and ZJ-PAL gene
2.4 蛋白質二級結構分析
提交WJ-PAL、ZJ-PAL蛋白的氨基酸序列進行在線預測分析(圖8)。分析得到,WJ-PAL蛋白含有58.29 %的無規則卷曲,是WJ-PAL蛋白二級結構中最大量的結構元件,其次含有35.68 %的α-螺旋和6.02 %的延伸鏈等結構元件,N-末端存在較多的無規卷曲結構,而C-末端則存在較多的延伸鏈結構。ZJ-PAL蛋白主要含有48.03 %的α-螺旋,是ZJ-PAL蛋白二級結構中最為重要的結構元件,其次含有46.03 %的無規則卷曲結構和5.68 %的延伸鏈。
在線預測WJ-PAL和ZJ-PAL蛋白的保守區域,結果表明這兩個蛋白屬于L-天冬氨酸家族(L-Aspartase-like),WJ-PAL和ZJ-PAL蛋白保守區域包含芳香族氨基酸裂解酶(IPR001106)活性位點。其中ZJ-PAL蛋白具有PAL-HAL信號(含有PAL-HAL活性位點),具備苯丙氨酸解氨酶(PAL)家族蛋白的特征。
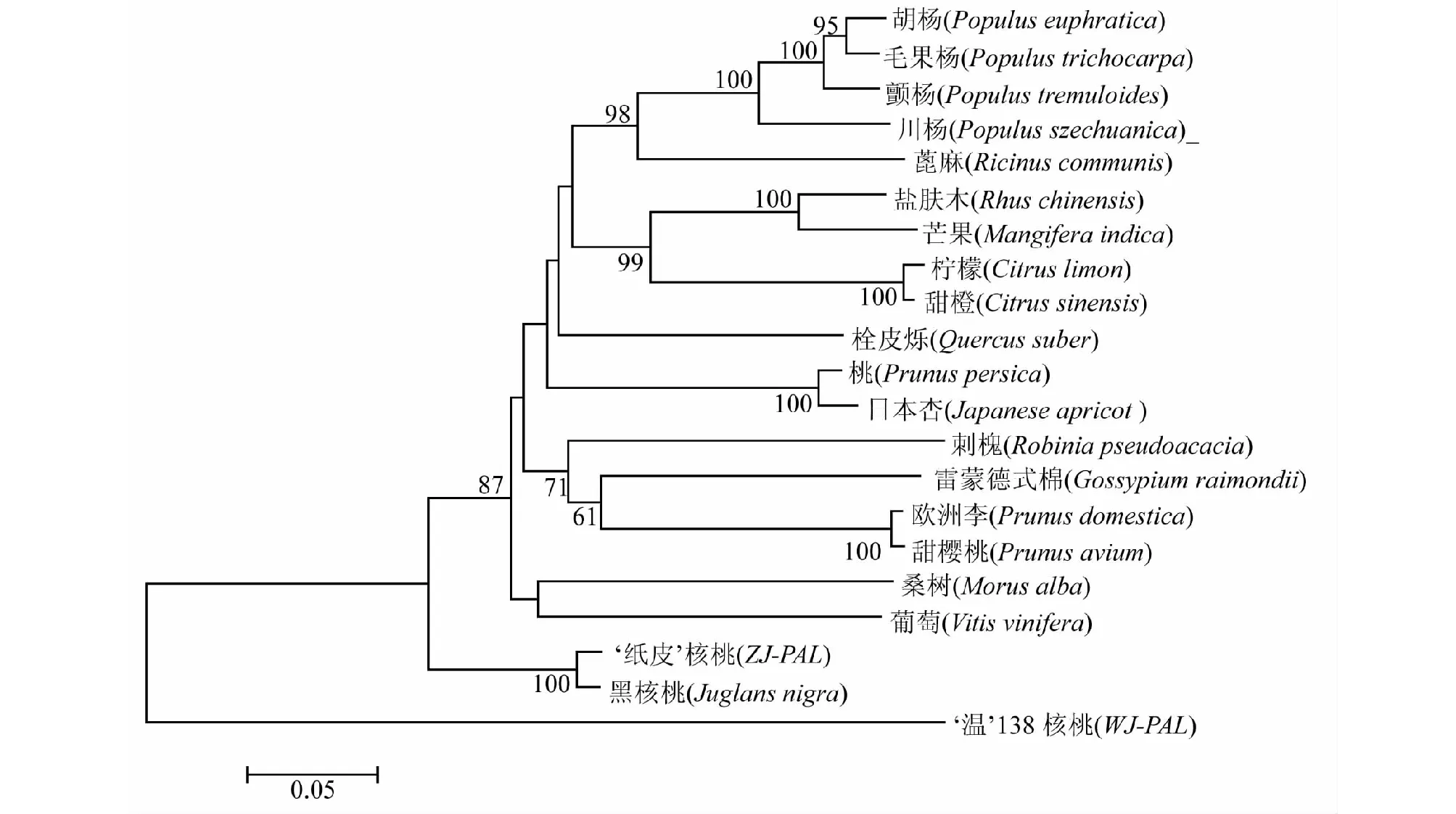
圖7 核桃與18種植物PAL基因系統進化樹分析Fig.7 Phylogenetic tree of PAL gene in walnut and 18 other plants

A: WJ-PAL蛋白二級結構; B: ZJ-PAL蛋白二級結構; 紅色:α-螺旋; 藍色:延伸鏈A: Secondary structure of WJ-PAL protein; B: Secondary structure of ZJ-PAL protein; Red: α-helix; Blue: Extended strand圖8 PAL蛋白二級結構Fig.8 Secondary structure of 7 PAL protein
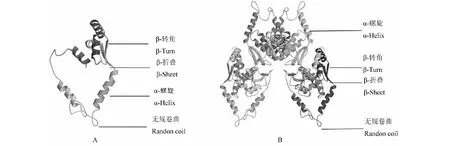
A: WJ-PAL蛋白三級結構; B: ZJ-PAL蛋白三級結構A:The tertiary structure of WJ-PAL protein; B: The tertiary structure of ZJ-PAL protein圖9 PAL蛋白三級結構Fig.9 The tertiary structure of PAL protein

A:WJ-PAL基因相對表達量; B: ZJ-PAL基因相對表達量; W: ‘溫138’核桃; Z:‘紙皮’核桃; W(Z)67: 花后51 d (6月7日); W(Z)613: 花后57 d (6月13日); W(Z)621: 花后65 d (6月21日); W(Z)628: 花后72 d (6月28日); W(Z)74: 花后78 d (7月4日); W(Z)712: 花后86 d (7月12日); W(Z)720: 花后94 d (7月20日); W(Z)726: 花后100 d (7月26日)A: Relative expression of WJ-PAL gene; B: Relative expression of ZJ-PAL gene; W: ‘Wen 138’ walnut; Z: ‘Zhipi’ walnut; W (Z) 67: 51 days after anthesis (June 7th); W (Z) 613: 57 days after anthesis (June 13th); W (Z) 621: 65 days after anthesis (June 21st); W (Z) 628: 72 days after anthesis (June 28th); W (Z) 74: 78 days after anthesis (July 4th); W (Z) 712: 86 days after anthesis (July 12th); W (Z) 720: 94 days after anthesis (July 20th); W (Z) 726: 100 days after anthesis (July 26th)圖10 核桃內果皮硬化過程PAL基因相對表達水平Fig.10 Relative expression level of PAL gene during hardening of walnut endocarp
2.5 蛋白質三級結構預測
分別建立WJ-PAL與ZJ-PAL蛋白三維空間結構模型(圖9)。WJ-PAL與ZJ-PAL這兩種蛋白質三級結構由α-螺旋、β-轉角、β-折疊和無規則卷曲構成,其中均α-螺旋為主要結構元件,其它元件較少。蛋白三級結構分析可知ZJ-PAL較WJ-PAL結構復雜的多,其α-螺旋、β-轉角、β-折疊和無規則卷曲結構等元件含量多。
2.6 WJ-PAL與ZJ-PAL基因相對定量分析
實時熒光定量PCR技術分析WJ-PAL與ZJ-PAL在核桃內果皮硬化過程中的表達水平(圖10)。WJ-PAL基因相對表達量在‘溫138’核桃的8個不同發育時期中有非常明顯的差異。相對表達量最高的花后100 d (117.78)是相對表達量最低的花后51 d (1.00)的117倍。這表明,‘溫138’核桃內果皮硬化過程的前期PAL基因表達量相對較小,至核桃內果皮硬化過程的后期其PAL基因大量表達。在核桃內果皮硬化過程的8個不同發育時期ZJ-PAL基因的相對表達量相較WJ-PAL差異較小,在這8個時期中,相對表達量最高的花后65 d (1.33)是相對表達量最低的花后72 d (0.25)的5倍。PAL基因在露仁核桃‘溫138’內果皮中前期木質素相對表達量均較低,于花后86 d開始增長,在花后100 d達到最高值,是前期的近100倍,硬殼完整核桃‘紙皮’在硬核期整個時期木質素相對表達量變化較平緩,于花后72 d最低,最高出現在花后65 d,整個種皮發育過程中,PAL基因在硬殼完整核桃內果皮中的表達量低于同期露仁核桃內果皮中的表達量。
3 討 論
PAL能夠催化高等植物中苯丙氨酸解氨生成氨氣和反式肉桂酸,是木質素單體合成過程中所需的第一步反應酶[21],其受環境影響較小可用作物種鑒定[22]。目前關于露仁核桃在PAL基因克隆及表達分析方面研究還未見文獻報道,PAL對核桃露仁現象的作用機理還不得而知。本試驗克隆得到露仁核桃種質‘溫138’及硬殼發育完整核桃種質‘紙皮’內果皮PAL基因,分別命名為WJ-PAL與ZJ-PAL,利用生物信息學預測獲得PAL基因的理化性質參數、二級結構、三級結構等,分析PAL基因的進化關系及在核桃內果皮硬化過程中不同時期中的相對表達特性。
WJ-PAL與ZJ-PAL基因均為苯丙氨酸解氨酶基因的家族成員,根據試驗結果得知,WJ-PAL與ZJ-PAL推導氨基酸序列存在相同的非特異性位點(PPK09367, HutH, hutH及PAL-HAL)及功能結構域(PLN02457, phe_am_lyase及Lyase_aromatic),這一結果白樺(Betulaplatyphylla)、美洲南瓜(Cucurbitapepo)相關研究結果吻合[11, 23]。同源性分析顯示核桃露仁種質WJ-PAL基因與其它植物沒有聚為一支,而硬殼發育完整ZJ-PAL基因與同科的植物較好地聚在一起,其與喬木類植物親緣關系較近,其中與黑核桃親緣性最高。WJ-PAL與ZJ-PAL核苷酸序列及其編碼的氨基酸序列與黑核桃PAL基因及其氨基酸序列的相似性分別為89 %和99 %。發現不同植物間PAL基因也有較高的一致性,推測PAL在核桃發育過程中對內果皮(硬殼)完整程度有較大的影響。
本試驗實時熒光定量PCR結果顯示,露仁‘溫138’核桃WJ-PAL在內果皮硬化的8個不同發育時期基因的表達量存在明顯差異,其中前期的相對表達量均較低,在核桃硬核期的花后100 d達到最大值,是前期的近100倍,對照‘紙皮’核桃ZJ-PAL在內果皮硬化的8個時期基因表達量差異較小,在花后51~100 d間相對表達量變化較小,在花后71 d出現最低值,這與薛應鈺的研究有一定的出入。美洲南瓜(Cucurbitapepo)研究發現木質素含量隨種子的成熟顯著降低,其種仁裸露裸仁現象是由于種皮發育過程中厚壁組織中缺乏木質素的合成和積累;時空表達研究發現,裸仁美洲南瓜在授粉后的8 ~18 d種皮中隨木質素的合成和積累PAL酶活性逐漸升高,隨后PAL酶活性明顯降低[24]。在自交授粉20 d后,PAL基因在硬殼完整美洲南瓜種皮中表達量上升,在裸仁美洲南瓜種皮中表達量則下降;其中在同一時期有殼美洲南瓜的表達量遠高于裸仁美洲南瓜中的表達量[11]。本試驗相對定量分析結果發現,PAL基因在硬殼完整核桃內果皮中的表達量低于同期露仁核桃內果皮中的表達量,本結果與美洲南瓜研究有較大的出入。說明在核桃生長發育過程中,PAL基因在露仁核桃中的表達量與酶活性具有一定同步性,‘溫138’核桃內果皮出現露仁現象可能由于前期木質素代謝中間產物形成太少,進一步說明了露仁核桃內果皮形成的原因可能是由于前期缺乏木質素代謝產物的積累。本試驗結果可以看出在硬核期木質素的改變對核桃內果皮發育存在一定影響,核桃出現露仁現象與PAL活性的關系還有待進一步深入研究。
4 結 論
通過基因克隆獲得露仁核桃及硬殼完整核桃內果皮PAL基因,WJ-PAL與ZJ-PAL推導氨基酸序列存在相同的非特異性位點、功能結構域,均屬于Lyase_I_Like超家族,黑核桃序列相似性極高。露仁核桃‘溫138’前期PAL基因表達遠低于后期表達,核桃露仁現象可能由于內果皮硬化過程前期木質素積累不足造成。
