“云新高原”核桃及其親本的葉綠體DNA差異分析
2018-11-30 06:46:40肖良俊陳少瑜陳海云寧德魯
西南農業學報 2018年10期
吳 濤,肖良俊,陳少瑜,陳海云,寧德魯*
(1.云南省林業科學院,云南 昆明 650201;2.云南省木本油料工程技術研究中心,云南 昆明 650201;3.云南省森林植物培育與開發利用重點實驗室/國家林業局云南珍稀瀕特森林植物保護和繁育實驗室,云南 昆明 650201)
【研究意義】云南省是深紋核桃(Juglanssigillata)的主要起源分布中心和栽培區域,截止2015年底已種植282萬hm2,產量達到85萬t,產值為266億元,面積、產量和產值均居全國之首。漾濞泡核桃是云南傳統主栽的深紋核桃品種,但存在結實晚(約10 a左右)、受益慢等問題。據此,云南省林業科學院通過種間雜交途徑,用漾濞泡核桃與早實核桃(J.regia)進行雜交,培育出云新高原等早實雜交新品種核桃,并于2004年通過云南省林木品種審定委員會審定[1]。云新高原核桃的母本漾濞泡核桃樹體高大、樹勢旺,小葉9~13片、多13片,結果枝多為中長果枝,種子刻紋深。云新高原核桃的父本為早實核桃優良單株‘云林A7’樹體中等、樹勢較弱,小葉7~9片、多7片,結果枝以中短果枝為主,種子表面光滑。云新高原樹勢中等,樹姿開張,小葉9~11片、多9片,結果枝多為中果枝,種子刻紋淺、表面較光滑。綜合來看,云新高原的樹高、干徑、冠幅與父本相近,在早結實(1~3年開花結果)、頂芽形狀、腋芽形狀、有無芽距(主副芽是否分開)等習性上偏向于父本,復葉長、小葉數、小葉形狀等性狀介于雙親之間,呈趨中變異。在調查云南省核桃資源中,發現一些兼具深紋核桃和核桃特點的資源。鑒于云南曾于40年代進行過核桃的大面積引種栽培,推測這些資源可能是二者的天然雜交種。為此,希望能通過研究掌握人工雜交種云新高原的葉綠體基因組的遺傳規律,來對這些“疑似雜交種”的身份進行確認。【前人研究進展】葉綠體DNA(cpDNA)屬于細胞質基因,為綠色植物所特有,其遺傳方式有別于核基因的遺傳方式,屬于細胞質遺傳,在被子植物中存在雙親質體遺傳和單親質體遺傳兩種類型,通常以單親母系遺傳方式為主,而父系質體遺傳在裸子植物中較為普遍[2-4]。葉綠體基因組很小,進化較慢,不存在基因重組和結構變異,更加便于遺傳進化研究[5]。近年來,以特定片段測序為代表的第三代DNA分子標記技術,因具特異性強、通量高和高效快捷的優點而受到廣泛的關注和應用[6]。研究者利用小片段葉綠體DNA測序的方法進行遺傳多樣性的研究,取得了較好的結果[7-10]。目前,DNA分子標記已經廣泛應用于核桃遺傳多樣性研究[11],但關于cpDNA標記在核桃遺傳多樣性研究中的應用較少。【本研究切入點】基于核桃葉綠體基因組DNA序列信息,設計核桃cpDNA分子標記引物,并對云新核桃及其親本的葉綠體DNA進行擴增產物的序列檢測、分析和比較。【擬解決的關鍵問題】一是檢測核桃葉綠體基因的遺傳特性,二是開發出可應用于核桃起源及遺傳多樣性研究的分子標記。
1 材料與方法
1.1 試驗材料
云新高原核桃(Juglanssigillata×J.reigacv. ‘Yunxingaoyuan’)、母本漾濞泡核桃(J.sigillatacv. ‘Yangpao’)、父本早實核桃(J.reigacv. ‘Yunlin A7’)的樹齡分別為10年、20年、25年,均種植在云南省林業科學院核桃資源圃內。2017年5月采集健康新鮮葉片,即時移入實驗室進行DNA提取。
1.2 研究方法
1.2.1 總DNA提取 總DNA用CTAB法提取。獲得的總DNA經0.8 %的瓊脂糖凝膠電泳進行質量檢測,用超微量紫外分光光度計ND2000測定濃度和純度,然后稀釋為40 ng/μl,于-20 ℃保存備用。
1.2.2 引物設計 Hu等[12]獲得了核桃屬5個種的葉綠體基因組全長。本試驗以其中的深紋核桃和核桃的cpDNA全長序列為基礎(NCBI登錄號分別為KX424843和KT963008),首先使用DNAMAN對二者的全長序列進行差異分析,然后使用Primer 5.0軟件針對差異位點分別設計引物序列(表1),引物均由昆明碩擎生物科技有限公司合成。
1.2.3 PCR擴增與測序 PCR反應體系總體積20 μl,其中PCR Mixer 10 μl,引物(濃度10 μmol/L)各0.5 μl, DNA模板1 μl,ddH2O 8 μl。PCR擴增反應在擴增儀(Biometra TGradient,Germany)中進行,反應程序:94 ℃預變性4 min;94 ℃變性30 s,58 ℃退火30 s,72 ℃延伸60 s,35個循環;72 ℃延伸7 min。PCR產物用1 %瓊脂糖膠電泳檢測,在凝膠成像儀上觀察并切取目的條帶,使用凝膠回收試劑盒(天根生化科技有限公司,北京)回收,回收產物送昆明碩擎生物科技有限公司采用ABI 3730XL自動測序儀(Applied Biosystems,USA)進行雙向測序。
1.2.4 數據處理與分析 測序結果使用ContigExpress軟件觀察峰圖并進行序列拼接及人工校對,手動刪除序列兩端50~60 bp不可靠的位點,拼接好的序列用 DNAMAN 9.0進行比對和差異位點的分析。
2 結果與分析
2.1 深紋核桃與核桃的葉綠體基因組序列差異分析
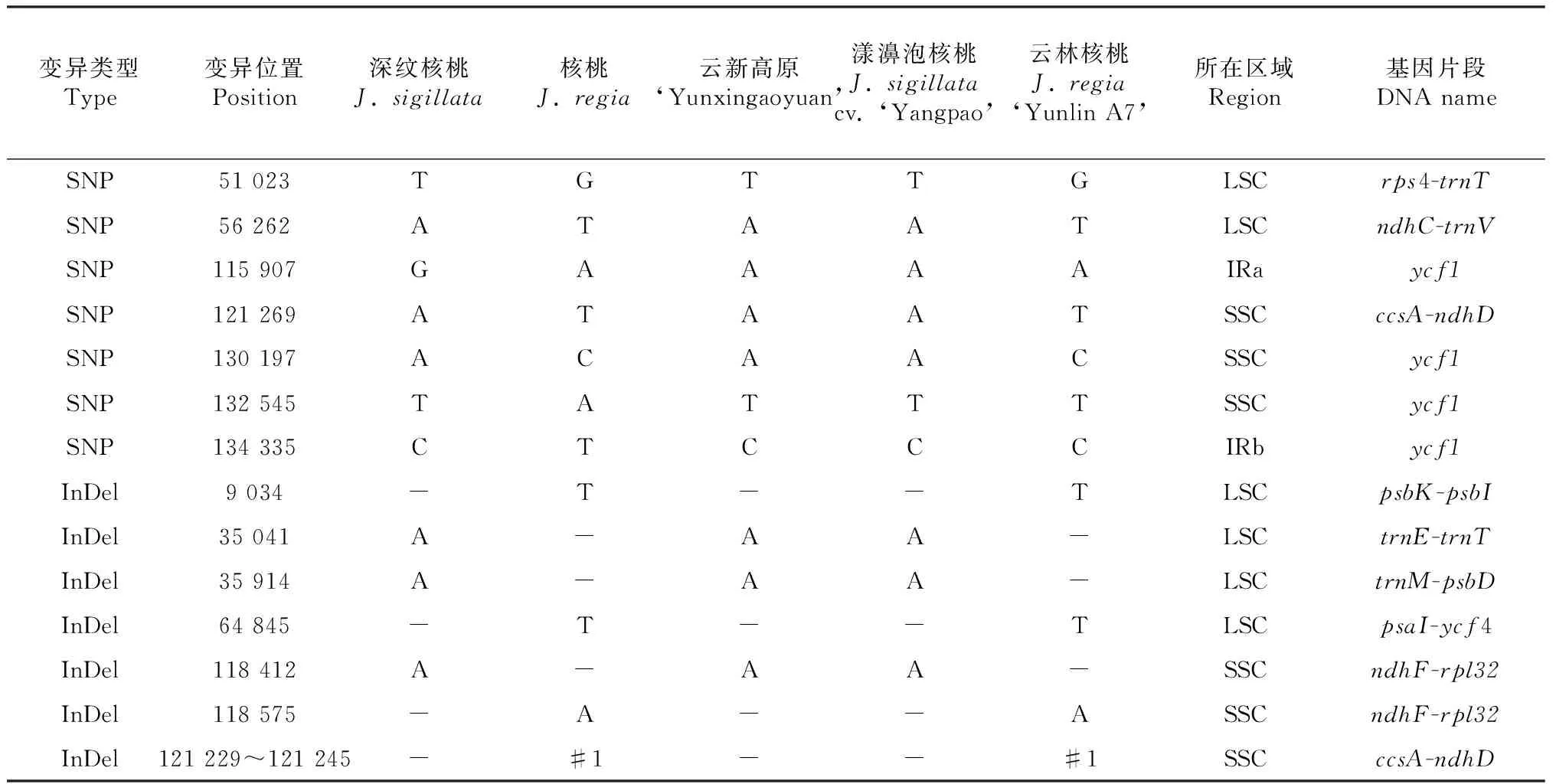
深紋核桃和核桃的葉綠體基因組全序列于2017年成功測序[12],全序列長度分別為160 350和160 367 bp。經序列對比分析,其序列變異信息見

表1 核桃葉綠體DNA擴增引物序列信息
表2。深紋核桃和核桃的葉綠體基因組間僅有14個差異位點,總變異序列長度為30 bp,變異率為0.019 %。其中,單核苷酸多態性位點數(SNP數)有7個,2個為堿基轉換,5個為堿基顛換;插入/缺失位點數(InDel數)有7個,6個為單堿基的InDel,1個為17堿基的InDel。分析結果表明,深紋核桃與核桃葉綠體基因組存在明顯的差異,但二者不存在大范圍的結構變異。
2.2 云新高原核桃與其親本的葉綠體基因組擴增片段多態性分析
根據深紋核桃與核桃的葉綠體基因組間存在的14個變異位點,分別對云新高原及其父本(云林A7)、母本(漾濞泡核桃)的14個葉綠體基因片段進行測序及分析。結果表明,云新高原與母本在14個變異穩點上,堿基完全一致,但二者均與深紋核桃葉綠體模式基因組序列(KX424843)在115 907位點存在1個SNP(G/A堿基轉換),變異之后與核桃葉綠體模式基因組序列(KT963008)保持一致;云新高原的父本與核桃葉綠體模式基因組序列(KT963008)存在2個SNP(132 545位點存在1個A/T堿基顛換,134 335位點存在1個T/C堿基轉換),而云新高原與其父本在所檢測的14個位點中,僅有11個位點存在差異。綜合表2可以看出,14個檢測位點中,云新高原與母本完全一致,而與父本存在11個位點的差異(占79 %),且差異位點僅有1個發生于基因編碼區(占9 %),其余10個位點均位于非編碼的基因間隔區(占91 %)。
另外,對9034(psbK-psbI基因間隔區)和118 575(ndhF-rpl32基因間隔區)變異位點檢測的過程中發現,這2個擴增片段還分別含有8和14個變異位點,具體信息見圖1。由圖1可以看出,在這22個變異位點中,云新高原與其母本在其中10個位點上具有相同的SNP或InDel,余下的12個位點與其父本的相同。
2.3 云新高原核桃與其親本的葉綠體基因單倍型差異分析
對云新高原核桃和其親本的11個基因或基因間隔區片段進行單倍型分析,共發現有11種單倍型(表3),其中10個單倍型均表現為母系遺傳,而剩余的ndhF-rpl32片段的單倍型較為復雜,存在7種亞單倍型,其中4種表現為母系遺傳,3種表現為父系遺傳,即云新高原核桃的ndhF-rpl32片段的單倍型表現為雙親遺傳。
表2深紋核桃、核桃、種間雜交種云新高原及其親本的葉綠體基因序列差異比較
Table 2 Comparisons of chloroplast DNA difference and polymorphism amongJ.sigillatacv. ‘Yangpao’,J.regia‘Yunlin A7’ and interspecific hybrid ‘Yunxingaoyuan’
變異類型Type變異位置Position深紋核桃J. sigillata核桃J. regia云新高原‘Yunxingaoyuan’漾濞泡核桃J. sigillata cv. ‘Yangpao’云林核桃J. regia ‘Yunlin A7’所在區域Region基因片段DNA nameSNP51 023TGTTGLSCrps4-trnTSNP56 262ATAATLSCndhC-trnVSNP115 907GAAAAIRaycf1SNP121 269ATAATSSCccsA-ndhDSNP130 197ACAACSSCycf1SNP132 545TATTTSSCycf1SNP134 335CTCCCIRbycf1InDel9 034-T--TLSCpsbK-psbIInDel35 041A-AA-LSCtrnE-trnTInDel35 914A-AA-LSCtrnM-psbDInDel64 845-T--TLSCpsaI-ycf4InDel118 412A-AA-SSCndhF-rpl32InDel118 575-A--ASSCndhF-rpl32InDel121 229~121 245-#1--#1SSCccsA-ndhD
注:“-”表示堿基缺失(InDel);“#1”表示序列TATTTTTAACTTAAGTT;“LSC”表示大單拷貝區域;“SSC”表示小單拷貝區域;“IR”表示反向重復序列。
Note:‘-’: Base deletion (indels); ‘#1’: TATTTTTAACTTAAGTT; ‘LSC’: Large single-copy; ‘SSC’: Small single-copy; ‘IR’: Inverted repeat.

*:堿基缺失*: single nucleotide deletion圖1 云新高原核桃及其親本與深紋核桃葉綠體模式基因組序列的2個片段的多態性比較Fig.1 Comparisons of two chloroplast DNA fragments polymorphism among ‘Yunxingaoyuan’, its parents and Juglans sigillata model sequence

編號No.基因片段DNA name變異位置Position單倍型 Haplotype漾濞泡核桃J. sigillata cv. ‘Yangpao’云林A7核桃J. regia ‘Yunlin A7’云新高原‘Yunxingaoyuan’1psbK-psbI9034A1A2A12psbI-trnS9256,9258,9279,9285,9312B1B2B13trnS9325,9329,9340C1C2C14trnE-trnT35 041D1D2D15trnM-psbD35 914E1E2E16rps4-trnT51 023F1F2F17ndhC-trnV56 262G1G2G18psaI-ycf464 845H1H2H19ndhF-rpl32118 412I1I2I1118 471I3I4I3118 473, 118 475, 118 481, 118 494, 118 503, 118 521, 118 533, 118 537I5I6I6118 575I7I8I7118 576, 118 594, 118 598I9I10I10118 600I11I12I11118 604I13I14I1410ccsA-ndhD121 229~121 245, 121 269J1J2J111ycf1130 197K1K2K1
注:粗體顯示云新高原葉綠體DNA的單倍型與其父本相同。
Note: Bold characters showed that the chloroplast haplotype of ‘Yunxingaoyuan’ was identical to that of its farther parent.
3 討 論
本項研究基于網絡數據庫中已公布的序列,從葉綠體全基因組層面分析,發現深紋核桃與核桃的葉綠體基因組存在14個變異位點、30個堿基的差異。總體上,93 %的差異類型(13/14)為單堿基多態性變異,其中7個為SNP,6個為單堿基InDel;7 %的差異類型(1/14)為17個堿基的短片段InDel,不存在大范圍的結構變異,這與水稻[13]、大麻[14]和其它植物[15]關于葉綠體基因組結構的研究相同。
針對云新高原核桃及其父母本的這14個變異位點進行多態性檢測,發現僅11個位點在三者間存在多態性,另外3個位點的堿基之間則完全相同,不具有多態性。這11個多態性位點中,云新高原與其母本完全一致,而與其父本完全不一致。并發現有1個位于基因編碼區,其余10個都位于非編碼的基因間隔區。在psbI-trnS和ndhF-rpl32兩個片段序列中,另發現存在22個變異位點,其中19個位點位于基因間隔區,云新高原與其母本在這22個位點中有10個保持一致,而與其父本在余下的12個位點保持一致。綜合而言,云新高原與其親本三者間共檢測發現33個變異位點,云新高原與其母本在其中21個位點上保持相同堿基,占全部變異位點數的64 %;云新高原與其父本在余下12個位點上保持相同堿基,占全部變異位點數的36 %。這33個變異位點分屬11個基因或基因間隔區片段,可以分為11種單倍型,云新高原的這11個單倍型中有10個與母本相同,1個為雙親的嵌合體。據此推斷,云新高原的葉綠體DNA為以母系遺傳為主的雙親遺傳。這與草本植物綠絨蒿(Meconopsis)相類似,其種間雜交種與其親本在trnL-trnF片段中存在25個變異位點,其中84 %的位點與母本一致,4 %的位點與父本一致,其余12 %的位點與親本均不相同[16]。另外,ndhF-rpl32DNA片段具有多個變異位點,是多態性高的葉綠體基因片段,有望廣泛應用于核桃群體遺傳分析、譜系地理和種內品種鑒定等研究。
大多數被子植物的葉綠體DNA通過雌配子以母系遺傳的方式傳遞[2-3],父系遺傳僅見于紫花苜蓿(Medicagosativa)[17]、甘草(Glycyrrhizaspp.)[18]和牽牛花(Pharbitisnil)[3]等少數種類中;裸子植物中父系遺傳較為普遍[3-4];天竺葵屬(Pelargonium)和月見草屬(Oenothera)植物是雙親遺傳方式的代表[3]。此外,隨著研究手段和研究材料的更新和豐富,之前認為獼猴桃(Actinidia)是嚴格的父系遺傳,但后來發現還具有母系及雙親遺傳的特性,與雜交親本的基因型有一定關系[19]。總之,植物葉綠體DNA的遺傳方式具有多樣性和復雜性。而胡桃科植物的葉綠體DNA遺傳方式的研究尚無相關報道,現有的相關葉綠體DNA研究都是針對系統進化和植物分類開展的。本項研究發現,核桃葉綠體DNA的遺傳方式為以母系遺傳為主的雙親遺傳,這為今后核桃種間雜交和遺傳變異研究提供了新的資料。
4 結 論
深紋核桃與核桃的葉綠體基因組存在14個變異位點、30個堿基的差異,其中7個為核苷酸多態性變異(SNP),7個為堿基插入/缺失變異(InDel)。對云新高原及其親本的這14個位點及其附近片段進行檢測,發現三者存在33個變異位點,云新高原與其母本在其中21個位點上保持相同堿基,占全部變異位點數的64 %;云新高原與其父本在余下12個位點上保持相同堿基,占全部變異位點數的36 %。這33個變異位點分屬11個基因或基因間隔區片段,可以分為11種單倍型,云新高原的這11個單倍型中有10個與母本相同,1個為雙親的嵌合體。根據質體遺傳的特性,推斷核桃葉綠體DNA的遺傳方式為以母系遺傳為主的雙親遺傳。另外,核桃ndhF-rpl32DNA片段存在多個變異位點,多態性高,該葉綠體基因片段可廣泛應用于核桃系統發育、群體遺傳和種內品種鑒定等研究。
