中藥青天葵黃酮醇合成酶基因的克隆與生物信息學分析
2018-11-30 06:46:40黃瓊林卓一南曾湘達詹若挺
西南農業(yè)學報 2018年10期
關鍵詞:植物
黃瓊林,卓一南,曾湘達,何 瑞*,詹若挺
(1.廣東醫(yī)科大學,廣東 湛江 524023;2.廣州中醫(yī)藥大學中藥資源科學與工程研究中心,廣東 廣州 510006;3.嶺南中藥資源教育部重點實驗室,廣東 廣州 510006)
【研究意義】嶺南道地藥材青天葵來源于蘭科多年生植物毛唇芋蘭Nerviliafordii(Hance) Schltr.,以葉片或全草入藥,具有清肺止咳,清熱解毒,散瘀消腫、鎮(zhèn)靜止痛等功效,在臨床上治療肺部疾病尤其是小兒呼吸道疾病的療效顯著[1]。青天葵萌發(fā)對環(huán)境要求苛刻,休眠期長(當年9月至次年4月),繁殖系數(shù)低,一顆球莖僅生長一片葉子,再加上人為的過度采挖,青天葵資源早已面臨匱乏甚至瀕危的困境,并在20世紀90年代被列入《中國南部石灰?guī)r稀有瀕危植物名錄》[2]。青天葵的優(yōu)質品種選育已成為當前青天葵資源可持續(xù)利用的重要課題。【前人研究進展】通過基因工程調控生物合成關鍵酶基因的表達來提高化學成分的積累被認為是植物或農作物品種選育和改善的有效途徑之一,并已具有較為成熟的基礎和廣泛的應用。前期的化學分析和藥理研究表明,黃酮類成分是青天葵的主要藥效物質[3-4]。因此,調控參與青天葵黃酮生物合成的關鍵基因從而促進青天葵有效化學成分的合成,是改善青天葵品種、提高青天葵藥用質量進而緩解青天葵資源嚴重短缺的可考慮途徑之一。黃酮醇合成酶(flavonol synthase, FLS)是連接類黃酮合成途徑和花青素合成途徑的重要分支點和關鍵酶,它以二氫黃酮醇為底物,催化黃酮結構中的C3位發(fā)生羥基化,從而衍生一系列黃酮醇類化合物[5-6]。FLS基因的過表達可以有效地提高黃酮醇類化合物的積累。如,Zhou 等將茶樹FLS基因在煙草中過量表達,轉基因煙草中的槲皮素、槲皮素-3-O-葡萄糖苷和槲皮素-7-O-葡萄糖苷等黃酮醇及其衍生物、總黃酮含量均顯著升高,而花青素的合成受到一定程度的抑制[7]。目前,F(xiàn)LS基因已從杜仲[8]、金銀花[9]、葡萄風信子[10]等植物中克隆出來。在前期的青天葵轉錄測序研究中,作者曾挖掘得至1條長度為840 bp的黃酮醇合成酶的基因片段[11]。【本研究切入點】采用RACE-PCR技術克隆青天葵FLS基因,命名為NfFLS,并采用生物信息學分析此基因的序列特征以及預測其編碼蛋白的和功能,【擬解決的關鍵問題】為NfFLS基因后續(xù)在青天葵植物中的功能鑒定并利用基因工程促進青天葵黃酮類成分的生物合成提供科學依據(jù)。
1 材料與方法
1.1 植物材料
青天葵植物樣品由廣西壯族自治區(qū)中藥研究所黃云峰副研究員采摘于廣西壯族自治區(qū)南寧市馬山縣,并鑒定為毛唇芋蘭Nerviliafordii(Hance) Schltr.,移栽至廣州中醫(yī)藥大學嶺南中藥資源教育部重點實驗室栽培。
1.2 試劑
RNAiso Plus (total RNA提取試劑)、PrimeScript?RT reagent Kit with gDNA Eraser (Prefect Real Time)、SMARTer?RACE 5’/3’Kit、dNTPs Mixture、DNA 聚合酶和markers均購自寶日醫(yī)生物技術公司;EZgeneTMGEL/PCR Extraction Kit購自美國Biomiga生物公司;pGEM-T克隆載體購自美國Promega生物公司;引物合成和測序由廣州華大基因公司完成。
1.3 青天葵總RNA提取及cDNA合成
取適量的青天葵新鮮葉片置于滅菌研缽中,加液氮快速研磨成粉末狀,按照RNAiso Plus試劑說明書提取總RNA,并采用瓊脂糖凝膠電泳檢測RNA完整性和純度。根據(jù)實驗目的,分別采用PrimeScript?RT reagent Kit with gDNA Eraser (Prefect Real Time)、SMARTer?RACE 5’/3’Kit制備用于NfFLS基因核心片段和編碼區(qū)、RACE擴增所需cDNA。
1.4 引物設計
如表1所示,根據(jù)在轉錄組測序研究中獲得的青天葵FLS基因片段的序列,設計本研究NfFLS基因克隆所需的引物。
1.5 NfFLS基因的核心片段擴增
PCR反應體系含10× ExTaqBuffer 10 μl,10 μM dNTPs Mixture 8 μl,10 μM FLSc-F 2.5 μl,10 μM FLSc-R 2.5 μl,5U/μl ExTaqDNA 聚合酶 1.2 μl,cDNA 4 μl,并用滅菌蒸餾水補充至100 μl。PCR擴增程序為:94 ℃預變性2 min;94 ℃變性30 s,61 ℃退火30 s,72 ℃延伸60 s,35個循環(huán);72 ℃延伸5 min。PCR產物經(jīng)瓊脂糖凝膠電泳檢測和切膠回收后送樣測序。
1.6 NfFLS基因的RCAE擴增
NfFLS基因的RACE擴增通過巢式PCR完成。5’端、3’端的第一輪RACE-PCR擴增分別以UPM Long/FLS5-1、FLS3-1/UPM Long為引物,反應體系和擴增程序均與核心片段的擴增相同。第二輪RACE-PCR擴增分別以相應的第一輪PCR產物為模板,以UPM short/FLS5-2、FLS3-2/UPM short為引物,反應體系與核心片段的擴增相同,擴增程序為:94 ℃預變性2 min;94 ℃變性30 s,66 ℃退火30 s,72 ℃延伸30 s,35個循環(huán);72 ℃延伸5 min。第二輪PCR產物經(jīng)瓊脂糖凝膠電泳檢測后切膠回收。RACE-PCR產物與pGEM-T載體連接后,熱擊轉化至E.coliTop10感受態(tài)細胞,采用載體通用引物對經(jīng)菌液PCR篩選的陽性重組子進行測序。
1.7 cDNA全序列的拼接及編碼區(qū)的克隆
采用DNAMAN軟件比對拼接前面所獲的NfFLS基因核心片段、5’端和3’端序列,得到該基因的cDNA全長序列。應用Omiga軟件在cDNA序列中搜索開放閱讀框(open reading fragment, ORF),并推導出氨基酸序列。將所獲的氨基酸序列在NCBI的BLASTP(http://blast.ncbi.nlm.nih.gov/)工具中進行同源檢索,檢驗其準確性和完整程度。
以FLSO-F/FLSO-R為引物擴增NfFLS基因的編碼區(qū),反應體系參考核心片段的擴增,但其中DNA聚合酶改用LATaq及其相應緩沖液替換ExTaq及其相應緩沖液進行反應,擴增程序為:94 ℃預變性3 min;94 ℃變性30 s,66 ℃退火30 s,72 ℃延伸60 s,35個循環(huán);72 ℃延伸7 min。PCR產物與pGEM-T載體連接后,熱擊轉化至E.coliTop10感受態(tài)細胞,采用載體通用引物對經(jīng)菌液PCR篩選的陽性重組子進行測序。

表1 用于NfFLS克隆的引物信息
注:帶*號引物為試劑盒自帶,其他為自行設計。
Note: The primers labeled with * were originated from the kit, and the others were designed by authors.
1.8 NfFLS及其編碼蛋白的序列分析和功能預測
使用ProtParam網(wǎng)站(http://web.expasy.org/protparam/)計算目的基因編碼蛋白的氨基酸組成、相對分子量和等電點等理化;采用ProtScale網(wǎng)站(http://web.expasy.org/protscale/)分析蛋白質的親/疏水性;通過SOPMA網(wǎng)站(http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_ sopma.html)預測編碼蛋白的二級結構;應用iPSORT網(wǎng)站(http://hc. ims.u-tokyo.ac.jp/iPSORT/)分析編碼蛋白的亞細胞定位。使用TMHMM(http://www.cbs.dtu.dk/services/TMHMM-2.0/)網(wǎng)站在線預測編碼蛋白的跨膜結構域。利用NCBI進行編碼蛋白的同源檢索和功能域分析,并通過DNAMAN軟件完成氨基酸序列的多重比對。
2 結果與分析
2.1 NfFLS基因的克隆和序列拼接
如圖1所示,經(jīng)PCR擴增出900 bp核心片段產物,通過巢式PCR分別擴增出約350和450 bp的5’端和3’端RACE-PCR產物。將上述PCR引物測得的序列進行拼接,獲得1268 bp的NfFLS基因cDNA全長序列,其中包含48 bp的5’端非編碼區(qū),993 bp的ORF(即編碼區(qū))和198 bp的3’端非編碼區(qū),最后以polyA序列結尾。
圖2所示,在獲得的cDNA全長序列的基礎上,設計用于擴增NfFLS基因編碼區(qū)的引物FLSO-F/FLSO-R并進行擴增,得到與前述ORF大小相符的編碼區(qū)產物。將該編碼區(qū)產物克隆至pGEM-T載體并篩選陽性重組子進行測序,獲得長度為993 bp的NfFLS編碼區(qū)序列,并將其翻譯成氨基酸序列。
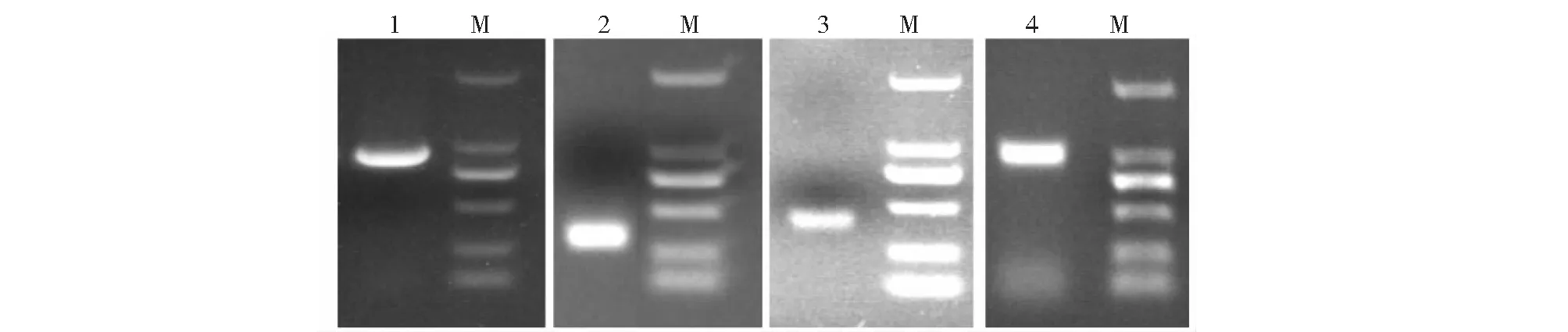
1.核心片段擴增產物;2.5’端RACE-PCR擴增產物;3.3’端RACE-PCR擴增產物;4.編碼區(qū)擴增產物;M.DL2000 marker,條帶依上至下的大小分別為2000、1000、750、500、200、100 bp圖1 NfFLS基因的PCR擴增Fig.1 Amplification of NfFLS gene
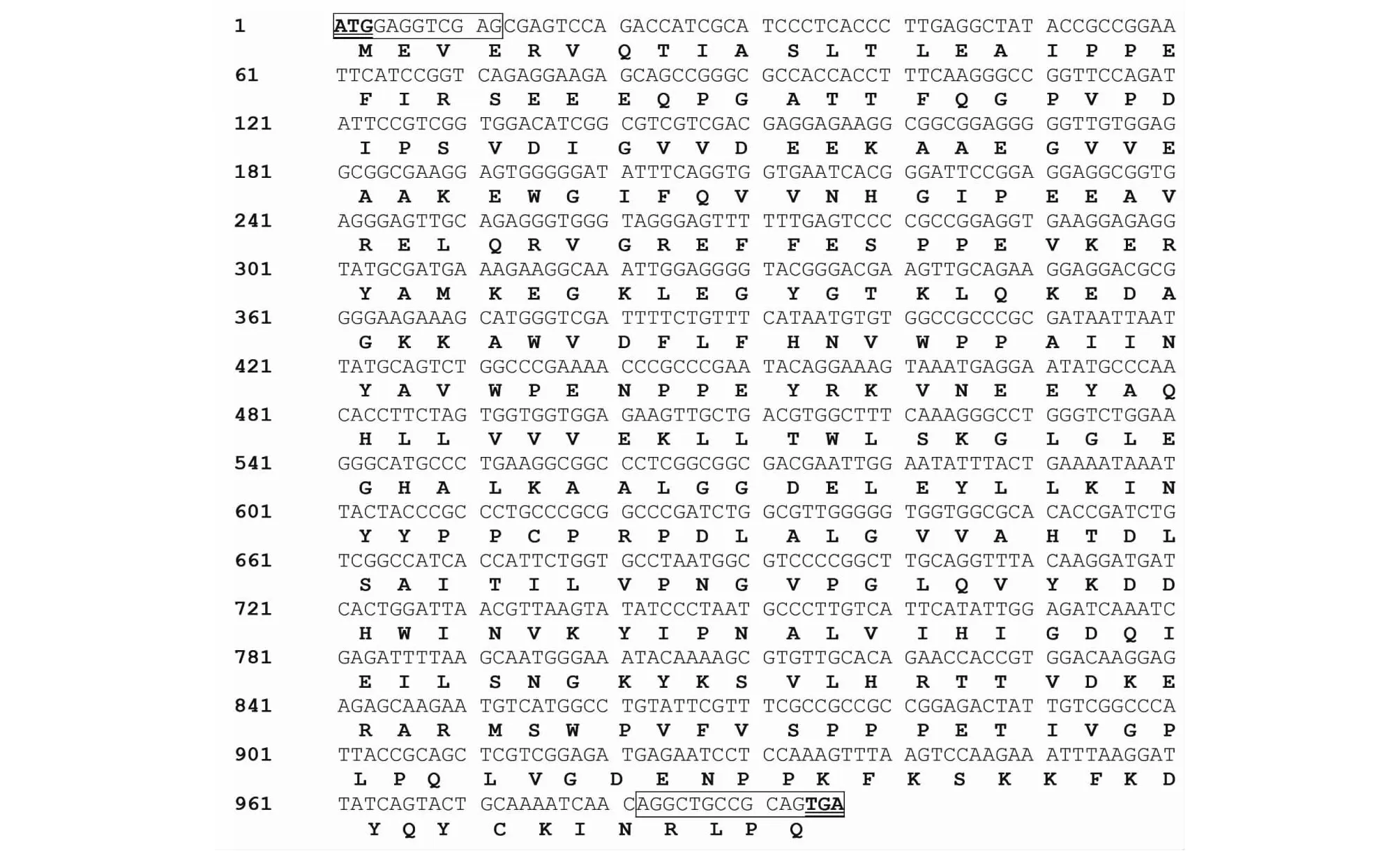
序列兩端的ATG和TGA分別為編碼區(qū)的啟動密碼子和終止密碼子以及方框所示分別為引物FLSO-F、FLSO-R部分序列圖2 NfFLS編碼區(qū)核苷酸和氨基酸序列Fig.2 Nucleotide and amino acid sequence of NfFLS gene
2.2 NfFLS核苷酸及其編碼氨基酸的理化性質分析
NfFLS編碼區(qū)GC含量為54.2 %。編碼蛋白含331個氨基酸,相對分子量為37.3 kDa。等電點為5.35,提示其屬于酸性蛋白質。在氨基酸構成上,谷氨酸(Glu)和纈氨酸(Val)所占比例最高,分別達到10.3 %和10.0 %。蛋白質不穩(wěn)定指數(shù)為35.56,小于40,說明其為穩(wěn)定蛋白質。
2.3 NfFLS蛋白的親/疏水性分析
NfFLS蛋白的親/疏水性分析發(fā)現(xiàn),NfFLS蛋白整體呈現(xiàn)為親水性,并且不含明顯的疏水區(qū)域,說明NfFLS為水溶性蛋白質。
2.4 NfFLS蛋白的保守功能域分析
NfFLS蛋白的保守功能域預測如圖3所示,該蛋白(第1~331個氨基酸)包含1個黃酮醇合成酶(flavonol synthase)功能域,說明它具有黃酮醇合成酶的功能。此外,它還包含1個2OG-FeII_Oxy功能域(第194~292個氨基酸)和1個PcbC功能域(第39~308個氨基酸),表明NfFLS蛋白為α-同戊二酸依賴性雙加氧酶(α-oxoglutarate-dependent dioxygenase)超家族和異青霉素N合成酶及相關雙加氧酶(Isopenicillin N synthase and related dioxygenases)超家族成員。
2.5 NfFLS蛋白的跨膜結構域預測以及信號肽、轉運肽分析
如圖4所示,NfFLS蛋白的整條多肽鏈均位于細胞膜外,不含有跨膜結構域。亞細胞定位分析顯示,NfFLS蛋白也不含信號肽、轉運肽和目標肽,提示其為非分泌性蛋白質。

圖3 NfFLS蛋白保守功能域Fig.3 Conserved domains of NfFLS protein
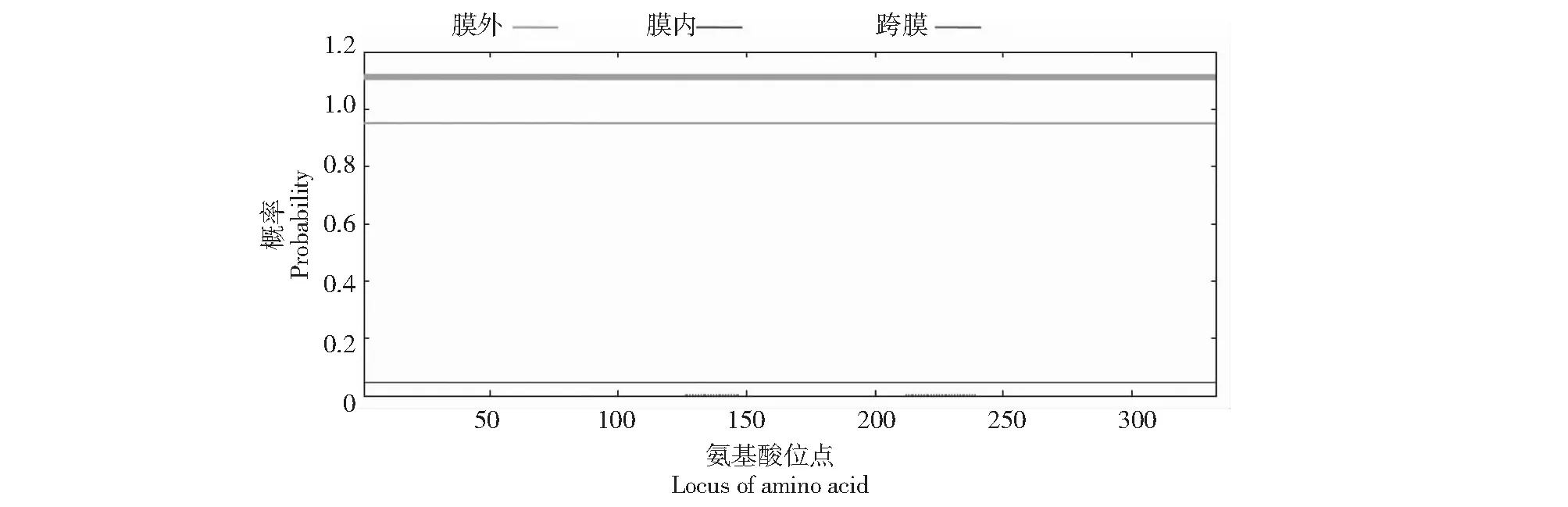
圖4 NfFLS蛋白的跨膜結構域Fig.4 Transmembrane structural domain of NfFLS protein
2.6 NfFLS蛋白的二級結構分析
如圖5(見封3)所示,NfFLS蛋白二級結構包含34.83 %的α-螺旋、36.04 %的隨意卷曲、18.02 %的延伸鏈和的β-轉角,表明NfFLS蛋白是由多種元件組成的混合結構型蛋白質。
2.7 NfFLS蛋白的同源比對和系統(tǒng)進化分析
如圖6(見封3)所示,NfFLS與同科的鐵皮石斛序列同源性最高,為82 %;與雙子葉植物淫羊藿的同源性較低,序列相似度也達65 %,說明FLS蛋白在不同植物中的保守性較高,且從圖中也可看見序列之間存在多處保守區(qū)域。
上述10種FLS蛋白的系統(tǒng)進化關系(圖7),NfFLS與同為蘭科植物來源的DcFLS、ChFLS首先聚在一起,然后再與單子葉植物來源的NtFLS和AcFLS形成一個分支,而其他雙子葉植物來源的FLS則聚集成另外一個分支。10種FLS蛋白的系統(tǒng)進化關系與其來源植物的進化關系相一致,較好地反映了植物FLS的自然進化關系。
3 討 論
目前,天然化學研究已經(jīng)從青天葵中分離出30余種類黃酮化合物,主要為黃酮醇、黃酮及其苷,這些化合物是青天葵發(fā)揮抗腫瘤、抗炎、抗氧化和防治心血管疾病等藥理活性的有效成分[12]。其中黃酮醇還參與抗紫外輻射、激素運輸、花色性狀等生理過程[13]。FLS作為類黃酮生物合成途徑和調控黃酮醇化合物生物合成的關鍵酶,其編碼基因的克隆和蛋白表達對青天葵黃酮類藥效成分的積累以及品種優(yōu)育具有重要的意義。轉錄組分析被認為是快速挖掘物種功能基因的高通量手段,尤其是無基因組參考物種[14-15]。本研究基于轉錄組測序挖掘到的青天葵FLS基因片段,設計高特異性引物和巢式PCR技術,經(jīng)過一次實驗就能得到NfFLS的核心片段并進而擴增出基因cDNA全長。相比傳統(tǒng)的基因克隆方式,設計簡并引物難度較大并需要多次PCR操作才能擴增出目的基因的核心片段,本研究通過轉錄組測序挖掘青天葵的功能基因具有較好的優(yōu)勢。本研究應用生物信息學方法分析發(fā)現(xiàn),NfFLS基因編碼包含331個氨基酸、分子量為35.3 KDa、不含有跨膜結構域和信號肽、定位于細胞質中的穩(wěn)定性親水性蛋白,NfFLS還具有植物黃酮醇所共享的保守結構域,并于其他植物來源的FLS蛋白具有較高的同源性。這些結論證實了NfFLS基因是青天葵黃酮醇合成酶的編碼基因,并為后續(xù)青天葵FLS基因及其編碼蛋白的功能鑒定和表達調控提供了理論依據(jù)。筆者曾對通過轉錄組測序獲得的黃酮醇合成酶基因片段進行表達分析,采用RPKM(Read Per Kb per Million reads)[16]方法探討該基因在青天葵葉片和球莖中的表達量差異。結果顯示,該黃酮醇合成酶基因在葉片中的表達量高于在球莖中的表達量,提示黃酮醇合成酶基因傾向于在青天葵地上部分表達和發(fā)揮調控功能,這一組織表達特性也與煙草[17]、紫山藥[18]等植物的FLS基因表達模式相同。由于本研究在此黃酮醇合成酶基因片段的基礎上進行基因克隆,克隆得到NfFLS基因也完全包含該片段,再加上青天葵植物材料的短缺,因此,本研究沒能夠采用熒光定量PCR分析NfFLS在青天葵組織器官中的表達差異。
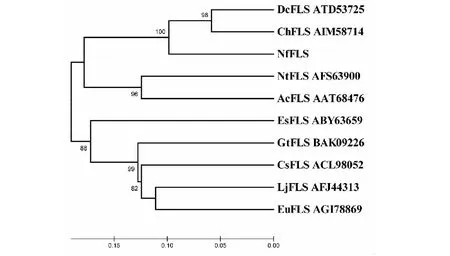
圖7 10種植物FLS蛋白的系統(tǒng)進化樹Fig.7 Phylogenetic tree of FLS proteins from 10 species
4 結 論
本研究成功克隆青天葵的類黃酮生物合成途徑關鍵酶基因NfFLS,該基因編碼區(qū)長度為993 bp,編碼331個氨基酸,其編碼蛋白具有黃酮醇合成酶保守功能域、與鐵皮石斛等蘭科植物來源FLS蛋白的同源性可達82 %。本研究為利用FLS基因調控青天葵黃酮類藥效成分積累,從而改良青天葵的種質,促進瀕危藥材青天葵的可持續(xù)性利用奠定了基礎。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數(shù)英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數(shù)英綜合(2020年11期)2020-12-16 02:57:22
學苑創(chuàng)造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
