利用CRISPR/Cas9技術定點敲除煙草多酚氧化酶基因NtPPO1
2018-11-30 03:08:54姚恒楊大海白戈謝賀
生物技術通報 2018年11期
姚恒 楊大海 白戈 謝賀
(云南省煙草農業科學研究院,昆明 650021)
在植物中,多酚氧化酶(Polyphenol oxidase,PPO)能將酚類物質催化反應出能直接殺滅病原菌的醌類物質,被認為對植物抗蟲、病菌等生物因素性脅迫是有益的。然而,因為PPO催化產生深褐色的醌類物質又直接危害果蔬、水稻及小麥等農產品的外觀品質,因此,過強的PPO酶活性對作物的品質有害。植物中的PPO的功能具有多樣性,在豆類[1]、番茄[2-3]、馬鈴薯[4]、紅三葉草[5-6]、蘋果[7]、水稻[8]和小麥[9-10]等植物中已有大量的報道。育種學家在研究PPO基因功能的基礎上,根據需求選育出PPO酶活性強或者弱的品種,創制出抗病蟲害或者不易褐化的產品。煙草也存在抗病蟲害與低褐變煙葉品種的需求,利用現代分子標記手段揭示煙草品種耐烤性的分子基礎,提出決定煙草品種耐烤性的候選基因[11-12]。但煙草PPO基因的功能研究主要集中在煙草調制過程中褐變與PPO酶活力相關性的研究[13-16]。針對煙草PPO基因功能的研究相對較少[17-23]。研究煙草PPO基因功能,需要創制突變體材料。CRISPR/Cas9技術于2015年成功應用于煙草[24]基因功能研究,目前,國內關于煙草中CRISPR/Cas9技術的應用正在普及。為了獲得NtPPO1的定點突變體,本研究利用CRISPR/Cas9技術構建煙草多酚氧化酶NtPPO1的編輯載體,篩選NtPPO1被定點編輯敲除的純合突變體。通過實時定量PCR技術檢測到突變體中NtPPO1的相對表達量,突變體的創制為研究其基因功能奠定了材料基礎。
1 材料與方法
1.1 材料
栽 培 煙 草 品 種(Nicotiana tabacum)K326、DH5α大腸桿菌與LBA4404農桿菌感受態為云南省煙草農業科學研究院實驗室保存。pORE-CRISPR/Cas9植物表達載體由西南大學惠贈。引物合成和測序均由上海英濰捷基貿易有限公司(Invitrogen)完成。
1.2 方法
1.2.1 引物設計和基因克隆 以煙草葉片cDNA為模板,根據NtPPO1序列設計引物(F:5'-ATGGC TTCTTCATTTGTTCTTCAAGC-3';R :5'-TTAACAA GGGACCAACTGGATCTCAAC-3'),利用 Q5 超保真DNA聚合酶進行擴增,擴增產物經過瓊脂糖凝膠電泳進行檢測。回收、純化、連接和轉化,經抗性篩選,PCR鑒定后挑選陽性單克隆菌落測序驗證。
1.2.2 sgRNA表達載體的構建 根據測序正確的NtPPO1序列,設計靶位點sgRNA序列:5'-CATTTGTTCTTCAAGCTCCA-TGG-3'作 為CRISPR/Cas9敲除NtPPO1的靶位點序列。
根據pORE-CRISPR/Cas9植物表達載體構建方法,需要在sgRNA靶序列添加核苷酸G與BsaⅠ的酶切接頭(F:5'-GATTGCATTTGTTCTTCAAGCTCCA-3';R :5'-AAACTGGAGCTTGAAGAACAAAT GC-3')。sgRNA退火反應:在PCR儀器中設定程序每8 s降低0.1℃,將混合單鏈引物從95℃降至4℃以形成引物DNA雙鏈。
利用BsaⅠ酶切pORE-CRISPR/Cas9植物表達載體,將其與已退火好的靶位點DNA引物進行連接,構建成NtPPO1的CRISPR/Cas9載體。將目的載體直接轉化大腸桿菌DH5α感受態細胞,利用含卡那霉素的抗性培養基篩選陽性克隆。利用上游引物:5'-TTAGGTTTACCCGCCAATA-3', 與 sgRNA 反向引物配合PCR篩選陽性菌落。
1.2.3 農桿菌轉化載體構建與煙草遺傳轉化 提取克隆載體中的質粒轉化農桿菌LBA4404。利用含載體農桿菌浸染煙草葉盤8-10 min。然后經過2 d共培養,轉移至含有NAA、6-BA和卡那霉素(50mg/L)以及頭孢噻肟鈉(500 mg/L)的MS固體培養基中進行組培再生,獲得轉基因煙草植株。
1.2.4 煙草NtPPO1突變體的篩選 提取轉基因煙草植株DNA。利用Kana引物(F:5'-CAGGTTCTCC G G C C G C T T G G-3',R :5'-GGAGATCCTGCCCCGG-CACT-3') 進 行PCR,檢測陽性植株。利用Cas9引物(F:5'-TGTCGGGACAGGGCGACAGT-3',R:5'-CGCCGCGTTCAGCCTTTGTG-3')進行 PCR,檢測陽性植株。
挑取經PCR檢測陽性的煙草小苗,設計包含靶位點區域的長度為770 bp的片段引物(F:5'-TTCACGCACTAATTCGTCTCATTGGA-3',R:5'-TCTGTCGGCAACGCCTTCA-3')進行檢測。
PCR擴增NtPPO1片段,擴增產物經測序、比對后,分析轉基因后代植株中NtPPO1在靶位點處核苷酸序列變化。
基因編輯成功的轉基因陽性T1植株,分單株套袋自交收獲T2種子。播種萌發T2種子獲得T2小苗100株,提取T2植株DNA,利用上述方法檢測NtPPO1突變情況,并分析T2純合突變煙草中NtPPO1序列在靶位點處突變類型。
1.2.5 煙草NtPPO1的Real time-PCR檢測 按照Quant qRT-PCR kit(SYBR Green)(TIANGEN 公司)使用說明在QuantStudioTM6 Flex熒光定量PCR儀器(ABI公司)上進行qRT-PCR檢測。3個生物學重復,3個技術重復,利用Excel進行統計分析,以sigmplot軟件繪制qRT-PCR分析圖。分析NtPPO1的相對表達量,內參基因為26S。所需引物為NtPPO1_qRT_F:5'-AGCACTGCCTTTATTGATGATGG-3',NtPPO1_qRT_R:5'-ACTTAGCTATGTATTCGTCATCTACAGTT-3'。26S_F:5'-GAAGAAGGTC C C A A G G G T T C-3',2 6 S_R :5'-TCTCCCTTTAACAC-CAACGG-3'。
2 結果
2.1 NtPPO1全長cDNA序列驗證
如圖1所示,利用設計的克隆引物以煙草cDNA為模板,特異性擴增出一條約1.8 kb的片段,經測序比對分析NtPPO1的基因座(圖2-A),NtPPO1編碼序列為1 746 bp,其外顯子有3個,其中第2外顯子可以選擇性拼接。測序發現5個克隆中有4個克隆的NtPPO1編碼序列為1、2、3外顯子拼接而成(圖2-B),另外1個克隆的NtPPO1編碼序列為1、3外顯子拼接而成(圖2-C)。NtPPO1可變剪切形式為外顯子跳過,在不改變起始位點的情況下,NtPPO1轉錄后的mRNA會提前出現終止密碼子。跳過的第2外顯子序列為5'-GTACTTATA CTTCCATGAGAGAATCTTGGCTTCCCTAATTGG TGACCCTACATTTGCTTTGCCATATTGGAATTGG GACCATCCTAAAGGCATGCATTTGCCTGACATG TTTGATGTCGAAG-3'。
在野生型的煙草植株中,NtPPO1就存在2種不同的轉錄形式,雖然轉錄本會提前出現終止子(圖2-C),但是煙草中保存這種轉錄剪切方式應該有其重要的生理功能,推測這種無義介導的mRNA降解(Nonsense-mediated mRNAdecay,NMD)可以監控細胞中的mRNA水平以便煙草能夠快速的響應外界刺激。

圖1 NtPPO1的cDNA擴增產物
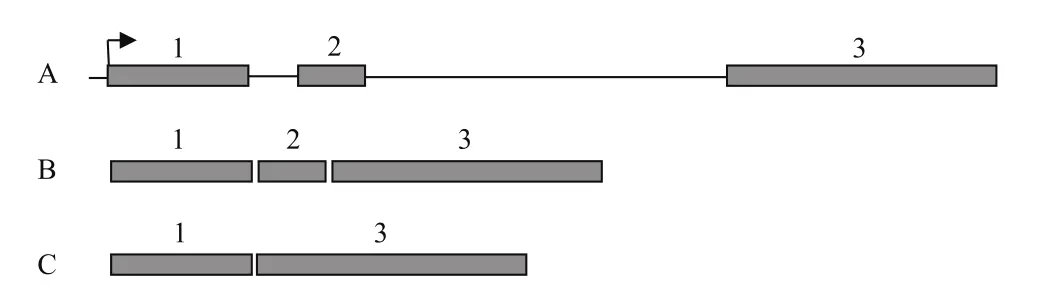
圖2 NtPPO1基因座圖
2.2 NtPPO1突變靶位點的選擇與載體構建
NtPPO1的CDS序列經過測序驗證后,選取翻譯起始密碼子ATG后第11-30的20 bp核苷酸序列作為敲除基因的靶位點序列,構建CRISPR/Cas9植物表達載體,目標載體轉入農桿菌后,圖3顯示,通過測序驗證篩選出含sgRNA序列正確插入到CRISPR/Cas9表達載體的農桿菌菌株。
含正確插入sgRNA的基因編輯載體,利用農桿菌葉盤法轉化煙草,經過組培再生成轉基因T1代植株。
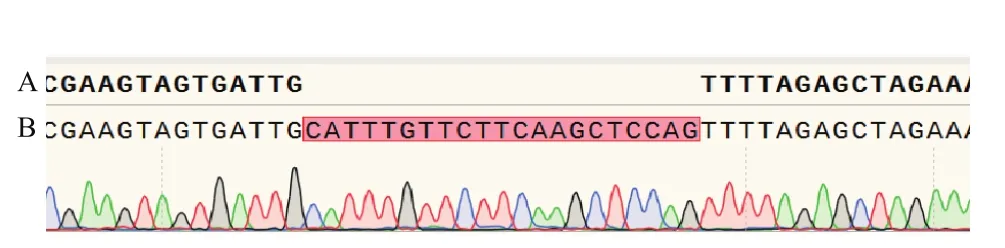
圖3 pORE-CRISPR/Cas9∷sgRNA基因編輯載體
2.3 NtPPO1敲除陽性T1植株篩選
通過測序方法篩選NtPPO1被編輯的轉基因植株T1(圖4),編輯成功的T1植株在sgRNA-PAM位點后序列經測序出現雙峰,并且測序峰圖紊亂的情況明顯不一致,說明Cas9蛋白在切斷目標序列后,DNA修復的情況有差異。將此三類轉基因T1雜合突變植株盆栽后自交收種。經過自交分離后產生的T2代植株中,會得到三類NtPPO1純合突變的T2植株。

圖4 NtPPO1定點敲除轉基因T1植株篩選
2.4 NtPPO1突變體的檢測
經測序分析T2植株NtPPO1在目標敲除序列后(圖5),突變體M1植株的NtPPO1缺失了1個G堿基,M2植株的NtPPO1缺失了2個G堿基,M3植株的NtPPO1突變是插入了1個A堿基,此3類突變都會造成NtPPO1編碼序列的移碼突變,如果轉錄出mRNA,在起始閱讀框不變的情況下會提前出現終止密碼子。

圖5 T2代轉基因植株NtPPO1的突變類型

圖6 T2轉基因植株中NtPPO1的相對表達量分析
2.5 突變體內NtPPO1相對表達量的檢測
測定NtPPO1純合突變植株M1、M2、M3的基因表達量如圖6所示,相對于野生型對照,M1、M2、M3突變體NtPPO1的表達量分別下降了75%、84%和76%。結果表明,利用CRISPR/Cas9技術創制的NtPPO1的突變體也影響了此基因的轉錄。在DNA水平上的點突變,造成了基因轉錄水平的改變。NtPPO1的突變是位于第1外顯子區域,而不是位于啟動子區域。理論上,造成NtPPO1表達量降低就應該發生在轉錄后的水平調控,具體是發生在轉錄后的哪個時期還有待進一步的研究。

圖7 NtPPO1純合突變體植株表型
2.6 轉基因煙株表型
如圖7所示,利用CRISPR/Cas9技術創制的NtPPO1突變體與對照野生型煙草之間的田間外觀表型無明顯差異。
3 討論
本研究選取了一個煙草多酚氧化酶NtPPO1進行基因序列克隆驗證,結果表明,NtPPO1存在一種可變剪切形式。在驗證了NtPPO1序列的基礎上,采用CRISPR/Cas9技術,創制1、2 bp堿基缺失和1 bp堿基插入的NtPPO1定點純合突變體M1、M2和M3。通過基因表達發現,突變體中NtPPO1表達水平有顯著的下降。
植物中的PPO功能具有多樣性。根據前人的研究結果,PPO與植物抗蟲、抗細菌性病害的能力相關。但是,在一些作物中通過降低作物中的PPO酶活,會減輕農作物發生褐變的程度,如在葡萄[25]、小麥[26]、水稻[27]等作物中的結果顯示,低的PPO活力能不同程度的顯示出不易褐變、耐貯藏的特性。根據本實驗基因測序的結果顯示,NtPPO1已經被成功敲除,并且基因的表達量也發生了顯著的下降。王曼玲等[28]報道植物的PPO一般存在多個基因編碼,而本研究普通栽培煙草中的PPO基因至少有15個(研究未發表),單獨敲除某一個PPO基因,可能會由于另外一個PPO基因在功能上互補,造成植株整體上的PPO活力下降并不顯著。所以需要研究煙草中的PPO基因的功能,還需要進一步的創制出各PPO基因的突變體,甚至是煙草PPO雙、三基因突變體,才能更全面的研究PPO基因功能。
根據qPCR測定結果顯示CRISPR/Cas9創制的NtPPO1的純合突變體其NtPPO1的表達水平顯著降低。說明利用CRISPR/Cas9在煙草中創制某個基因的定點突變時,也可能會影響此基因轉錄。這個結果和Cai等[29]在大豆中的研究結果類似,Cai等在大豆中靶向敲除控制開花的基因GmFT2a,在長日照和短日照的條件下,GmFT2a的相對表達量下降。根據Wu等[30]的研究表明,真核生物中,如果基因序列存在早期終止子(Premature termination codons,PTCs),相對于沒有PTCs的mRNA,攜帶PTCs的mRNA會通過無義mRNA降解途徑(Nonsensemediated mRNA decay pathway)快速降解,防止因翻譯出無意蛋白的積累才啟動降解蛋白調控。本研究利用CRISPR/Cas9在NtPPO1靠近起始密碼子的核苷酸序列上引入了移碼突變,結果提前出現了終止密碼子,可能啟動了煙草本身的NMD調控途徑,進而造成了NtPPO1的轉錄下降。當然,這只是一個推論。本研究利用CRISPR/Cas9基因編輯技術創制NtPPO1突變體,其NtPPO1表達水平發生下降的機理還有待于進一步的研究。
本研究利用CRISPR/Cas9基因組編輯技術獲得了煙草PPO基因NtPPO1的編輯敲除突變體,對NtPPO1的功能進行了初步研究,為煙草PPO基因功能研究和煙草育種奠定了材料基礎。
4 結論
本研究克隆驗證了煙草多酚氧化酶NtPPO1編碼序列,并利用CRISPR/Cas9技術成功創制了3個NtPPO1定點突變體材料,NtPPO1在突變體中的表達量顯著降低,而突變材料與對照煙草表型上無顯著差異。
猜你喜歡
奧秘(創新大賽)(2023年3期)2023-05-06 01:48:20
中等數學(2022年2期)2022-06-05 07:10:50
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
小學生學習指導(低年級)(2020年6期)2020-07-25 02:31:36
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
小學生學習指導(低年級)(2018年9期)2018-09-26 05:59:44
瘋狂英語·新讀寫(2018年2期)2018-09-07 09:32:10
浙江中西醫結合雜志(2017年2期)2017-01-12 18:23:59
