南海水域生物固氮作用研究進展
2018-12-06 03:37:18李丹陽
廈門大學學報(自然科學版) 2018年6期
李丹陽,張 潤
(廈門大學海洋與地球學院,福建 廈門 361102)
氮是海洋生物所必需的營養元素.海水中氮最大的儲庫是氮氣(N2),但海洋中絕大部分的初級生產者不能直接利用海水中的氮氣,因此,開闊大洋的生物生產力常受到氮營養鹽的限制[1-2].少數固氮生物可以在固氮酶的作用下,將N2還原成氨氮或者有機氮,這一過程被稱為生物固氮作用.通過固氮作用,固氮生物既可以滿足自身的氮需求,又可將新固定的氮傳遞給其他生物,緩解其受到的氮限制,支持海洋新生產力[1,3-4].因此,生物固氮作用是重要的新氮來源之一,影響著海洋對大氣中二氧化碳的吸收能力,進而對全球氣候產生重要的影響[1,3-4].
Dugdale等[5]利用15N2示蹤培養法,首次實測了馬尾藻海中束毛藻(Trichodesmium)的固氮速率.隨后,在一些低緯度寡營養鹽海區陸續開展了生物固氮作用的研究,采樣區域主要集中于亞熱帶北太平洋環流區(夏威夷HOTS站)及北大西洋等少數海區[3,6-7].隨著認識的不斷更新,海洋固氮研究已不再局限于低緯度寡營養鹽開闊大洋,而是拓展至更大的空間范圍,對于固氮生物組成的多樣性、復雜性也有了全新的認識[8-11].了解海洋固氮作用的時空分布,對全球海洋氮收支的評估及海洋氮循環的研究具有重要意義.
南海位于熱帶-亞熱帶地區,是世界第二大邊緣海.南海總面積約為3.5×106km2,平均水深1 300 m[12].南海大部分海區具有水體溫暖、寡營養鹽、距離大氣塵降源地較近等特點,這些環境特點被認為有利于固氮作用的發生[3].自20世紀末以來,陸續開展了南海固氮生物(主要針對束毛藻)豐度、固氮速率的時空分布、固氮作用的受控因子等方面的研究[2,13-14],極大地拓展了對南海生物固氮作用的認識.
本文中主要回顧近年來南海水域生物固氮作用的研究成果,并結合全球范圍內海洋固氮作用研究的最新認識,對南海固氮作用的研究提出展望.
1 南海不同區域的生物固氮作用
南海幅員遼闊,可根據理化條件的差異分為幾個亞區.南海北部受黑潮水入侵的影響[2,15-16],而沿岸區域又受珠江、湄公河等淡水徑流的影響[17-18].除此之外,在季風的影響下,南海常出現上升流[11,19-20],上升流為南海表層水體輸送大量的營養物質,影響南海表層營養鹽分布[21].其他海區的研究表明,生物固氮作用的調控機制往往具有很大的時空可變性[7].鑒于此,可將南海生物固氮作用的研究區域分為南海海盆區、南海北部陸架區、沿岸上升流區和半封閉海灣4個部分,近年來生物固氮作用研究站位分布情況如圖1所示.
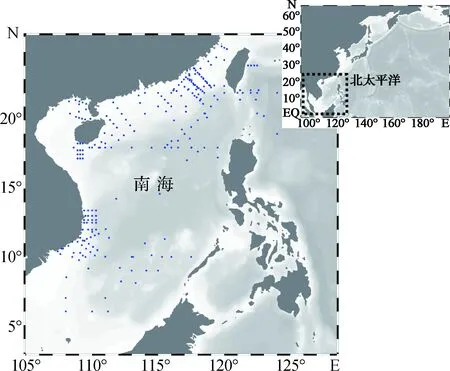
圖1 近年來南海生物固氮作用研究站位分布圖
1.1 南海海盆區
南海海盆區面積遼闊,約占南海總面積的1/2.目前,南海海盆區固氮作用的研究區域以北部海盆為主,特別集中于東南亞時間序列站(South East Asia time-series study station,SEATS站;位于116° E,18° N)及其鄰近海區.
早期,Wong等[22]通過計算SEATS站的氮異常指數,指出SEATS站可能存在較強的生物固氮作用.氮異常指數是指在反硝化作用和固氮作用的影響下,海水中氮營養鹽的過剩量,常被用來指征固氮作用的強度[23].近期,Kao等[24]在SEATS站發現了較高的氮異常指數,根據穩定同位素質量平衡模型計算,指出SEATS站的生物固氮作用引入的氮總量約為(20±26) mmol/(m2·a),占新氮總量的5%~10%.SEATS站可觀測到束毛藻和胞內植生藻(Richeliaintracellularis)[21],二者在春季的平均固氮量均可達到40 μmol/(m2·d).南海生物可利用的新氮來源主要包括大氣沉降、水體輸送和生物固氮三方面.南海海盆區固氮生物組成復雜[25-26],若僅考慮束毛藻和胞內植生藻的固氮作用,SEATS站中3種氮來源所提供的氮總量所支持的生產力僅占該區域群落凈生產力(4.47 mmol/(m2·d))的55%[27].這一方面揭示了再生氮對SEATS站群落凈生產力的支持,另一方面揭示了其他類型固氮生物的重要貢獻.若以SEATS站的生物固氮量為參考,將此調查結果外推到整個南海海盆(總面積≈1.8×106km2),南海海盆每年通過生物固氮作用收入的氮總量可達3.6×107mol.
盡管束毛藻長期被認為是全球海洋最重要的固氮生物[3],但在南海很少觀察到束毛藻的高豐度.近年來,單細胞固氮生物被發現廣泛分布于全球海洋中[9-10,28],在南海也可檢測到單細胞固氮生物的高豐度.Zhang等[25]于2011年報道了南海海盆區固氮生物的組成及豐度,指出南海海盆區的固氮生物以變形菌為主,其中α-變形菌和γ-變形菌的豐度最高(約為104~105L-1),相比之下,絲狀藍藻(如束毛藻)的豐度較低.單細胞固氮生物在南海廣泛分布,而不僅局限于海盆區[15-16,26],對南海的生物固氮作用可能有重要貢獻.
1.2 南海北部陸架區
南海北部廣闊的陸架區也是生物固氮作用研究的重要場所.黑潮入侵、陸地徑流等因素都可能對此處的固氮作用產生影響.
黑潮水自呂宋海峽進入南海,是影響南海北部陸架區固氮作用的一個重要因素.雖然具體原因不明,但是黑潮上游往往具有高固氮生物豐度[15,29-30].據報道,黑潮水中無論是束毛藻的平均豐度,還是水體實測固氮速率均比鄰近南海海區高出近1個數量級[15,31],隨著黑潮水的入侵,一定豐度的固氮生物也可能被攜帶進入南海.近年來的研究結果進一步證明,黑潮上游區的固氮生物群落也具有較高的多樣性,單細胞固氮生物是不可忽視的固氮貢獻者[15].Wu等[16]在2018年的報道中指出了黑潮及南海中單細胞固氮生物的重要性,報道指出:單細胞固氮生物的固氮速率變化范圍為0.63~7.61 nmol/(L·d),均值可達3.67 nmol/(L·d),單細胞固氮生物貢獻了采樣區域內近75%的固氮通量.
臺灣海峽南部也是固氮作用較為活躍的海區,林峰[32]利用15N2示蹤法測定了臺灣海峽南部海域的生物固氮速率,結果表明夏季臺灣海峽南部海域的生物固氮速率介于0.168~1.08 nmol/(L·d)之間,平均為0.537 nmol/(L·d),其中次表層的固氮速率更高.Chen等[15]指出,次表層較強的生物固氮作用是由單細胞固氮生物所貢獻.
受陸地徑流的影響,珠江口附近海域結合態氮營養鹽較為豐富,此處也是生物固氮作用的一個熱點區域.劉甲星等[33]報道了2014年秋季珠江口附近海域具有較高的固氮速率,表層水的固氮速率均值高達(0.77±0.54) nmol/(L·d).較高的無機氮營養鹽濃度并未抑制固氮作用的發生,這和該區域的固氮生物組成有著較大的關系.Kong等[18]發現珠江口附近海域固氮生物組成較為多樣,包括異養型變形菌、光合藍藻、Cluster Ⅲ類固氮生物等,而束毛藻所占比例不高,鹽度可能在一定程度上調控著固氮生物的組成.
1.3 沿岸上升流區
沿岸上升流是南海西側的一個典型現象.沿岸上升流區的固氮作用在很長時間內被忽視了,這是由于上升流水體具有的典型理化條件(較低的水溫、較為充足的結合態氮營養鹽等)與固氮作用的傳統觀點不符.但是,近年來越來越多的實測研究證據表明,沿岸上升流區并非固氮作用的禁區;相反地,熱帶-亞熱帶沿岸上升流區(包括赤道上升流區)往往是固氮作用發生的熱點,已有研究報道了南海越南沿岸上升流區、瓊東上升流區及臺灣海峽南部上升流區的固氮作用[11,19-20].
目前,在熱帶越南沿岸上升流區域開展的生物固氮作用的研究較多.在非季風時期,越南沿岸附近以寡營養鹽為特征,而在季風期,受上升流影響,越南沿岸的營養鹽含量明顯提升,除上升流外,湄公河徑流也是越南沿岸海域中重要的營養物質來源[34-36].湄公河河水每年向南海輸入大量淡水,使越南沿岸海域出現明顯的鹽度梯度,同時攜帶大量陸源的顆粒物質及微量元素進入南海[17,36].在越南沿岸上升流區,西南季風盛行期固氮速率較非季風期明顯升高,這表明湄公河羽狀流可能在很大程度上調控著該區域固氮速率及固氮生物的空間分布[11,37].目前具體作用機制不明,推測可能的原因包括河流淡水對水體垂向穩定度的提升、痕量元素的輸入、硅營養鹽輸入促進硅藻-固氮藻共生體的生長等方面[11,17-18].
Moisander等[13]檢測了越南沿岸流域中固氮生物的組成和豐度,結果表明:調查區域內的固氮生物主要由束毛藻和α-變形菌組成,除此之外,單細胞的固氮藍藻和共生型藍藻也廣泛存在.Grosse等[17]研究發現單細胞固氮生物多出現在湄公河河口附近,束毛藻僅在鹽度較高的(鹽度>33.5)區域中出現,而與硅藻共生的固氮生物則是廣泛分布于調查區域內.由此可見,湄公河徑流影響著越南沿岸流域中固氮生物的分布.Voss等[11]檢測了越南沿岸上升流區域內的生物固氮速率,結果表明:當西南季風盛行時,調查區域內的固氮速率變化范圍為21.4~190.6 μmol/(m2·d);非季風期時,固氮速率的變化范圍為1.9~36.0 μmol/(m2·d).相比之下,季風盛行時,固氮作用要明顯強于非季風期,這可能是由于湄公河徑流的影響造成的[11,38].來自湄公河的淡水一方面可以加強水體的穩定度,另一方面可攜帶大量微量元素進入南海,從而促進固氮作用[39].除微量元素外,湄公河向南海輸送了大量的硅,這也可能通過促進硅藻的生長而對共生型固氮生物產生間接的影響[11,40].因此,越南沿岸上升流區域內的固氮作用可能受上升流和湄公河徑流的共同影響.整體而言,在越南沿岸上升流區,絲狀藍藻和單細胞藍藻對固氮的貢獻量相當,固氮作用對初級生產力的貢獻率的變化范圍為1%~47%[17],是不容忽視的氮來源.
瓊東上升流是亞熱帶南海北部一個典型的沿岸上升流區.Zhang等[20]報道了2013年夏季海南島東側上升流區域的生物固氮作用,結果表明:該區域表層固氮速率的變化范圍為0.1~5.6 nmol/(L·d)(均值為1.0 nmol/(L·d)),固氮速率從沿岸區域到開闊海域有增強的趨勢,其對初級生產力的貢獻率變化范圍為0.01%~2.52%.若將瓊東上升流區的調查結果外推到南海西北陸架區(總面積≈33×104km2,17° N以北),假設夏季時長為90 d,生物固氮作用引入氮的總量約為1.4 ×109mol[20].該報道提出了一個固氮作用受控的物理-生物耦合機制,即:上升流向表層輸送低溫、富營養鹽且鐵含量豐富的深層海水,促進固氮生物增長,而非固氮生物對磷和鐵的吸收可能會抑制生物固氮作用,因此,隨離岸距離的增加,初級生產力減小,生物固氮作用增強[20],這一機制對于亞熱帶沿岸上升流區的固氮作用調控可能具有普遍性.Wen等[19]最新的調查研究中報道了臺灣海峽附近上升流區域的固氮作用,結果表明:調查區域內固氮作用強烈,最大固氮速率可達7.51 nmol/(L·d),在氮磷摩爾比較低的區域中生物固氮速率更高.
1.4 半封閉海灣
南海存在諸多半封閉海灣,如北部灣、大亞灣等,這些海區往往具有迥異于開闊水體的特點,也是潛在的固氮熱點區域.Dong等[41]研究發現鐵氏束毛藻(Trichodesmiumthiebautii)是三亞灣內固氮生物的優勢種,已成為三亞灣的一個重要氮源.林峰[32]的研究發現,北部灣固氮速率存在南高北低的特點,受溫度、無機營養鹽等理化因素的影響.固氮作用可支持北部灣近2%初級生產力的氮需求[32].束毛藻曾經是大亞灣的赤潮種[42],但是近年來束毛藻水華未見報道,可能暗示在日益加劇的人類活動壓力下,該區域固氮生物組成在過去數十年間可能發生了顯著的變化.已有研究顯示,大亞灣生態環境正在發生深刻的變化[43],對于其他南海半封閉海灣亦然.
表1給出了部分研究所報道的南海各區域生物固氮速率.南海生物固氮速率的變化范圍為0~8.5 nmol/(L·d).相對而言,上升流區域的固氮作用更為活躍,在海盆區也可觀測到活躍的固氮作用,而受淡水徑流影響的珠江口和三亞灣固氮速率則相對較低.由于調查季節不同,采用方法也有差異,結果之間的可比性較差.除此之外,各區域固氮生物的組成有所差異,且會隨季節發生變化.在受黑潮影響的北部陸架區及越南沿岸區域,束毛藻是固氮生物的重要組分[13,15,31],而在珠江口附近海域和開闊南海海盆區,束毛藻的豐度非常小,固氮生物主要由單細胞藍藻組成,同時存在大量的固氮變形菌[18,25,33].不同類型固氮生物的固氮能力不同,這也是造成各區域生物固氮速率差異的重要原因.整體而言,南海生物固氮速率的報道值分布在北大西洋報道值的低值范圍內[1,7],與北太平洋的報道值處于相同量級[3].

表1 南海固氮速率的研究報道
2 南海生物固氮作用的研究方向
2.1 拓展調查范圍
目前,在全球范圍內,近岸海域中生物固氮的研究正逐漸增加[44-46].沿岸海域理化環境復雜,受人類活動影響很大,固氮作用的調控機制可能迥異于開闊水體.這些研究表明沿岸海域中也可觀測到較強的固氮作用.沿岸海域理化環境復雜,既受陸源物質的影響,又可能受人類活動的影響,生物量及生物多樣性相對更高,有更大的研究空間.然而,南海中沿岸海域中生物固氮作用的相關報道目前仍較少,有待補充.
在過去20年,已有報道證明,南海中的固氮生物除束毛藻外還包括共生型藍藻(胞內植生藻)、單細胞藍藻、固氮變形菌等[13,25],其中單細胞固氮生物的生物量及固氮量甚至可以超過束毛藻等絲狀藍藻[15-16,26].事實上,許多非藍藻類固氮生物也廣泛分布于全球海洋[8-10].一些異養型的固氮生物,由于不受光的限制,具有更廣闊的分布范圍[9].已有報道指出:在無光層也可以觀測到異養固氮生物的固氮活動,引入的氮可占調查海域總固氮量的50%以上[47].目前,南海中生物固氮作用的研究仍多局限在真光層內.后續研究可將南海生物固氮作用的調查范圍由真光層拓展到弱光層.
2.2 探索調控機制
生物固氮作用受多種因素共同調控[1,3,48].南海生物固氮作用的研究多數關注無機營養鹽和鐵對固氮作用的調控[11,15,20,36].事實上,除無機營養鹽和鐵的含量之外,還有諸多因素可能調控固氮作用的分布.
近年來,有機物與生物固氮作用之間的關系被廣泛研究,已有許多報道指出:海水中的溶解有機質可能會刺激生物固氮作用[47-49],甚至自養型的束毛藻也會吸收水體中的溶解態有機質[50].生物固氮是一個極其耗能的過程,有機質可能是固氮生物的一個重要的能量來源.此外,鐵是固氮酶合成與表達所必須的元素.海水中有機質的存在可能為生物提供了有效的鐵的有機配體,很大程度上提升了鐵的生物可利用性,緩解了生物固氮過程受到的鐵限制[2,51].南海受湄公河、珠江等淡水的徑流影響[17,33],淡水徑流攜帶大量陸源有機質入海,除此之外,沿岸的海灣由于受人類活動的影響,也可能向南海中輸入大量的有機物質,因此,未來可在南海深入研究有機質對固氮作用的調控機制.
在氣候變化的大背景下,海洋酸化對南海固氮作用的影響可能亦不能小覷[14].固氮作用同全球氣候密切相關,因此在這方面的研究需進一步深化.由于生物固氮作用是在多種因素的共同調控之下進行的,今后的研究可更多關注多種因子之間的協同作用.
2.3 分析固氮與固碳過程的耦合
固氮作用與固碳作用之間可能存在復雜的聯系.非固氮生物可利用固氮生物所提供的氮來滿足自身需求[1,3].同時,在其他營養物質方面,非固氮生物同固氮生物之間存在競爭作用,從而間接地調控著固氮過程[2,20].除了營養物質的競爭外,非固氮生物可能會通過釋放固氮生物可利用的溶解態有機質來間接地影響生物固氮過程:海洋中存在大量的溶解態有機質,但其中生物可利用的僅占很小一部分[52],而浮游植物在光合作用后會釋放溶解態有機質到水體中[48,53],這可能為固氮過程提供了重要的能量來源[48,50].因此,海洋生物固氮作用與固碳過程之間存在密切的關聯.在南海,固氮作用與固碳作用二者的關系如何?是否存在初級生產者對固氮生物的調控?需要進一步研究加以回答.
為進一步探索南海生物固氮過程與固碳過程之間的耦合關系,首先可在實驗室進行培養實驗,通過結合細胞計數、穩定同位素分析等技術,探索碳和氮在固氮生物和非固氮生物之間的傳遞,以期獲得可應用于實際海洋的結論.
2.4 規范研究方法
生物固氮速率的測定方法主要包括乙炔還原法和15N 示蹤法[54].15N示蹤法直到20世紀90年代才開始廣泛應用[55].15N示蹤法通過向密閉體系中加入15N2標記氣體,經一段時間的培養后,測定該體系中固氮生物對15N2的吸收情況來測定固氮速率[55].南海中生物固氮速率的研究,在測定生物固氮速率方面使用的方法不一,一些研究中使用的方法可能會使結果偏離真值,且各項研究之間的可比性較差.在利用15N示蹤法測定生物固氮速率時,培養時間的不同也會造成測定結果的差異[16,56].今后,有必要對南海中生物固氮作用的調查方法進行規范與統一,一方面可以對南海中各區域生物固氮活動進行對比,另一方面可以真實地評估南海的氮收支情況,這對南海氮循環及碳循環而言具有重要意義.
3 總結與展望
南海是研究生物固氮作用的理想場所.南海固氮生物組成較為復雜,具有較大的時空可變性,主要包括束毛藻、胞內植生藻、單細胞藍藻和固氮變形菌等,其中單細胞固氮生物對南海生物固氮的貢獻不容忽視.南海的固氮速率與其他熱帶、亞熱帶海區相近,其中海盆區的年固氮量可達3.6×107mol/a.整體而言,生物固氮作用對南海初級生產力的貢獻率較低.南海生物固氮作用受黑潮水入侵、陸地徑流的影響,除此之外,上升流為南海表層水體輸送大量的營養物質,從而間接調控著生物固氮作用.本文中結合生物固氮作用在全球范圍內的研究現狀,指出今后需進一步拓展南海中生物固氮作用的調查范圍,并對其調控因素以及其與海洋生物固碳過程之間的耦合進行更全面的分析,同時對生物固氮的調查方法進行規范與統一,進而為研究南海中的海洋氮循環提供準確的信息.
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
今日農業(2021年9期)2021-11-26 07:41:24
科學大眾(2021年9期)2021-07-16 07:02:54
發明與創新·小學生(2021年3期)2021-03-25 11:48:49
軍事文摘(2020年20期)2020-11-28 11:42:50
航空世界(2020年10期)2020-01-19 14:36:20
中國科技博覽(2016年2期)2016-04-25 20:32:39
小學生導刊(2016年34期)2016-04-11 00:49:44
