高黎貢山北段東、西坡捕食線蟲真菌分布格局研究
2019-01-04 08:09:12王家亮劉碩然周漢宇房以好楊曉燕
大理大學學報 2018年12期
關鍵詞:物種
王家亮 ,劉碩然 ,周漢宇 ,房以好 ,肖 文 ,楊曉燕 *
(1.大理大學公共衛生學院,云南大理 671000;2.大理大學東喜瑪拉雅研究院,云南大理 671003;3.云南省高校洱海流域保護與可持續發展研究重點實驗室,云南大理 671003)
微生物物種資源極為豐富,是地球生物多樣性的重要組成部分〔1〕。微生物的空間分布格局受到眾多驅動因子的影響,主要包括當代環境條件(溫度、降水、光照、土壤養分、pH)和歷史因素(地理阻隔、物理屏障、擴散限制等)〔2-3〕,而溫度、降水等環境因子與山地微生物物種豐富度有著密不可分的聯系。研究人員發現坡向對土壤的溫度、含水量等理化性質均有影響〔4〕。不同坡向上的水分和光照強度存在差異,從而影響土壤微生物的生長和分布格局〔5〕。現階段有關坡向對土壤微生物分布格局影響的研究還較少。
高黎貢山國家級自然保護區東坡垂直落差高達6 000 m,氣候變化復雜,是全球生物多樣性的重點區域,具有非常重要的生態意義〔6〕。高黎貢山西坡巨大的垂直落差造成氣候、土壤和植被的垂直地帶性差異明顯〔7〕。高黎貢山坡向的差異使溫度、降水等環境因子的變化顯著,因此可作為研究坡向對土壤微生物分布格局影響的理想區域。
捕食線蟲真菌是一種既可以營腐生生活,也可以以營養菌絲特化形成捕食器官捕食線蟲的真菌類群,具備較強的環境適應能力〔8〕。有關捕食線蟲真菌的分離、純化技術和鑒定方法已很成熟。綜合以上特點,捕食線蟲真菌是作為微生物多樣性分布格局研究的理想對象。
因此,本研究擬以滇西北高黎貢山國家自然保護區為研究區域,以捕食線蟲真菌為研究對象,研究不同坡向上捕食線蟲真菌的分布格局,探究坡向對捕食線蟲真菌空間分布格局的影響。
1 材料與方法
1.1 樣品采集土樣采自貢山縣至福貢縣的獨龍江公路,每1 km設置1個樣點,以獨龍江隧道為分界線,將高黎貢山北段分為東、西坡,每個采樣點對角線5點取樣(5~30 cm深土層),充分混勻,取約200 g裝于采樣袋,帶回實驗室低溫密封保存,并于1周內進行撒樣處理。共100個樣點,其中東、西坡各50個樣點,共500份土樣,樣點采集的海拔范圍在1 400~3 400 m。
1.2 地理、環境指標在采集土樣時,記錄該區域樣品的采集時間、經度、緯度、海拔和坡向等指標。
1.3 培養基的制備玉米瓊脂培養基(CMA)和馬鈴薯葡萄糖瓊脂培養基(PDA)參照文獻〔8〕進行制備。
1.4 誘餌線蟲的制備用貝爾曼氏法制備全齒復活線蟲(Panagrellus redivivus)幼蟲懸液,0.1 mL約含線蟲500條,備用〔8〕。
1.5 分離純化參照李天飛等〔9〕的誘餌平板法進行分離純化。
1.6 捕食線蟲真菌的鑒定通過菌株形態特征及捕食器官類型〔10〕進行形態學鑒定,同時通過ITS(internal transcribed spacer region of the ribosomal RNA gene,核糖體RNA上的非轉錄間隔區)和TUB(β-tubulin gene,微管蛋白編碼基因)基因序列同源性分析進行分子生物學鑒定〔11-12〕,兩者結合進行種屬的鑒定。
1.7 數據處理 (1)檢出率(occurrence frequency,OF)=(某個種出現的土樣數∕總土樣數)×100%〔13〕。
(2)總檢出率=(檢出菌的土樣數∕總的土樣數)×100%。
(3)物種豐富度:種群中物種數目的多少。物種豐富度往往作為生物多樣性的代表性測度,具有簡單直觀等優點。
運用Excel 2016軟件對原始數據進行整理分析。
2 結果
2.1 東、西坡捕食線蟲真菌分布的整體差異本研究從500份土樣中共分離和鑒定出2屬15種120株捕食線蟲真菌,其中東坡共檢出2屬12種46株,西坡共檢出2屬13種74株。見表1。西坡的物種豐富度和檢出率均高于東坡。見圖1。
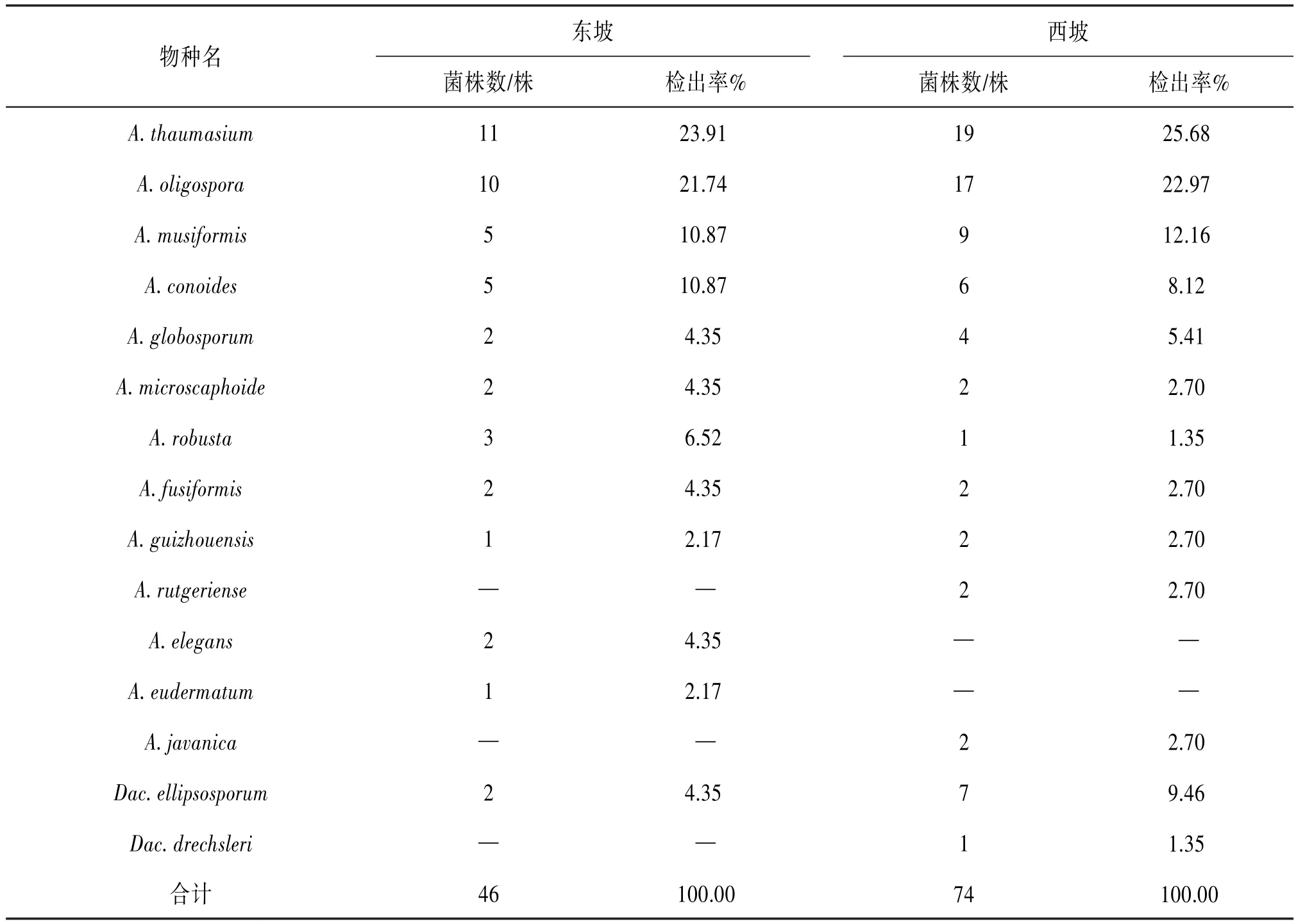
表1 東、西坡捕食線蟲真菌檢出情況
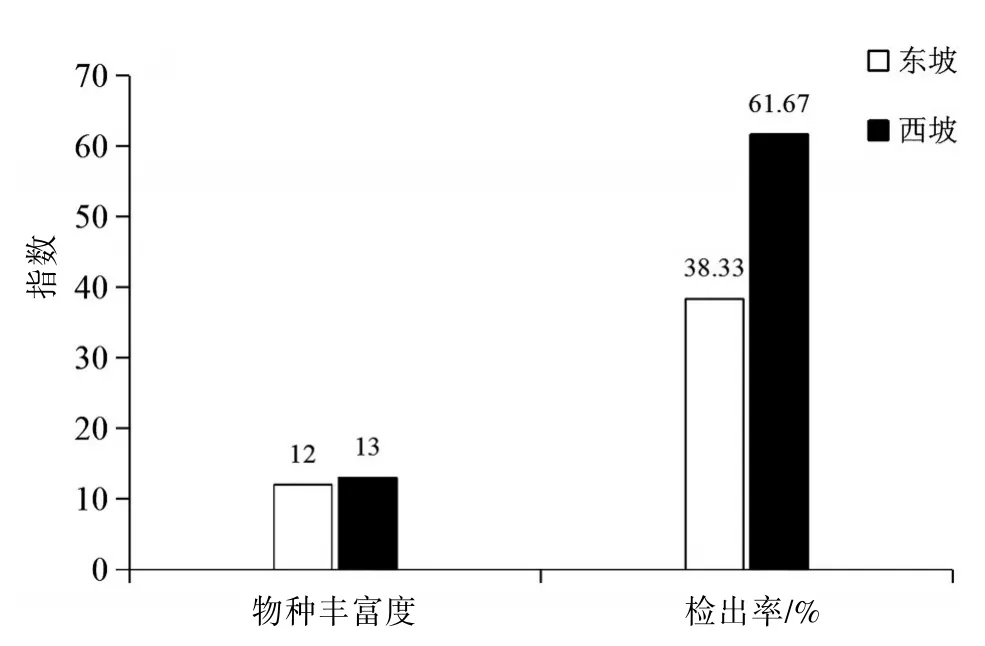
圖1 高黎貢山東、西坡各指數的比較
2.2 東、西坡不同坡位捕食線蟲真菌的多樣性差異將高黎貢山研究區域分為5個坡位,分別為下坡位(1 400~1 800 m)、中下坡位(1 800~2 200 m)、中坡位(2 200~2 600 m)、中上坡位(2 600~3 000 m)、上坡位(3 000~3 400 m)。對高黎貢山東、西兩坡相同坡位的捕食線蟲真菌物種豐富度和檢出率進行比較。結果表明東、西坡捕食線蟲真菌的物種豐富度和檢出率隨著坡位的升高均呈先上升后下降的趨勢,其中東坡各指數均在中下坡位處呈現最大值,西坡物種豐富度在中下坡位處呈現最大值,檢出率在中坡位處呈現最大值。見圖2~3。
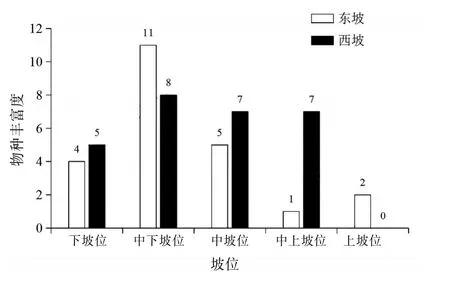
圖2 高黎貢山東、西坡相同坡位物種豐富度的比較
2.3 東、西坡捕食線蟲真菌優勢種和特有種的差異對高黎貢山東、西坡的優勢種和特有種進行分析,結果表明A.thaumasium為高黎貢山東、西兩坡的優勢種,東坡特有種為A.elegans和A.eudermatum,西坡特有種為A.rutgeriense、A.javanica和Dac.drechsleri。見圖4。
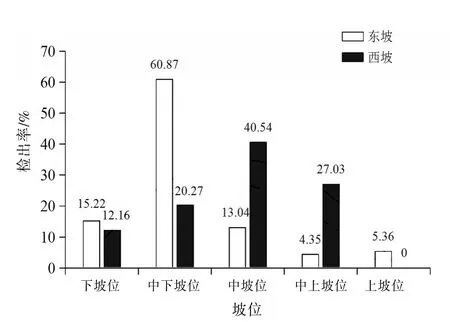
圖3 高黎貢山東、西坡相同坡位檢出率的比較
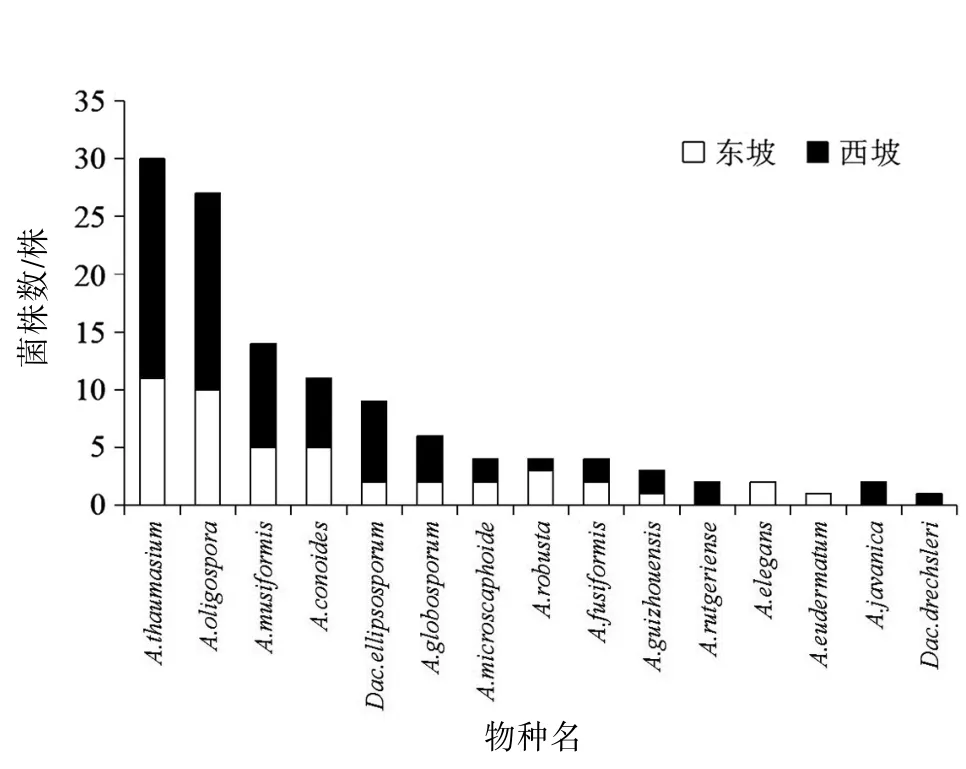
圖4 高黎貢山東、西坡每個樣點菌株檢出數
3 討論
3.1 東、西坡捕食線蟲真菌的整體差異高黎貢山東坡共檢出2屬12種46株捕食線蟲真菌,西坡共檢出2屬13種74株捕食線蟲真菌,表明高黎貢山西坡的捕食線蟲真菌資源較東坡更為豐富。由于西南和東南季風大氣水分輸送、流域高山河谷地貌的阻擋因素以及地面水分蒸散的共同作用,形成滇西北三江并流區域特殊的多元立體氣候特征:三江并流區域降水量自西向東逐步減少,蒸發量逐步增大〔14〕。高黎貢山西坡屬于中亞熱帶西南季風氣候,由于高黎貢山、碧羅雪山山脈均呈南北向排列,當印度洋氣流由西南向東北行進時,水氣遇到高聳的山脈上升之后逐漸形成降雨,使該區域常年具有豐沛的降雨。高黎貢山西坡小氣候十分明顯,立體氣候尤為突出,一山之內熱、溫、寒三帶氣候兼有,雨量特別充沛,年均降雨量達4 000 mm,年平均氣溫為16℃,為東南亞3個多雨中心之一〔15〕。東坡的氣候為半濕潤氣候,據貢山縣氣象資料顯示,該區域年平均氣溫為14.7℃,年均降雨量為1 667.4 mm。因此,與坡向相關的溫度和降雨量的差異可能是導致西坡捕食線蟲真菌物種豐富度、菌株檢出數和檢出率高于東坡的原因。
3.2 東、西坡相同坡位捕食線蟲真菌差異東、西坡捕食線蟲真菌的物種豐富度、菌株檢出數和檢出率隨著坡位(海拔)的升高,呈先上升后下降的趨勢。Mccain〔16〕發現土壤微生物生物量的周轉及其數量隨著土壤含水量的提高而有所增加。張萍等〔17〕對高黎貢山土壤微生物分布及生化特性的研究表明高黎貢山土壤的含水量隨海拔高度的升高呈先上升后下降的趨勢,土壤含水量在中海拔段呈現最大值。相關研究表明高黎貢山土壤微生物的分布格局與生態環境有著密不可分的聯系,高黎貢山土壤微生物數量隨海拔的下降而增加,中海拔段土壤中物質能量代謝較快,微生物種類較為豐富,高海拔段由于溫度偏低的原因,物質能量代謝較為緩慢,微生物數量明顯減少,而低海拔段由于人為干擾的影響,植被破壞嚴重及氣候干燥、水分含量較低的原因,使得土壤微生物的數量有所下降〔17〕。因此,土壤含水量和溫度作為坡向的間接影響因子可能是高黎貢山捕食線蟲真菌物種豐富度和檢出率隨海拔的升高呈先上升后下降趨勢的原因。
3.3 東、西坡捕食線蟲真菌優勢種和特有種分布高黎貢山北段捕食線蟲真菌的優勢種群為A.thaumasium,這與楊浩然等〔18〕對云南文山進行捕食線蟲真菌多樣性的調查結果相一致。其原因可能是Arthrobotrys屬捕食線蟲真菌的腐生能力較強,對營養要求不嚴格,在不同生境(水生、陸生、糞生及植物根際、溫泉)中,該屬捕食線蟲真菌資源最為豐富,優勢種也都是在該屬中出現〔19-23〕。根據Gray〔24〕的調查發現,不同捕器類型的捕食線蟲真菌對營養的要求不同,產黏性菌網的種類因生長速度快,對營養需求不嚴格且腐生能力較強而處于優勢。而特有種的差異可能是不同的生態環境所導致,表明產不同捕食器官的捕食線蟲真菌對環境的適應能力存在差異,表現為不同的地理區域、不同的生態環境下,該類型真菌的優勢種和特有種存在差異。
綜上所述,溫度和降雨量作為坡向的間接影響因子可能是造成捕食線蟲真菌分布存在異質性的主要因素。
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
