土荊芥化感脅迫下蠶豆葉綠體超微結構和光合關鍵基因表達的變化
2019-01-18 02:45:54阿的魯驥馬丹煒
西南農業學報 2018年12期
阿的魯驥, 周 健,李 潔,馬丹煒
(1. 四川師范大學生命科學學院,四川 成都 610101;2. 西南民族大學青藏高原研究所,四川 成都 610064;3.四川省農業科學院農業信息與農村經濟研究所, 四川 成都 610066)
【研究意義】植物的光合作用是一系列復雜代謝反應的總和。由于步驟繁多、過程復雜,任何一個反應步驟受到影響都有可能影響到光合作用。生物體的所有生長發育過程都受基因表達的調控,同樣光合作用也是相關基因表達調控的結果[1]。【前人研究進展】光合作用過程對環境變化十分敏感,在逆境脅迫下,植物光合產物減少,植物生長發育受阻[2]。生物入侵已經成為導致全球生物多樣性下降的一個重要因素,擴散到新生境的入侵植物比原產地的個體具有更強的化感作用,使其在與其他種群爭奪環境資源的競爭中獲得優勢地位[3]。入侵植物釋放到周圍的化感物質具有較大的細胞毒性[4],這些物質通過細胞膜進入細胞后往往會破壞細胞的內部結構[5],并干擾受體植物的基因表達[6]。入侵植物土荊芥(ChenopodiumambrosioidesL.)釋放的揮發油及其主要成分α-萜品烯和對傘花素誘導周圍植物的根細胞[7]和氣孔保衛細胞[8]凋亡,并干擾了根尖細胞的抗氧化酶活性及其基因表達[9]。【本研究切入點】有關土荊芥揮發性化感物質是否影響與光合作用密切相關的細胞器葉綠體的結構以及光合關鍵基因表達的研究卻鮮有報道。【擬解決的關鍵問題】本研究采用盆栽試驗,以土荊芥入侵地廣泛栽培的農作物蠶豆(ViciafabaL.)為受體,運用透射電鏡技術和RT-PCR技術,研究了土荊芥揮發油及其主要成分α-萜品烯和對傘花素對受體植物幼苗葉片葉綠體超微結構和光合相關基因表達的影響,以期為深入探討土荊芥的入侵機制提供理論依據。
1 材料與方法
1.1 材料
供試土荊芥揮發油由本實驗室提供。色淡黃,密度843 mg/mL,其主要成分α-萜品烯和對傘花素的含量分別為151和156 mg/mL;α-萜品烯和對傘花素的標準品購自成都市銳可思生化試劑公司;蠶豆種子(成胡14#)購于成都市五塊石種子市場。
1.2 幼苗培養和化感脅迫處理
幼苗培養方法見本項目組前期研究[4]。待蠶豆幼苗長至4~5周進行化感脅迫處理;本研究共設置3個處理組(揮發油處理組、α-萜品烯處理組、對傘花素處理組)和1個對照組。在預實驗基礎上確定揮發油的處理劑量,將該處理劑量揮發油中對傘花素和α-萜品烯的含量定為二者的處理劑量;挑選長勢一致、生長良好的蠶豆幼苗若干盆,隨機分為4組,分別置于特制的、帶有密封蓋子的玻璃箱(長25 cm、寬20 cm、高40 cm)中,每箱均勻放置5盆,在每個箱底部正中央放置一個小培養皿;分別將80 μl揮發油、14.88 μl α-萜品烯和3.52 μl對傘花素滴加在玻璃箱中央的小培養皿中,迅速蓋上蓋子,并用凡士林封住玻璃箱口。對照組不作處理但同樣用凡士林封住玻璃箱口。將4組玻璃箱均置于25 ℃光照培養箱中處理3 h取出揮去揮發物,再放入培養箱中培養。每天處理時段為10:00-13:00,連續處理3 d,第4天取樣電鏡觀察和real-time PCR分析。
1.3 葉綠體超微結構的觀察
取處理第4天后的葉片,在中脈兩側剪取1 mm×1 mm的小塊若干,2.5 %戊二醛前固定,0.1 mol/L PBS(pH 7.4)漂洗,1 %鋨酸后固定, 0.1 mol/L PBS(pH 7.4)漂洗,乙醇梯度系列脫水,丙酮︰Epon 812包埋劑=1∶1的混合液滲透,Epon-812環氧樹脂包埋,Leica UC7切片機切片,鈾鉛雙染色(2 %醋酸鈾飽和水溶液和檸檬酸鉛分別染色15min),室溫干燥過夜。Tecnai G2 20 TWIN透射電子顯微鏡觀察并采集圖像。
1.4 葉片RNA的提取與RT-PCR分析
總RNA的提取: 取處理第4天后的葉片,用液氮充分研磨,按照天根生化科技有限公司的植物總RNA提取試劑盒操作說明提取蠶豆幼苗葉片的總RNA,除去DNA后分裝于EP管中,-80 ℃保存待測。
引物設計:用Primer Premier 5.0引物設計軟件,設計蠶豆核酮糖羧化酶基因(Rbcs-3C)、光系統ⅡCP47蛋白基因(PsbB)、ATP合成酶β亞基基因(beta)、質體藍素基因(PetE)和內參基因(EF-1-alpha)特異性引物,序列如下。EF-1-alphaF:5′-ACGAGGCTCTCACTGAGGCTCTTCC-3′;EF-1-alphaR:5′-CCTTGGCAGGGTCATCCTTGGAGTTG-3′;Rbcs-3CF:5′-CCGTGAGCACAACAAGTCACCAGGAT-3′;Rbcs-3CR:5′-GGTAAGCCGCAACAACTTCATCAAGC-3′;PsbBF:5′-CCGCTCTAACAATTCCGTCTCCGTCT-3′;PsbBR:5′-CGAAGAGTTGGTGCTGGGCTAGTCAA-3′;betaF:5′-TTCTTCACCGACGATGCGAGGTTGGA-3′;betaR:5′-GACTGATCCTGCTCCTGCCACGACAT-3′;PetEF:5′-GCTGTAAGTCTCACCAGGAGCGTTGA-3′;PetER:5′-AGCAATGCCTTGGCTGTTGAAGTCTT-3′。
引物均由上海生工生物工程技術服務有限公司合成。
RT-PCR分析:使用實時熒光定量PCR儀(PIKORed 96,美國Thermo Fisher)進行RT-PCR分析。反應體系為20 μl,含SYBR Primix ExTaqⅡ10 μl, PCR Forward Primer 0.8 μl,PCR Reverse Primer 0.8 μl,cDNA 2 μl和ddH2O 6.4 μl;熒光 PCR 反應程序為:95 ℃ 1 min,95 ℃ 5 s,62 ℃ 30 s,45個循環;每處理重復3次;使用Thermo Scientific PikoReal軟件分析PCR過程的CT(Threshold cycle)值。
1.5 數據統計
參照Williamson et al.[10]的方法計算基因相對表達量的化感敏感指數(RI):RI=1-C/T(T≥C);RI=T/C-1(T 用透射電子顯微鏡觀察發現,對照組中的蠶豆葉片葉綠體的形態結構正常(圖1 a, e, i),葉綠體長梭形,膜結構清晰完整,排列有序,并緊貼于細胞壁,葉綠體中基質均勻且濃密,基粒類囊體垛疊整齊緊密,淀粉粒發育正常;揮發油處理組(圖1d, h, l)中,葉綠體脫離細胞壁散亂分布于細胞中央,結構紊亂,表現為腫脹、膜結構受損明顯,葉綠體內部基質不均勻、基粒類囊體排列紊亂、淀粉粒消失、片層扭曲且排列松散;α-萜品烯處理組中(圖1 c, g, k),葉綠體出現腫脹現象,部分葉綠體脫離細胞壁,葉綠體膜和內部結構變化不明顯;對傘花素處理組(圖1 b, f, j)中,葉綠體超微結構與對照組相比無差異,表明對傘花素對蠶豆葉綠體超微結構沒有產生影響。 a, e, i: 對照組;b, f, j: 對傘花素處理組;c, g, k: α-萜品烯處理組;d, h, l: 揮發油處理組;CH:葉綠體;SG:淀粉粒;CW:細胞壁;Me:葉綠體膜;G:基粒;GL:基粒片層;N:細胞核a, e and i are control groups;b, f and j were cymene-treated groups; c, g and k are α-terpinene-treated groups;CH is Chloroplast; SG is Starch Grain; CW is Cell Wall; ME is Chloroplast Membrane; G is Grana; GL is Grana Lamella; N is Nucleus圖1 土荊芥揮發油、α-萜品烯和對傘花素作用下蠶豆幼苗葉片超微結構Fig.1 TEM micrographs of leave of Vicia faba exposure to the volatile oil from Chenopodium ambrosioides, α-terpinene and cymene 不同小寫字母表示P<0.05 的顯著水平Different lowercase letters indicates P<0.05圖2 土荊芥揮發油、α-萜品烯和對傘花素對蠶豆幼苗Rbcs-3C、PsbB、beta和PetE 基因相對表達量的影響Fig.2 Effects of volatile oil from Chenopodium ambrosioides L., α-terpinene and cymene on relative expression of Rbcs-3C, PsbB, beta and PetE in Vicia faba L. RT-PCR研究表明,在土荊芥揮發油、α-萜品烯和對傘花素作用下,蠶豆幼苗光合關鍵基因的表達發生了不同程度的變化(圖2)。其中,揮發油處理組中蠶豆幼苗光合關鍵基因表達的變化幅度最大,核酮糖羧化酶基因(Rbcs-3C)、光系統ⅡCP47蛋白基因(PsbB)、ATP合成酶β亞基基因(beta)、質體藍素基因(PetE)等4個基因的相對表達量均顯著高于對照組(P<0.05);與對照組相比,對傘花素處理組的Rbcs-3C和PsbB的相對表達量均下調,beta和PetE的相對表達量均上調,但只有PetE相對表達量與對照組的差異達到顯著水平(P<0.05);α-萜品烯處理組中,PsbB的相對表達量低于對照組,但未達到顯著水平(P>0.05),而Rbcs-3C、beta和PetE的相對表達量均顯著高于對照組(P<0.05)。由此表明,土荊芥揮發性化感物質不同程度影響了受體植物光合關鍵基因的表達,無疑會干擾受體植物的光合作用過程。 從圖2-A可見,揮發油處理組和α-萜品烯處理組之間Rbcs-3C的表達量差異不顯著(P>0.05),但二者的相對表達量均顯著高于對傘花素處理組(P<0.05);從圖2-B可知,PsbB的相對表達量在對傘花素處理組和α-萜品烯處理組之間無顯著差異(P>0.05),但二者與揮發油處理組之間的差異達到顯著水平(P<0.05);3個處理組的beta相對表達量均呈現顯著差異(P<0.05,圖2-C),而PetE的相對表達量則無顯著差異(P>0.05,圖2-D)。 3種揮發物對蠶豆幼苗光合關鍵基因化感作用的綜合效應分別為對傘花素0.525,α-萜品烯0.535,揮發油0.666,由此可見,揮發油對光合關鍵基因的相對表達量影響最大,而揮發油主要成分α-萜品烯和對傘花素的化感作用強度不及揮發油,說明揮發油中多種成分存在正向的協同效應。 4種基因相對表達量對土荊芥揮發油的敏感程度從大到小依次為PetE,Rbcs-3,CPsbB和beta。 葉綠體是植物體內葉綠素存在的部位,是進行光合作用的場所[11]。化感作用對葉綠體等細胞器超微結構的影響將直接改變受體植物的生長發育。葉綠體結構受損時,葉綠素合成酶活性受抑制,葉綠素降解酶活性增強,導致葉綠素的含量下降[12]。項目組前期研究發現,在土荊芥揮發油及其兩個主要成分α-萜品烯和對傘花素作用下,蠶豆幼苗的最大光化學效率(Fv/Fm)、實際光化學效率(ΦPsⅡ) 和葉綠素含量均呈現下降趨勢,土荊芥化感脅迫可能干擾了葉綠素的合成過程或引起葉綠素降解,從而降低了光合速率[13]。本研究結果表明,在土荊芥揮發油、α-萜品烯和對傘花素的作用下,蠶豆幼苗葉片的葉綠體超微結構受到不同程度的破壞,其中揮發油處理組中葉綠體受損程度最為嚴重,葉綠體散亂分布于細胞中央,結構紊亂,表現為腫脹、膜結構受損明顯,葉綠體內部基質不均勻、基粒類囊體排列紊亂、淀粉粒消失、片層扭曲且排列松散。相對而言,對傘花素對葉綠體的損害較小。由此可見,土荊芥化感脅迫抑制鄰近植物的生長發育[14-15],無疑與其化感物質破壞了葉綠體超微結構、影響光合作用過程有關。 化感物質通過上調或下調某些基因,或誘導新基因的表達,影響受體植物相關基因的表達調控[16]。例如,珍珠繡線菊(SpiraeathunbergiiBl.)釋放的順式肉桂酸改變了擬南芥(ArabidopsisthalianaL.)237個基因的表達量[17],土荊芥揮發性化感物質下調了玉米(ZeamaysL.)抗氧化酶基因的表達[9]。光合作用涉及多個相互協調且復雜的反應步驟,其每一步反應均容易受到各種環境脅迫的影響[18-19]。本研究結果表明,在揮發油、α-萜品烯和對傘花素作用下,與光合作用有關的關鍵基因表達發生了不同程度的變化。整體來看,大部分處理中,基因表達表現為上調,僅僅3個處理的基因表達下調(對傘花素處理組的Rbcs-3C和PsbB;α-萜品烯處理組的PsbB)。PsbB和PetE的表達產物是光合電子傳遞鏈的重要成員,beta的表達產物與葉綠體ATP的合成有關,而Rbcs-3C表達異常將直接改變受體植物的碳同化過程。本研究結果顯示,土荊芥釋放到環境中的揮發性化感物質干擾了蠶豆幼苗光合關鍵基因的正常表達,打破了蠶豆幼苗的光合作用過程各個環節協調性,進而影響了幼苗的正常生長。 土荊芥揮發油的成分十分復雜[22],這些成分在植物化感作用中的地位不盡相同,且可能相互之間可能存在著拮抗效應或協同效應。從本研究結果可見,揮發油對葉綠體結構的損害和對光合作用關鍵基因表達的影響最大,α-萜品烯次之,對傘花素的影響最小。這一結果與本項目組前期的研究結果相同[7-9, 20-21]。綜合分析可知,土荊芥向周圍環境釋放的揮發性化感物質之間存在著協同作用,進而使揮發油的化感效應大于其主要成分α-萜品烯和對傘花素。 在土荊芥揮發油、α-萜品烯、對傘花素作用下,蠶豆幼苗葉片葉綠體的超微結構受損,葉綠體脫離細胞壁散亂分布于細胞中央,結構紊亂,表現為腫脹、膜結構受損明顯,葉綠體內部基質不均勻、基粒類囊體排列紊亂、淀粉粒消失、片層扭曲且排列松散;核酮糖羧化酶基因(Rbcs-3C)、光系統ⅡCP47蛋白基因(PsbB)、ATP合成酶β亞基基因(beta)、質體藍素基因(PetE)等光合關鍵基因的表達異常。綜上所述,破壞受體植物葉綠體結構、干擾光合作用關鍵基因表達是土荊芥化感脅迫的機制之一。2 結果與分析
2.1 土荊芥揮發油、α-萜品烯和對傘花素對蠶豆幼苗葉綠體超微結構的影響

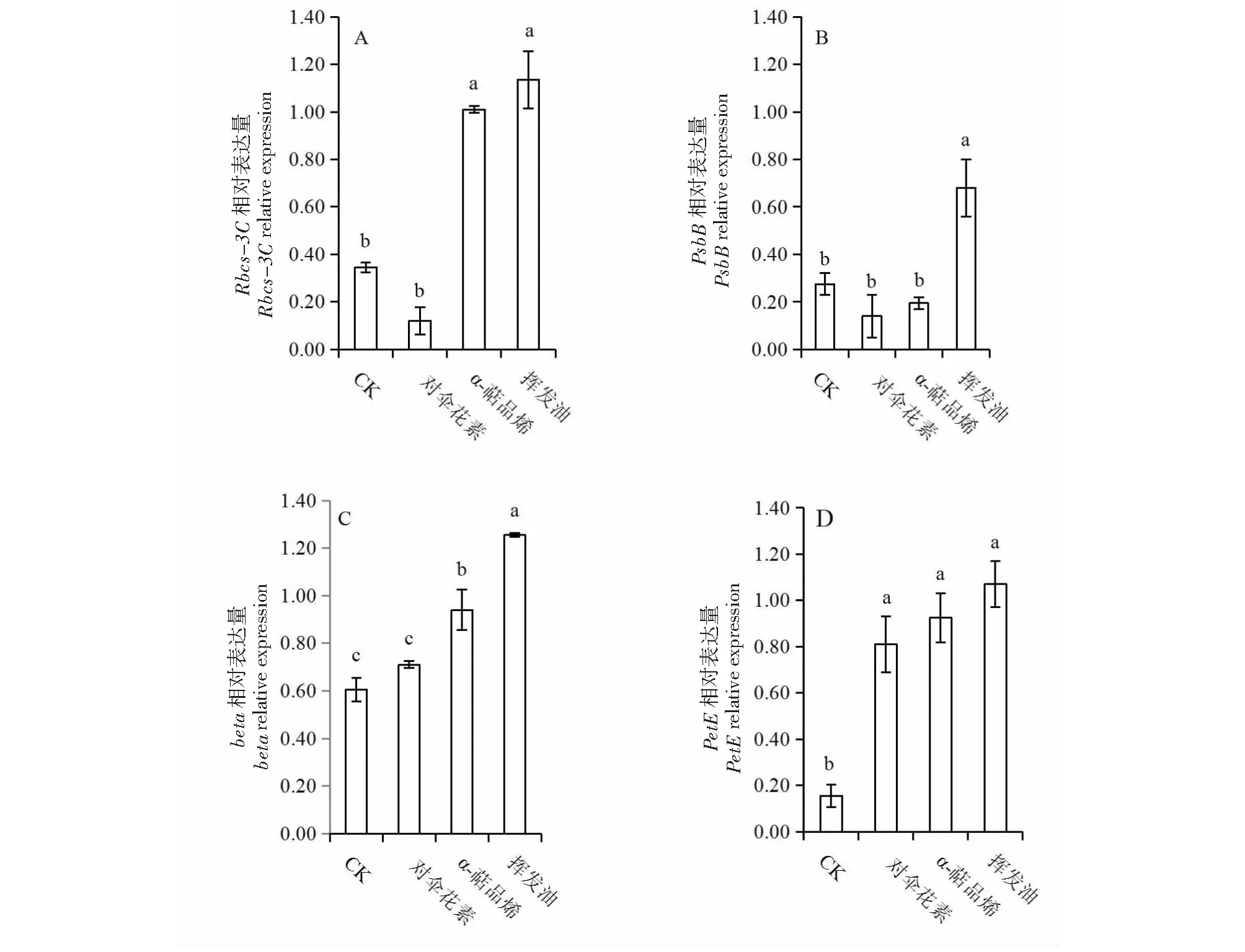
2.2 土荊芥揮發油、α-萜品烯和對傘花素對蠶豆幼苗光合作用相關基因表達量的影響
3 討 論
4 結 論
