團頭魴和斑馬魚鱗片的發生發育過程比較研究
2019-01-22 02:30:52王志強耿瑞靜王衛民
淡水漁業 2019年1期
王志強,劉 寒,耿瑞靜,王衛民
(華中農業大學水產學院,農業部淡水生物繁殖重點實驗室,武漢 430070)
魚鱗作為魚類重要的皮膚衍生物,由皮膚原始干細胞定向分化而來[1]。魚類鱗片和鰭條一樣,同屬魚類外骨骼,除對魚體起到保護作用外[2-3],魚類鱗片形態結構也是研究魚類分類特征、生存環境和生長特性的重要依據[4-5]。對于鱗片的發生位置和覆蓋過程的研究,不僅可以了解魚類的生長規律,判斷魚苗生長狀況,還能夠對魚類的演化過程和親緣關系進行探討和鑒定[6-8]。此外,有研究表明不同物種的魚類之間的鱗片發育過程差異不大[9],但其發生的位置和覆蓋方式則有很大不同[10-11]。
團頭魴(Megalobramaamblycephala)隸屬于鯉形目(Cypriniformes)鯉科(Cyprinidae)鲌亞科(Culterinae)魴屬,是我國特有的草食性經濟魚類[12]。為合理保護和開發利用團頭魴資源,諸多學者針對團頭魴的早期發育、人工繁殖與養殖等開展了大量研究工作[13-15]。在其鱗片相關研究中,孟慶聞等[16]在對團頭魴相關器官進行觀察研究時,對團頭魴鱗片的發育進行了簡要概括。熊金林等[17]對低齡團頭魴鱗片上的環紋與生長之間的關系做了探究。但是關于團頭魴鱗片的發生發育過程及與斑馬魚(Daniorerio)的比較研究尚未見報道。本研究主要采用形態學和組織學方法系統地比較研究了團頭魴稚魚和斑馬魚稚魚鱗片的發生位置和覆蓋過程,初步闡明了團頭魴和斑馬魚鱗片發育的基本規律,探討了鱗片覆蓋與個體年齡和生長的關系,為探討魚類鱗片的發育模式提供了豐富的材料,為進一步研究魚類鱗片發生發育的分子機制奠定基礎。
1 材料與方法
1.1 實驗魚來源與管理
團頭魴親本均來自湖北省鄂州市國家級團頭魴良種場。經人工授精獲得團頭魴胚胎,在25 ℃水溫中靜水孵化,充氧泵充氧。出膜后仔魚放入白色塑料盒(80.0 cm×55.0 cm×20.0 cm)中喂養,水溫(24.5±0.5)℃。出膜3 d后,開始投喂人工孵化的鹵蟲無節幼體。出膜14 d后,開始投喂浮游動物,每天早晚各一次。及時清理食物殘渣及糞便,每3 d換水一次。
斑馬魚親本均購自國家斑馬魚資源中心,放入斑馬魚循環系統內養殖。人工配對得到斑馬魚胚胎,收集至玻璃皿中孵化,水溫控制在28 ℃,每天換水一次。出膜3 d后轉至白色盒子中暫養,開始喂食草履蟲,每天早晚各一次。出膜20 d后,轉至較大白盒子中,充氧泵充氧。及時清理食物殘渣及糞便,每3 d換水一次。
1.2 樣本的收集與處理
第一批團頭魴出膜后20 d開始取樣一直到出膜后80 d,斑馬魚出膜后25 d開始取樣一直到出膜后55 d。每間隔2 d取樣一次,每次取樣10尾,用4%多聚甲醛固定,其中5尾用于茜素紅染色觀察鱗片出現的位置及鱗片的覆蓋情況,5尾用于組織切片觀察。
1.3 魚體鱗片染色與組織切片觀察
鱗片染色的方法結合李思發[11]與鮑寶龍等[18]的茜素紅染色法稍作調整,具體如下:(1)將固定于4%多聚甲醛中的標本用ddH2O漂洗30 min,重復3次;(2)用TBST(1 L體積:6 g Trisbase,pH 7.4,8.275 g NaCl,1 mL Trition X-100)溫和漂洗,進一步除去殘留甲醛和樣本脂肪組織;(3)將魚浸泡在1% KOH中,滴加1% H2O2,強光下照射除去大部分色素,當標本表面出現氣泡時終止反應(具體時間根據魚體大小調整);(4)待色素去除后,用蛋白酶液(100 mL體系:65 mL ddH2O,35 mL飽和硼酸鈉鹽,1 g胰蛋白酶)消化肌肉組織,直至魚體變透明;(5)將透明樣本放入1% KOH溶液中,慢慢滴加茜素紅溶液(1 g茜素紅溶于100 mL 0.5% KOH),待魚體表面染成淺紫色為止;(6)將標本放入1% KOH溶液中漂洗,除去多余染液后在體式顯微鏡下觀察、拍照。
對4%多聚甲醛固定的樣本進行常規石蠟切片,分別進行H&E和Masson染色,在正置顯微鏡下進行鱗片發生的組織學觀察、拍照。
1.4 數據處理
用鱗片覆蓋率(鱗片出現的排數)作為判斷鱗片覆蓋的指標。應用軟件SPSS 17.0對數據進行相關性分析,并繪制鱗片覆蓋率與日齡、體長的關系曲線。
2 結果
2.1 鱗片發生的位置及覆蓋過程
團頭魴在體長15.5 mm之前未觀察到有鱗片出現(圖1A)。出膜后24 d,體長達到16.3~19.7 mm時,團頭魴鰓蓋后緣側線處開始出現鱗片(圖1B),之后從該位置沿側線向尾柄部延伸,同時也沿鰓蓋后緣上下擴展,前者的速度要快于后者,并且沿鰓蓋后緣向下要早于向上覆蓋。出膜26 d,體長達到19.9~20.6 mm時,團頭魴軀干前部的側線下鱗已覆蓋兩排,而側線上鱗還未發現(圖1C)。出膜后30 d,體長達到21.1~21.8 mm時,魚體軀干軸下側鱗片基本覆蓋完全(圖1D)。出膜后34 d,體長達到22.3~23.2 mm時,鱗片開始向脊椎上側發展,尾部側線以上部位也開始有鱗片出現,并與軀干側線以上部位出現的鱗片慢慢匯合(圖1E)。出膜后44 d,體長達到23.9~24.6 mm時,團頭魴全身鱗片基本覆蓋完成(圖1F)。
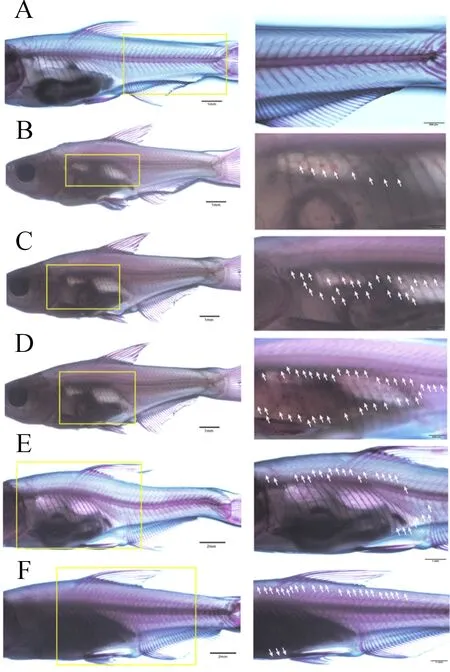
圖1 不同發育時期團頭魴鱗片覆蓋過程Fig.1 The scales development at different stages of M.amblycephala(A)出膜后第22 d,體長14.54 mm;(B)出膜后第24 d,體長19.62 mm;(C)出膜后第26 d,體長20.33 mm;(D)出膜后第30 d,體長21.55 mm;(E)出膜后第34 d,體長22.80 mm;(F)出膜后第44 d,體長24.12 mm
斑馬魚鱗片的起始位置和覆蓋方式與團頭魴有所不同。斑馬魚在體長約6.9~7.1 mm之前未觀察到有鱗片出現(圖2A)。出膜后39 d,體長達到8.5~9.2 mm時,在斑馬魚尾柄側線處首先出現鱗片,隨后鱗片開始由尾部沿側線向前開始覆蓋,同時側線上下兩側也開始出現鱗片(圖2B)。當體長達到9.7~10.2 mm時,斑馬魚尾部及側線鱗已覆蓋完全,側線上、下鱗已有1~2排出現(圖2C)。當體長達到10.9~11.5 mm時,斑馬魚側線上、下鱗已出現2~3排,魚體鱗片總排數達到5排(圖2D)。隨著鱗片繼續沿側線向上、下擴展,當體長達到11.6~11.9 mm時,斑馬魚體表除腹部以及背部以外,大部分區域鱗片已基本覆蓋(圖2E)體長達到12.1~12.5 mm時,斑馬魚全身鱗片覆蓋完全(圖2F)。

圖2 不同發育時期斑馬魚鱗片覆蓋過程Fig.2 The scales development at different stages of D.rerio(A)出膜后第33 d,體長7.04 mm;(B)出膜后第39 d,體長8.96 mm;(C)出膜后第45 d,體長9.93 mm;(D)出膜后第45 d,體長11.30 mm;(E)出膜后第43 d,體長11.63 mm;(F)出膜后第47 d,體長12.20 mm
2.2 鱗片發生的組織學觀察
為了清楚地研究團頭魴和斑馬魚鱗片發生的主要過程,實驗采用不同的組織切片染色方法,即H&E染色和Masson染色對其進行了觀察,結果表明團頭魴的鱗片發生過程主要經歷了以下幾個階段:
形態發生早期:成纖維細胞在表皮的生發層下聚集(圖3A,圖4A),隨著成纖維細胞在表皮與真皮臨界處的大量出現,形成一層單獨細胞層(圖3B,圖3C,圖4B)。
形態發生晚期:在表皮與真皮臨界處的成纖維細胞繼續分化成更大的細胞,開始形成鱗片乳突(圖3D,圖4C,圖4D)。
分化早期:鱗片乳突不斷分化成為兩層鱗片生成細胞,并不斷分泌骨質,形成骨質薄片即為鱗片原基(圖3E)。鱗片上下層出現新的真皮細胞層,形成包圍鱗片的小囊,即鱗囊(圖3F,圖4E,圖4F)。
分化晚期:鱗囊繼續擴大,鱗片基質更加明顯(圖3G,圖4G)。
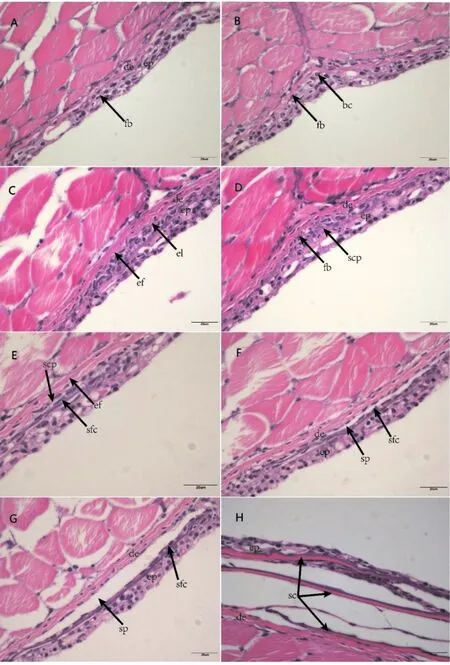
圖3 團頭魴稚魚鱗片發生的組織結構(H&E染色)Fig.3 The histology of scales development in juvenile M.amblycephala(H&E staining)(A-C)形態發生早期:(A)皮膚的表皮和真皮;(B)成纖維細胞開始在表皮層發生聚集;(C)成纖維細胞進入真皮層中部;(D)形態發生晚期:成纖維細胞堆積形成乳突;(E-F)分化早期:(E)鱗片乳突不斷分泌骨質,鱗片原基形成;(F)鱗囊形成;(G)分化晚期:鱗囊繼續擴大,鱗片基質更加明顯;(H)折疊期:表皮在鱗片末端折疊,鱗片形成ep:表皮;de:真皮;fb:成纖維細胞;bc:基底細胞;el:外層細胞;ef:彈性纖維;scp:鱗片乳突;sfc:鱗片形成細胞;sp:鱗囊;sc:鱗片
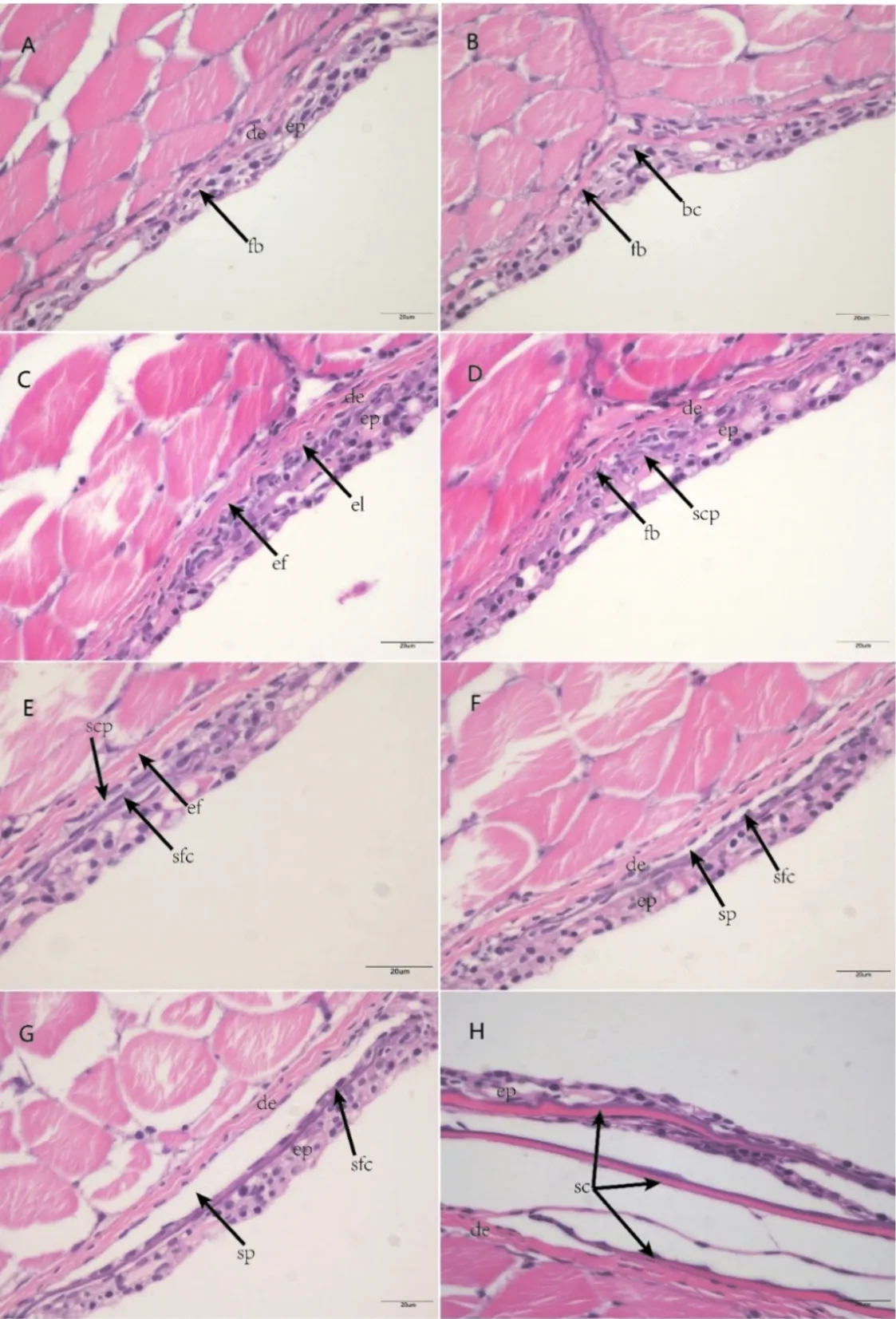
圖4 團頭魴稚魚鱗片發生的組織結構(Masson染色)Fig.4 The histology of scale development in juvenile M.amblycephala(Masson staining)
折疊期:表皮開始在鱗片后端邊緣折疊,鱗囊進一步擴大,相鄰的鱗片發生重疊,形成成熟的鱗片(圖3H,圖4H)。
經研究發現,斑馬魚的鱗片發生過程與團頭魴基本相同,也主要經歷了形態發生早期、形態發生晚期、分化早期、分化晚期和折疊期,共五個時期,各個發育時期的具體變化與團頭魴相同,詳見圖5、圖6。
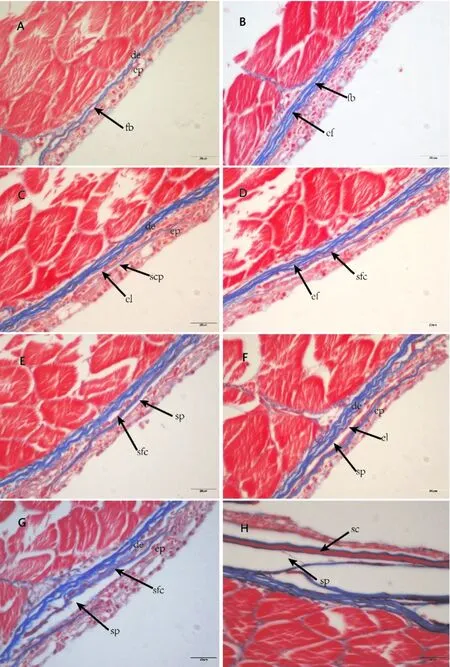
圖5 斑馬魚稚魚鱗片發生的組織結構(H&E染色)Fig.5 The histology of scales development in juvenile D.rerio(H&E staining)A-B形態發生早期:(A)成纖維細胞開始在表皮生發層聚集;(B)成纖維細胞進入真皮層中部;(C)形態發生晚期:成纖維細胞堆積形成乳突;D-E分化早期:(D)鱗片乳突不斷分泌骨質,鱗片原基形成;(E)鱗囊形成;F-G分化晚期:(F)鱗囊繼續擴大,鱗片基質更加明顯;(H)折疊期:表皮在鱗片末端折疊,鱗片形成。
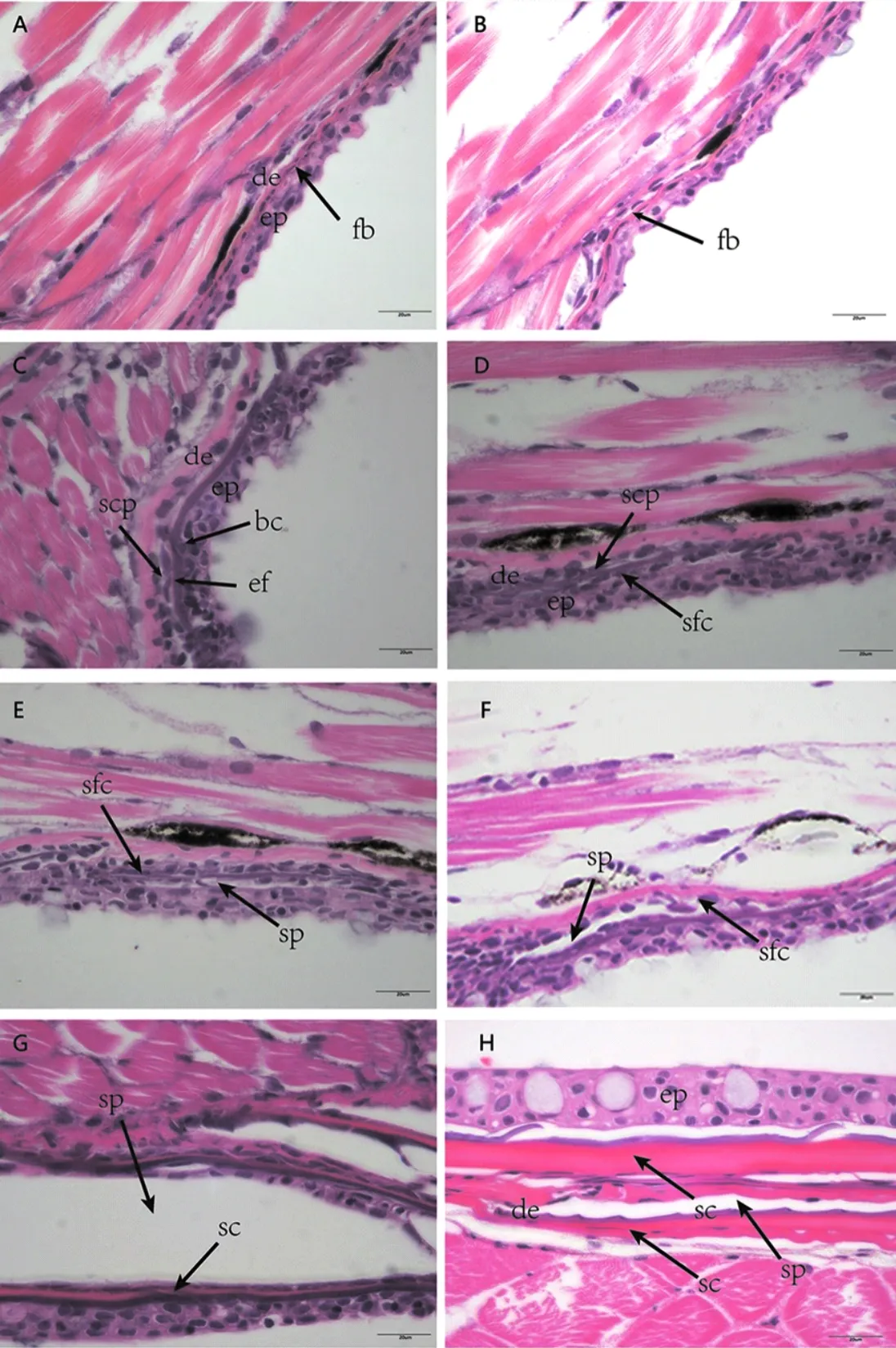
圖6 斑馬魚稚魚鱗片發生的組織結構(Masson染色)Fig.6 The histology of scales development in juvenile D.rerio(Masson staining)
2.3 鱗片覆蓋與體長和日齡的關系
鱗片覆蓋率(coverage rate,CR)指標,與體長(body length,L)和出膜后的日齡(days post hatching,D)的關系如圖7所示。團頭魴鱗片覆蓋率與體長的關系為CR=-0.000 7L3+0.040 3L2- 0.683 3L+3.560 1(R2=0.794 5,P=0.000)(圖7A);團頭魴鱗片覆蓋率與日齡的關系為CR=0.000 0D3- 0.000 3D2+0.039 0D- 0.684 5(R2=0.412 9,P=0.000)(圖7B)。通過SPSS 17.0軟件計算得到團頭魴鱗片覆蓋率與體長的相關系數為0.864,相關性較強(r=0.864,P=0.000),團頭魴鱗片覆蓋率與日齡相關系數為0.609,成弱相關性(r=0.609,P=0.000)。斑馬魚鱗片覆蓋率與體長的關系為CR=-0.004 8L3+0.162 1L2- 1.609L+4.951 7,(R2=0.803,P=0.000)(圖7C);斑馬魚鱗片覆蓋率與日齡的關系為CR=-0.000 3D3+0.029 5D2- 1.059 9D+12.169(R2=0.524 5,P=0.000)(圖7D)。斑馬魚鱗片覆蓋率與體長的相關系數為0.858,呈強相關性(r=0.858,P=0.000),斑馬魚鱗片覆蓋率與日齡相關系數為0.603,呈弱相關性(r=0.603,P=0.000)。
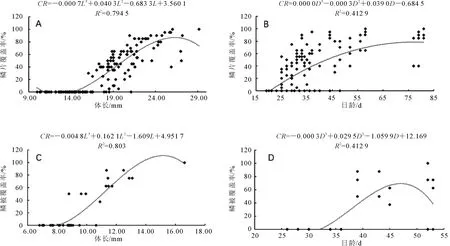
圖7 團頭魴(A和B)和斑馬魚(C和D)稚魚鱗片的覆蓋率與體長、日齡的關系Fig.7 Variation in areal coverage of scales by body length and age during development in juvenile M.amblycephala(A and B)and D.rerio(C and D)
3 討論
3.1 魚類鱗片發育的起始位置和數量
魚類鱗片的發生發育是一個非常復雜的過程,不同魚類的鱗片發育模式不盡相同。本研究結果顯示,團頭魴鱗片最先起始于鰓蓋后緣側線處,這種鱗片起始位置與鯉科魚類的草魚(Ctenopharyngodonidellus)[11]、鰱(Hypophthalmichthysmolitrix)[11]、鯉(Cyprinuscarpio)[11]、鳙(Aristichthysnobilis)[11]、重口裂腹魚(Schizothoraxdavidi)[19]和齊口裂腹魚(S.prenanti)[20]的鱗片起始位置一致。而斑馬魚的鱗片起始位置與團頭魴的鱗片起始位置明顯不同,其為尾柄側線處,這與小孔亞口魚(Minytremamelanops)[8]、尼羅羅非魚(Oreochromisniloticus)[11]、莫桑比克羅非魚(O.mossambicus)[21]、奧克蘭鰻鱺(Anguillaaustralis)[22]、大鰻鱺(A.dieffenbachii)[22]的鱗片起始位置一致。此外有些魚類鱗片起始位置并不屬于以上兩種類型,如梭魚(MugilsoiuyBasilewsky)[5]、河川沙塘鱧(Odontobutispotamophila)[23]、暗斑棘蓋太陽魚(Pomoxisnigromaeulatus)[24]鱗片最早出現于尾柄側中央至體側中央的連線上,又如烏鱧(Channaagrus)[25]出現在頭頂部,條石鯛(Oplegnathusfasciatus)[26]出現在鰓蓋后緣至胸鰭基部,黑鯛(Sparusmacroeephlus)[27]出現在胸鰭基部至肛門直上方的側線處。本實驗結果顯示,團頭魴和斑馬魚鱗片起始中心數量均只有1個起始中心。與草魚、鰱、鯉、鳙、尼羅羅非魚[11]、莫桑比克羅非魚[21]、河川沙塘鱧(O.potamophila)[23]、小孔亞口魚[8]、奧克蘭鰻鱺[22]、大鰻鱺[22]、黑鯛[27]鱗片起始中心數量一致。澳大利亞虎鯊(Heterodontusportusjacksoni)[28]有2個,梭魚[5]、暗斑棘蓋太陽魚[24]、條石鯛[26]有3個,烏鱧[22]有4個,重口裂腹魚[19]和齊口裂腹魚[20]有6個。
由此可見魚類鱗片的起始位置以及起始中心數量在不同魚類中差異明顯,即便是同科的團頭魴和斑馬魚的鱗片起始位置和覆蓋方式均有很大差異。嚴太明等[19-20]指出魚類鱗片起始位置在物種間差異較大,具有種屬特異性;但是鱗片起始位點的個數,不同物種差異較大,規律性不強。
3.2 魚類鱗片的覆蓋與生長
利用鱗片覆蓋率來描述鱗片覆蓋的程度。對于鱗片較小、數目不清楚的魚類,如鰱、鯉、烏鯉、鳙等,習慣用鱗片覆蓋面積來進行覆蓋率的描述。但對于鱗片較大、數目清楚的魚類,如草魚、梭魚等,經常用鱗片出現的排數來描述覆蓋程度。本研究中的實驗對象團頭魴和斑馬魚的鱗片均清晰明了,故采用后一種方法來對鱗片的覆蓋程度進行描述比較。
諸多研究結果表明,魚類鱗片的發生與魚類的生長存在密切聯系。本實驗中團頭魴、斑馬魚的體長和日齡與鱗片覆蓋率均有重要影響,但相比之下體長與鱗片覆蓋率的相關性更強。這與已有研究結果一致。李思發[11]對鯉、草魚、鰱、鳙和尼羅羅非魚的研究結果表明,這幾種魚類的鱗片發育主要決定于體長,體長越長,鱗片越多。對重口裂腹魚[19]、齊口裂腹魚[20]鱗片的研究結果表明,體長和日齡對鱗片的覆蓋率均具有重要影響。這些研究結果說明了魚類鱗片發育過程可能與體長、日齡有一定關聯。在魚類的生長發育過程中,環境因子存在不可忽視的作用,同樣作為魚類生長發育的一部分,鱗片的形成也受到環境因素的影響[26],比如餌料、水溫、密度大小等。
3.3 鱗片發生組織學研究
本實驗通過對團頭魴、斑馬魚皮膚切片染色的觀察發現。兩者與其他真骨魚類骨鱗的發生并無很大差別,均要經歷真皮基質的形成,鱗片的乳突的出現、鱗囊的分化和基質沉積的過程[2,19]。這表明,不同魚類鱗片發生的起始位置、覆蓋方式差異明顯,但鱗片發育的組織學過程基本一致。由于魚類真皮和表皮中富含結締組織和膠原纖維,本研究中對皮膚組織切片不僅運用了傳統的HE染色,還運用了對膠原纖維具有良好顯色效果的Masson染色。通過兩種顯色結果的比較,從而更準確地觀察到團頭魴和斑馬魚鱗片發生的組織學過程。這也為以后利用模式生物從分子水平研究魚類鱗片發生機制奠定了基礎。
