鰱早期發育對過飽和總溶解氣體響應與仔魚耐受性的研究
2019-01-22 02:30:54楊傳順吳湘香杜開開茹輝軍倪朝輝李云峰
淡水漁業 2019年1期
楊傳順,吳湘香,杜開開,柳 凌,茹輝軍,張 燕,倪朝輝,李云峰
(1.上海海洋大學,上海 201306;2.中國水產科學研究院長江水產研究所,農業部長江中上游漁業生態環境監測中心,武漢 430223)
高壩泄水時,大量氣體被卷入水體深處使水體中的溶解氣體(主要包括溶解氧(DO)、溶解氮(DN))過飽和,稱為水體總溶解氣體(TDG)過飽和[1]。這些過飽和水體流入下游河道后,沿程并不會很快釋放出來[2],使得生活在其中的水生生物長期處于溶解氣體過飽和的環境中,導致魚類患氣泡病,影響魚類生理及繁殖[3]。
國外關于TDG過飽和水體對魚類影響的研究開展較早[4,5]。早在1905年Marsh和Gorham就第一次具體地描述了氣栓在魚類血液循環系統的形成過程和危害程度[6]。20世紀60年代,哥倫比亞斯內克河(Columbia and Snake River)修建了大量蓄水和發電的高壩,在汛期高速下泄的河水卷入大量的氣體,導致大壩下游溶解性氣體達到過飽和狀態,造成了魚類尤其是大馬哈魚、虹鱒大量死亡[7-9]。Weitkamp等[10]研究發現在1997—2000年汛期期間,Clark Fork河下游TDG飽和度最高達到150%,導致當地魚類患上嚴重的氣泡病。還有研究表明魚類在不同的生長階段對氣體過飽和的忍耐性是不同的[11-14]。
在國內,隨著長江上游河流梯級開發,高壩密集建設,水體溶解氣體過飽和問題越來越嚴重,TDG過飽和水體嚴重威脅到了壩下游水生生物尤其是魚類的生存[2]。針對長江中上游TDG過飽和對魚類的影響,國內開展了一系列[15-20]研究。董杰英等[21]發現四大家魚暴露在溶解氣體過飽和的環境中容易出現眼球突出、鰭充血和身上出現氣泡等一系列氣泡病癥狀。梁瑞峰等[22]研究發現孵化率會隨總溶解氣體飽和度的升高而降低,較高的總溶解氣體飽和度能加快其孵化速率。長江中上游水電工程開發所帶來的TDG過飽和問題,不僅使長江主要經濟魚類及珍稀保護魚類的生存面臨挑戰,而且也對魚類的早期資源量產生嚴重的影響。本研究以中上游主要經濟魚類——鰱(Hypophthalmichthysmolitrix)為對象,通過研究TDG過飽和水體對鰱早期生活史的影響及耐受性,以豐富TDG過飽和水體對魚類早期發育影響的資料,同時也為高壩下游生態保護措施的制定提供借鑒。
1 材料與方法
1.1 實驗材料來源
實驗用鰱受精卵取自中國水產科學研究院長江水產研究所窯灣試驗場,從產卵池的集卵箱一次采集3 000~5 000粒卵,放入盛有產卵池水的塑料箱中,隨即轉運至實驗室的養魚池中,準備實驗用。
實驗用幼魚同樣取自長江水產研究所窯灣試驗場,分別在鰱水花下塘后第15、25、35、45和120天的魚種塘拉網獲得。每次取5 000尾左右的幼魚迅速轉運至實驗室的水泥池(水體約2 m3)中暫養。暫養水是經過曝氣的井水,水溫為18.2~21.4 ℃,溶解氧(DO)濃度為7 mg /L。暫養3 d后挑選出規格一致、體表完整、體質健康個體準備實驗。
1.2 不同濃度TDG水的制備
實驗裝置包括模擬高壩泄水生成TDG過飽和水的試驗系統(專利號ZL 2017 2 0786771.7)如圖1,主要包括水泵、蓄水池、壓力罐、空壓機、管道等部件。
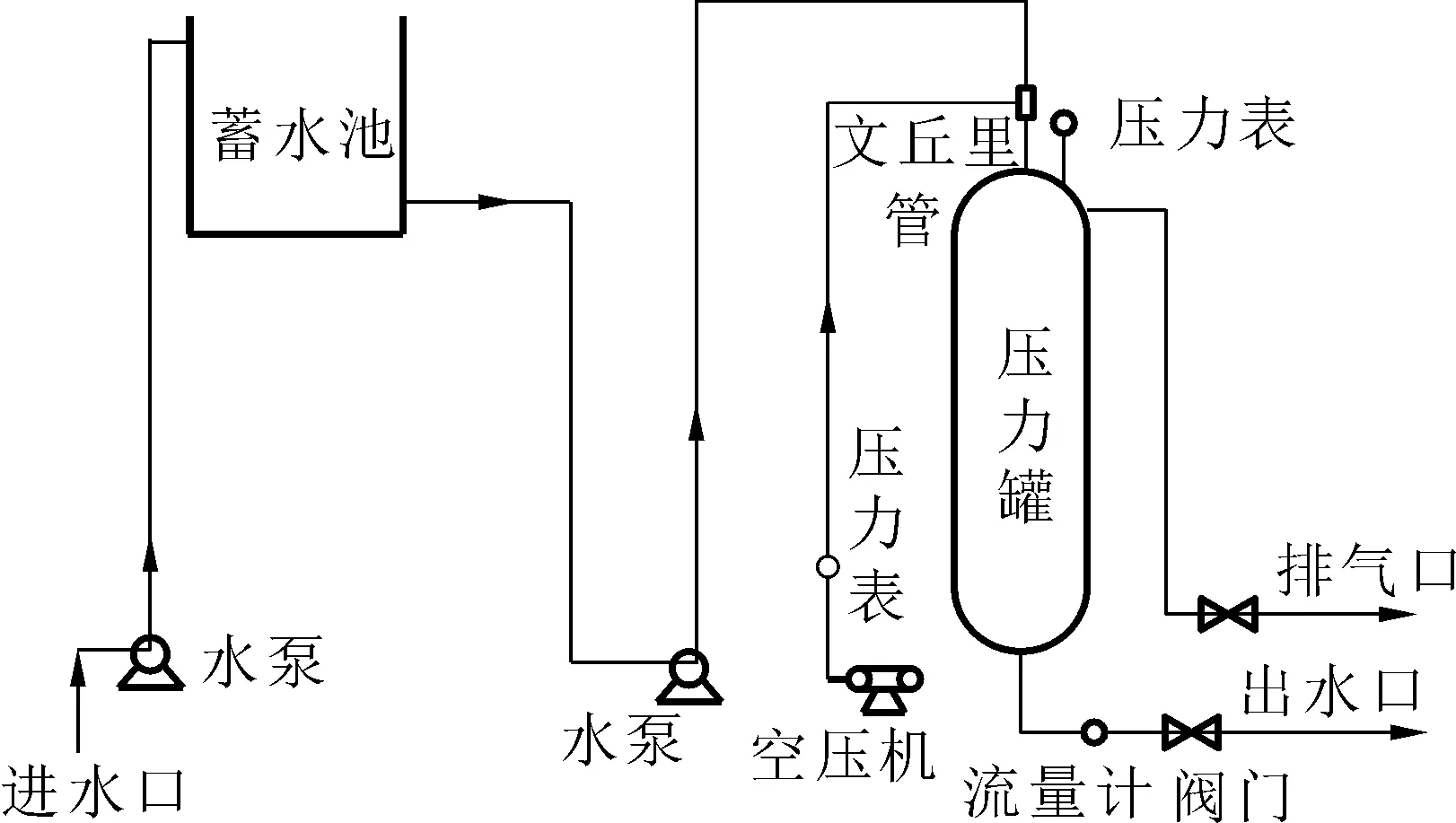
圖1 過飽和水發生裝置Fig.1 Schematic of an experimental system
過飽和水的產生:空壓機向壓力罐中打入一定量的空氣后,打開水泵向壓力罐中泵入一定量的水,泵入的水通過文丘里管使水流分流,增大水體與氣體的接觸面積,使水體快速與空氣混合。進水量一定時,通過調節出水口出水量的大小保持壓力罐罐內一定的壓強,產生高飽和度的水體,將生成的高濃度過飽和水體與正常水通過控制流量以不同比例混合,可同時產生不同飽和度的水。實驗過程中,保持魚卵塑料箱或者實驗池的水體積一定,以一定的速率向魚卵塑料箱或者實驗池加入高飽和度的原水且等流量的溢出相同體積的水體,以此維持飽和度相對穩定,實驗用水是井水,溫度基本穩定。實驗過程中每隔2 h用TDG儀器進行測量和記錄,并根據測量的情況隨時對飽和度進行及時調節,保證實驗飽和度的相對穩定。本實驗過程中TDG飽和度的測定選用丹麥Oxyguard公司生產的Handy Polaris TGP(Total Dissolved Gas Pressure)測定儀,其量程為0~200%,其精度為±1%。
1.3 實驗方法
1.3.1 受精卵孵化實驗
將準備好的鰱受精卵分別放入加有不同TDG飽和度水體的塑料箱中,箱底部有一個直徑20 mm的進水口,箱頂部有兩個溢水口,每個塑料箱約含0.25m3的水體,放入約300粒左右的受精卵。試驗飽和度分別設定為110%,120%,130%和140% 4個試驗組,每個試驗組4組平行。對照組用水采用調制的井水,飽和度為80%。試驗期間的孵化水溫控制在(21±0.5)℃。
試驗開始后,按事先設定好的不同發育時期,即原腸晚期、脊索形成期、心跳期和尾芽期,從各試驗組中隨機挑選20粒左右的受精卵置于10倍的體視鏡(型號SZ61,生產廠家奧林巴斯)下觀察胚胎發育情況,記錄被觀察卵的數目和畸形數。孵化約35 h后,每隔0.5 h,觀察各試驗組和對照組的出膜情況,記錄出膜幼苗數和出膜時間。
待出膜完成后,對各試驗組和對照組換一次水,同時挑出死卵,繼續觀察幼苗的胚后發育情況。每天統計各組的死亡數量并記錄相應的死亡時間,直到仔魚平游之前。
1.3.2 不同階段的幼魚暴露實驗
將暫養3 d,經過挑選的試驗用幼魚,放入架設在灌有不同濃度TDG水的不銹鋼圈支撐的圓柱形網箱(直徑100 cm、高50 cm)中。15、25、35、45 d發育的鰱幼魚每個圓柱形網內放入200尾左右,120 d發育的鰱幼魚每個圓柱形網內放入50尾左右。每個飽和度組設4個平行組,水溫(21±0.5)℃。連續觀察96 h,記錄各試驗組中魚的活動及死亡情況,若出現死亡,記錄死亡時間和死亡數量。
1.4 數據整理
實驗數據采用SPSS21軟件進行方差分析,并采用Duncan多重比較進行差異顯著性分析(P<0.05)。
2 結果
2.1 過飽和水體對鰱受精卵孵化的影響
不同TDG飽和度下對鰱受精卵孵化的影響結果見表1。從表1中可以看出,各試驗組在發育過程中均有一定程度的畸形率,與對照組比較差異不顯著。出膜率最高是過飽和110%試驗組,其平均出膜率達到了92.3%,與對照組比較存在顯著差異。
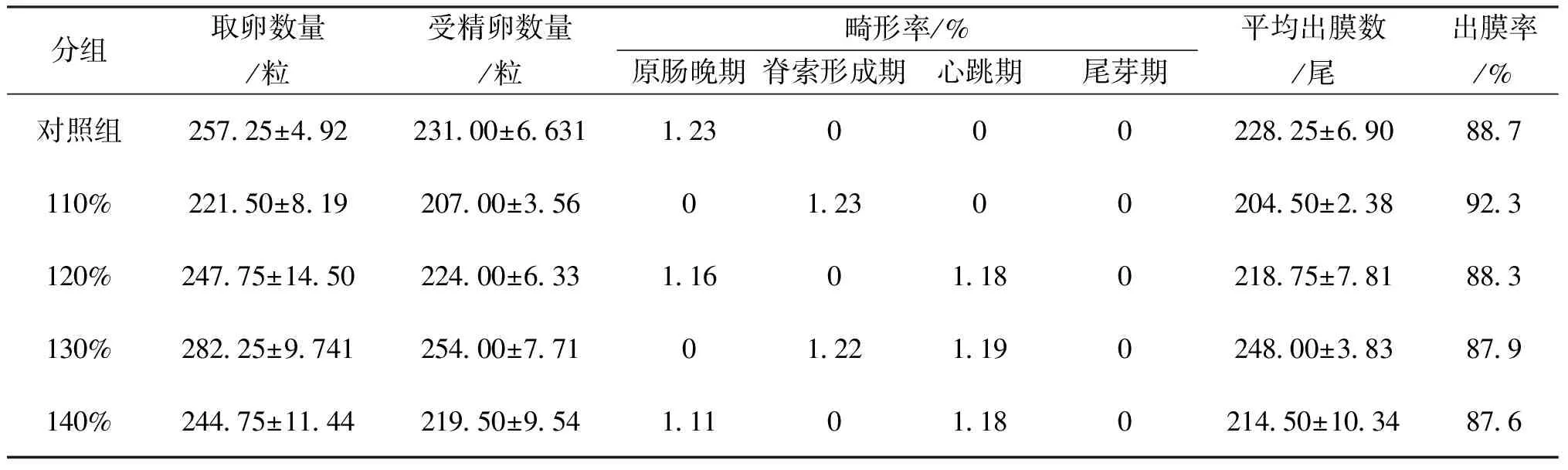
表1 不同TDG飽和度下鰱受精卵孵化的影響Tab.1 Effect of various TDG supersaturation levels on silver carp fertilized eggs incubationin different experimental groups
受精卵孵化的出膜時間和數量統計結果見圖2。圖2顯示,(21±0.5)℃溫度下,試驗組受精卵出膜峰值時間較對照組出現不同程度的提前,但過飽和110%試驗組出膜時間最早,出現在38.5 h左右,而同一溫度下,對照組出現在41.5 h左右,隨著試驗過飽和濃度升高,受精卵出膜的峰值時間逐漸接近對照組。
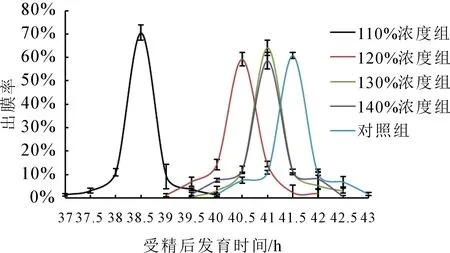
圖2 不同TDG飽和度下鰱受精卵出膜率與時間的關系Fig.2 Relationship between hatching rate and exposure time
對出膜后的幼體進行胚后發育觀察的結果見表2。從表2可看出,在(21±0.5)℃溫度下,無論試驗組還是對照組都經歷了5 d左右的胚后發育,發育時間和發育速度均無明顯差異。
從表2可看出,試驗組存活率均比對照組低,存在顯著性差異,130%試驗組與140%試驗組沒有顯著性差異。試驗組120%、130%和140%的最后存活率高于試驗組110%,且隨著飽和度的增加,存活率會稍微升高。各試驗組幼苗的死亡主要集中在出膜后第2~3天。110%試驗組在出膜第2天的平均死亡率為13.6%,第3天的平均死亡率為13.7%,均明顯高于其他組。

表2 不同TDG飽和度對鰱幼苗的影響Tab.2 Effect of various TDG supersaturation levels on silver carp fries in different experimental groups
注:n=4,凡有相同字母的表示差異性不顯著,反之則表示差異顯著。
上述研究結果表明水體的過飽和對鰱受精卵的發育速度和畸形率沒有明顯的影響,但110%試驗組顯著提高了受精卵的出膜率,同時提早了受精卵的出膜時間。其它試驗組雖然在出膜時間和出膜率上與對照組比較有一定的差異,但無顯著性差異,因此表明高飽和度的水體對受精卵的出膜率和出膜時間均無影響。對胚后發育的觀察發現,與對照組比較各種飽和度的水體均能導致胚后發育幼苗的存活率降低,幼苗死亡的時間主要集中在出膜后第2天和第3天。
2.2 不同生長階段的鰱幼魚對過飽和氣體的耐受性研究
不同生長階段的鰱幼魚對不同TDG飽和度耐受性的結果見圖3。從圖3可以看出,不同生長階段的鰱幼魚對不同飽和度水體的耐受性存在差異。圖3(a)和(b)顯示了15和25 d的鰱幼魚在不同飽和度水體中的耐受性。結果表明15和25 d鰱幼魚在各試驗飽和度水體中死亡率很低,與對照組幾乎無差異。96 h后15和25 d的鰱幼魚總死亡率分別為2.5%和5.0%。因此,不同飽和度水體對15和25 d的鰱幼魚存活率幾乎無影響。圖3(c)顯示了35 d鰱幼魚在不同飽和度水體中的耐受性。結果表明110%試驗組中鰱幼苗的總死亡率與對照組比較無顯著性差異。但120%,130%和140%試驗組中鰱幼苗均出現了較高的死亡率,且TDG飽和度越高,死亡率也越高,96 h后3個試驗組的總死亡率分別為19.1%、21.0%和25.0%。由此可見,對于生長35 d的幼苗TDG飽和度越高,死亡率也越高,最高可達到25.0%。圖3(d)和(e)顯示了45 d和120 d鰱幼魚在不同飽和度水體中的耐受性。從圖中可以看出,隨著鰱幼苗在TDG過飽和水體中暴露時間的延長,其死亡率也會相應增加,并且隨著TDG飽和度的增加,死亡率相應增加。96 h后兩個實驗組總死亡率為60%~100%。
綜上所述,鰱幼苗在生長25 d前,對TDG不敏感,幾乎不受TDG飽和度影響。生長到35 d時,低飽和度的TDG對幼苗仍然沒有影響,但120%以上的高飽和度TDG對幼苗會產生一定程度的影響,導致幼苗死亡率隨TDG飽和度升高而增加。45 d后到120 d的鰱幼苗,對TDG過飽和水體非常敏感,各飽和度的TDG均能導致鰱幼苗迅速死亡,但各飽和度的TDG對鰱幼苗死亡率的影響有一定差異。幼苗死亡均發生在暴露7 h左右,暴露96 h后,45 d的死亡率在60%~81%,120 d的死亡率為86%~100%。
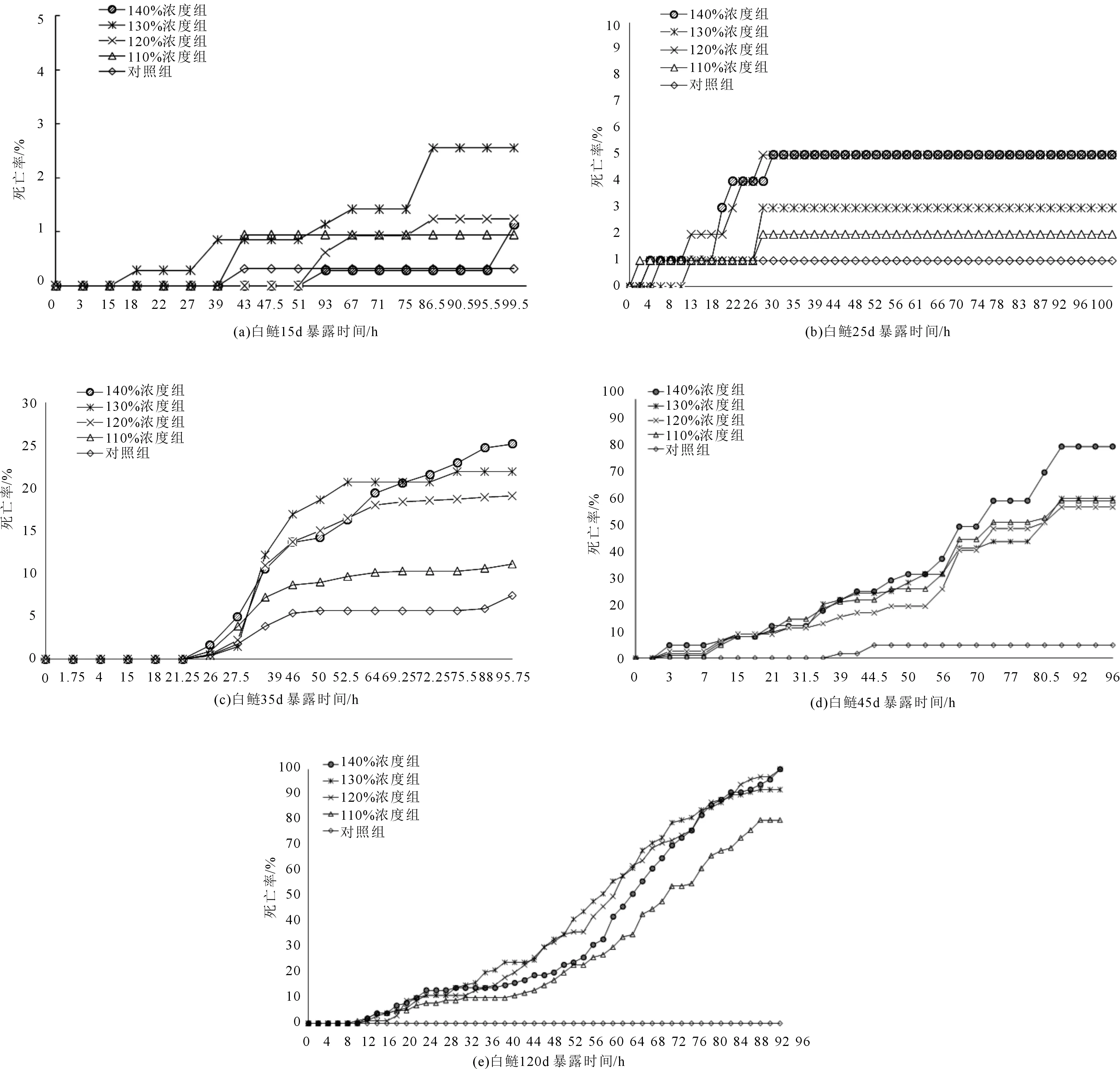
圖3 不同階段鰱暴露時間和死亡率關系(a-e分別表示:15、25、35、45、120 d)Fig.3 Relationship between exposure time and mortality in different stages of silver carp
3 討論
3.1 TDG對鰱受精卵孵化的影響分析
有學者研究認為TDG在一定飽和度下對有些魚類的受精卵發育是有影響的,Meekin等[12]發現當氣體飽和度達到112%時,在同一個孵化場內虹鱒魚卵發生了嚴重的死亡,但大鱗大馬哈魚魚卵卻沒有受到影響。郝思平等[23]也發現,溶氧量越高,在某些情況下,孵化期內胚胎死亡率下降[24]。但Marsh等[11]和許多學者均報道了過飽和水體對魚卵的孵化不會產生影響且不會導致氣泡病的發生。本研究結果表明,溫度穩定的情況下,水體TDG過飽和對鰱受精卵的發育速度和畸形率沒有顯著影響,且沒有發現受精卵得氣泡病。魚的胚胎發育是一個耗氧過程,有充足的氧可以保證胚胎的正常發育,有利于孵化率的提高。本研究的結果與Marsh等[11]的研究結果基本一致。
本研究發現,當TDG飽和度在110%時,對受精卵的出膜率和出膜時間均有顯著性的影響,它有效提高了受精卵的出膜率和出膜時間。Oppenberntsen等[25]試驗表明低氧環境下,大西洋鮭胚胎的孵化時間比高氧環境情況下短。Rombough[26]研究也認為當受精卵內氧氣供應不足時,仔魚迫于氧氣的需求提前孵化而保證生存,但畸形率會提高,導致孵化率下降。因此,推測110%試驗組的提前出膜與受精卵內低氧環境有關。由于過飽和氣體極易在受精卵膜表面吸附一層空氣泡,部分阻斷了卵內外的水體交換,使卵內的氧氣含量降低,造成受精卵處在一個低氧環境,從而使受精卵提早出膜。但是提早出膜的鰱幼體缺乏保護,極易受到損傷,因而各種TDG飽和度中的幼體存活率與對照組比較均降低了,且出膜越早存活率越低。這個結果與Rombough[26]的研究結果是一致的。
3.2 不同生長階段的鰱幼魚對過飽和氣體的耐受性分析
已有研究表明魚類在不同的生活階段對于氣體過飽和的忍耐性是不同的[11-14],Meekin等[12]報道隨著大鱗大馬哈魚年齡的增長,對于溶解性氣體的忍耐性卻逐漸降低,死亡時間也逐漸縮短。Rucker[13]用不同體長的銀大馬哈魚進行了實驗研究,結果顯示體長小(40 mm)的魚比體長大(53 mm、67 mm)的魚對于氣體過飽和水體具有更強的耐受性。Counihan等[14]進行了高首鱘在個體發育過程的不同階段氣泡病發生的研究,結果表明孵化后6 d內,發育時間越長,對氣體飽和度的耐受性越弱。梁瑞峰等[22]研究了過飽和TDG對重口裂腹魚生長、繁殖的影響,結果表明孵化后相同發育日齡的重口裂腹魚對過飽和水體的耐受性隨著飽和度的增加而降低。Smiley等[27]發現,暴露在122%飽和水體中,成齡鱸在23 ℃和18 ℃水體中的死亡率比幼齡鱸魚高20%。
本研究中鰱幼苗在生長25 d前,對TDG不敏感,幾乎不受任何飽和度TDG影響。生長到35 d時,低飽和度的TDG對幼苗仍然沒有影響,但120%以上的高飽和度的TDG對幼苗產生了一定程度的影響,導致幼苗死亡率隨TDG飽和度升高而增加,跟梁瑞峰等[22]研究的結果基本相似。45 d后到120 d的鰱幼苗,對各種飽和度的TDG都非常敏感,各種飽和度的TDG均能導致鰱幼苗迅速死亡,且飽和度越高死亡率也越高。這些研究結果與梁瑞峰等[22]、Smiley等[27]的研究結果基本一致。
分析認為早期幼苗在各種飽和度水體中的死亡率都比較低,隨著個體發育各個飽和度的死亡率開始升高,這可能由于魚類發育過程中呼吸方式的改變所致。魚類的呼吸方式主要包括水呼吸和氣呼吸,是魚類對生活環境和生活方式的適應。魚類的呼吸器官有主要呼吸器官和輔助呼吸器官之分,前者是鰓,絕大部分魚類通過鰓進行水呼吸,但一些魚類為了適應特殊的生活條件還通過皮膚、鰓上器官、腸管、口腔黏膜、鰾、氣囊等輔助呼吸器官進行氣呼吸[28,29]。魚類的呼吸器官在仔魚孵化后一段時間才發育完善,在此之前,氣體交換通過體表皮膚、鰭褶和卵黃囊上豐富的血管直接與水接觸來進行[29]。早期階段魚類氣體通過體表皮膚、鰭褶交換,這些組織可能對TDG不敏感,因此降低了TDG對其本身的傷害。隨著發育時間的延長,呼吸方式開始改變,其對TDG的敏感度不斷上升。Hughes[30]發現,活躍魚類的鰓絲數、鰓絲長、鰓小片密度大于中等活動性魚類和活動遲鈍魚類,但活躍性魚類的單個鰓小片面積小于緩慢性魚類,這可能是因為活躍性魚類代謝速率和能量需求較高,需要較大的交換面積,但較小的鰓小片對水的阻力較小,符合其對快速活動的要求。同種但不同規格魚類的呼吸面積也有所不同,一般鰓絲數量、長度、單個鰓小片面積、總呼吸面積隨體質量的增加而增大,鰓小片密度、相對呼吸面積隨體質量的增加而減小,其中單側鰓絲數、總鰓絲數與體質量呈對數關系,鰓小片數、單個鰓小片面積、總呼吸面積及相對呼吸面積與體質量呈冪指數關系[31,32]。從生活環境看,鰱喜居于水的中上層,是屬于活躍魚類。隨著其發育的時間的增長,呼吸器官不斷完善,呼吸方式慢慢轉變。當其基本發育完全時,由于其鰓的構造等導致其氣體交換比較快,對于高壓下的高溶解氣體環境產生強烈不適應,因而表現出對氣體過飽和水體較為敏感,導致其隨著發育時間的延長,在過飽和水體中的死亡率升高。
