紫花苜蓿高溫誘導啟動子pMsMBF1c的克隆與功能分析
2019-01-23 00:57:08李小冬莫本田牟瓊婁芬陳文貴陳光吉張瑜韓永芬
草業學報 2019年1期
關鍵詞:植物
李小冬,莫本田,牟瓊,婁芬,陳文貴,陳光吉,張瑜,韓永芬*
(1.貴州省農業科學院草業研究所,貴州 貴陽 550006;2.貴州金農富平生態農牧科技有限公司,貴州 松桃 554100;3.大方縣農牧局,貴州 大方 551600)
高等植物的基因行使其功能受到復雜而精細的調控,按照調控時間的先后順序可分為轉錄前調控、轉錄調控、轉錄后調控、翻譯調控和翻譯后調控幾個主要階段[1]。啟動子是基因轉錄前調控的主要元件,通過對下游基因表達時期與表達量的調節,決定植物完成特殊生長發育階段的轉變或應答特殊環境脅迫[2]。
按照轉錄啟動方式可以將啟動子分為組成型、組織特異型和誘導型啟動子[3],但彼此之間可能有重疊,例如,果實特異啟動子同時也有可能是乙烯誘導型啟動子,因為其上游區域可能存在多個順式作用元件,能夠與多種不同的反式作用因子相互作用[4],特異性啟動子與誘導型啟動子能根據植物特殊生長發育階段或特殊環境應激啟動相應基因表達,能夠實現精準高效的調控,在基因功能研究與農業分子育種中有巨大的應用潛力,目前研究較多的有光誘導[5-6]、溫度誘導[7-9]、干旱誘導[10]、激素誘導[11]、傷害誘導型啟動子[12]等,并且已經成功的在煙草(Nicotianatabacum)、玉米(Zeamays)、水稻(Oryzasativa)等農作物中培育了誘導型啟動子驅動的抗逆種質資源。
熱激轉錄因子Hsf與熱激蛋白Hsp是植物中重要的高溫誘導基因,其啟動子結構比較保守,在模式植物擬南芥(Arabidopsisthaliana)中,熱激轉錄因子Hsf與其下游基因Hsp啟動子區都存在HSE模體(AGAAnnTTCT),并且21個Hsf中除HsfA4c和HsfA8外,其余都被報道參與抗熱反應[13]。多橋蛋白因子(MBF1)是動植物中高度保守的轉錄共激活子,在植物中,MBF1c參與高溫脅迫應答,并顯著受熱誘導[14]。MBF1c蛋白具有DNA結合的能力,能夠調節DREB2A、HsfA3、HsfB2a以及HsfB2b等基因的表達[14-15],并且其抗熱反應受水楊酸和乙烯等多條激素途徑調節,是連接Hsf-Hsp途徑和激素途徑的中樞基因,然而關于MBF1c啟動子的研究報道還很少。研究并開發利用其啟動子為豐富高溫誘導型啟動子庫具有重要的理論研究與實際應用價值。
1 材料與方法
1.1 材料
本研究采用的紫花苜蓿材料為“中苜1號”(Medicagosativacv.Zhongmu 1#),由中國農業科學院北京畜牧獸醫研究所高洪文團隊饋贈),擬南芥為哥倫比亞型野生型,擬南芥與紫花苜蓿種植條件參考Li等[16]的方法,植物生長溫度為22 ℃,相對濕度為60%,光周期為16 h光照/8 h黑暗,光照強度為230~300 μE·m-2·s-1,在人工智能氣候室中進行,試驗時間為2014-2017年。
1.2 方法
1.2.1紫花苜蓿DNA提取 稱取1 g新鮮“中苜1號”紫花苜蓿葉片,用液氮研磨成粉末,加入1 mL 65 ℃預熱的2% CTAB提取緩沖液(2% CTAB,20 mmol·L-1EDTA,100 mmol·L-1Tris-HCl, 1.4 mol·L-1NaCl),65 ℃孵育1 h,12000 r·min-1離心10 min,將800 μL上清轉移到新離心管;加入200 μL氯仿,震蕩萃取30 min,12000 r·min-1離心10 min,將600 μL上清液轉移到新離心管;加入1 mL預冷的無水乙醇,-20 ℃孵育1 h,12000 r·min-1離心10 min,加入700 μL 75%的乙醇,10000 r·min-1離心5 min,棄上清,通風廚中瀝干沉淀,加入50 μL dd H2O溶解備用。
1.2.2紫花苜蓿MsMBF1c啟動子序列的分離克隆 采用基于巢式PCR的酶切連接技術分離MsMBF1c啟動子序列,具體操作如下:1) 合成接頭引物:AF 5′-GTAATACGACTCACTATAGGGCACGCGTGGT-3′,AER (EcoRⅠ) 5′-AATTACCACGCGTGCTG-3′,AHR (HindⅢ) 5′-AGCTACCACGCGTGCTG-3′, ABR (BamHⅠ/BglⅡ) 5′-GATCACCACGCGTGCTG-3′,AXR (XhoⅠ) 5′-TCGAACCACGCGTGCTG-3′,APR (blunt-ended) 5′-ACCACGCGTGCTG-3′; 2) 制備接頭:在PCR儀中進行,采用梯度變性退火方法制備,每一種內切酶對應一種接頭,體系如下:取AF (10 μmol·L-1) 10 μL,A*R (10 μmol·L-1) 10 μL (A*R指AER、AHR、ABR、AXR、APR等5條反向引物中的一條),混合均勻,進行接頭制備反應,反應程序如下:94 ℃ 3 min,70 ℃ 10 min,60 ℃ 10 min,50 ℃ 10 min,30 ℃ 10 min,25 ℃ 10 min,結束連接,接頭制備完成。
采用EcoRⅠ、BamHⅠ、HindⅢ、XhoⅠ、SmaⅠ (Fermentas)分別酶切苜蓿基因組DNA,酶切反應體系如下:紫花苜蓿基因組DNA 2 μL,10×酶切反應緩沖液1 μL,限制性內切酶 0.5 μL,dd H2O 6.5 μL,37 ℃酶切0.5~2.0 h。酶切反應完后進行連接反應,體系如下:酶切反應樣品 10 μL,連接接頭1 μL,10×連接反應緩沖液 2 μL,T4 DNA連接酶 1 μL,dd H2O 6 μL,4 ℃連接16 h。
巢式PCR擴增,引物序列如下:AD1,5′-GTAATACGACTCACTATAGGGC-3′,AD2,5′-ACTATAGGGCACGCGTGGT-3′,MBFR1,5′-TAGTTTTAGGGATTAGGAGC-3′,MBFR2,5′-GTTGACGGCTT TCTCATCTT-3′,MBFR3,5′-TTCTTGCGGATAACGACTGG-3′。以稀釋20~50倍的酶切連接產物為模板,以AD1與MBFR1為引物,配置PCR反應混合物(上海生工 2×Taq PCR反應試劑盒),反應體系如下: 2×Taq PCR Mixture 10 μL,AD1 (10 μmol·L-1) 1 μL, MBFR1 (10 μmol·L-1) 1 μL,模板 2 μL,dd H2O 6 μL。反應程序為:94 ℃ 3 min,94 ℃ 30 s,72 ℃ 3 min,7個循環,94 ℃ 30 s,68 ℃ 3 min,32個循環,72 ℃ 5 min,25 ℃ 5 min,反應結束。以稀釋50~100倍的第一輪PCR產物為模板,進行第二輪巢式PCR擴增,引物組合為AD2與MBFR2,反應程序為:94 ℃ 3 min,94 ℃ 30 s,72 ℃ 3 min,5個循環,94 ℃ 30 s,68 ℃ 3 min,25個循環,72 ℃ 5 min,25 ℃ 5 min,反應結束。第三輪巢式PCR引物組合為AD2與MBFR3,反應體系與程序與第二輪相同。
分別取第二輪與第三輪巢式PCR產物進行水平膠電泳,將第二輪與第三輪都有明顯擴增,且第三輪產物略小于(43 bp)第二輪的條帶進行回收(上海生工膠回收試劑盒),參照試劑盒說明書,得到的序列即為MsMBF1c側翼序列的候選序列。
1.2.3pBI21-pMsMBF1c::GUS雙元載體的構建 利用巢式PCR測序的結果設計引物:pMBF1cF 5′-AAGCTTAATGAATTTAAGTGG-3′,pMBF1cR 5′-GTTTATCTCTTTGGTTATG-3′,采用phusion DNA聚合酶(Thermo)進行擴增,反應體系如下:10×buffer,2 μL;dNTP mix,0.5 μL;MgCl2,1.5 μL;pMBF1cF,1 μL;pMBF1cR,1 μL;DNA模板,2 μL;DNA Polymerase,0.2 μL;dd H2O,12 μL。程序如下:95 ℃ 1 min,95 ℃ 10 s,58 ℃ 10 s,72 ℃ 1 min,30個循環,72 ℃ 5 min,25 ℃ 5 min。將回收產物連接到pEASY-Blunt克隆載體(pEASY-Blunt Cloning Kit,Transgene)并進行測序分析,提取pEASY-MsMBF1c載體質粒,與pBI121載體一起,分別利用HindⅢ與BamHⅠ雙酶切,回收線性化的pBI121載體片段和MsMBF1c片段, 用T4連接酶(Fermentas)連接成pBI21-pMsMBF1c::GUS雙元載體,具體操作參照試劑盒說明書。
1.2.4RNA的提取及反轉錄 稱取100 mg新鮮“中苜1號”紫花苜蓿葉片或擬南芥葉片,用液氮研磨成粉末,采用TRIZOLTMKit RNA提取試劑(Invitrogen,USA)提取總RNA。采用RevertAid H Minus First Strand cDNA Synthesis Kit (Fermentas) 反轉錄cDNA 第一鏈,參照試劑盒說明進行操作。
1.2.5pBI21-pMsMBF1c::GUS擬南芥轉化與轉基因擬南芥篩選 擬南芥轉化與篩選參考文獻[17],其操作方法如下:將活化的農桿菌懸浮于新鮮配置的5%蔗糖誘導培養基中[10 g 蔗糖充分溶解于200 mL蒸餾水中,在轉化之前加入20 μL Silwet L-77(上海生工)]。去除處于盛花期野生型擬南芥的花和角果,將其浸泡在懸浮菌液中30 s,收取轉基因種子,并利用卡那霉素抗生素篩選,并以卡那霉素抗性基因以及MsMBF1c正向引物與GUS反向進行分子鑒定。具體操作如下:首先將轉基因的種子用75%乙醇表面消毒1 min,用50% 84消毒液消毒3 min,然后用無菌水清洗種子3~4次,將消毒好的種子均勻鋪布于1/2 MS+300 mg·L-1特美汀+ 50 mg·L-1卡那霉素培養基中,在人工氣候室中篩選15~30 d,挑選綠色健壯的植株移栽到營養土中備用。采用天根植物DNA提取試劑盒(DP305)提取轉基因與野生型擬南芥的新鮮葉片組織的DNA,程序方法參照試劑盒說明書。利用NPTF+NPTR和pMBF1cF+GUSR兩個引物組合分別對轉基因植物進行PCR 鑒定(上海生工 2×Taq PCR反應試劑盒),引物序列為NPTF:5′-GAGGCTATTCGGCTATGACTG-3′,NPTR:5′-ATCGGGAGCGGCGATACCGTA-3′,GUSR:5′-GCCCACAGGCCGTCGAGT-3′。反應體系如下:2×Taq PCR Mixture,10 μL;正向引物,1 μL;反向引物,1 μL;DNA,2 μL;dd H2O,6 μL。混勻后進行PCR擴增反應,NPTF+NPTR程序如下:94 ℃ 3 min,94 ℃ 30 s,55 ℃ 30 s,72 ℃ 30 s,30個循環,72 ℃ 5 min,25 ℃ 5 min。pMBF1cF+GUSR程序如下:94 ℃ 3 min,94 ℃ 30 s,58 ℃ 30 s,72 ℃ 2 min,30個循環,72 ℃ 5 min,25 ℃ 5 min。分別取8 μL PCR產物電泳檢測,挑選陽性植株用于表型考察與表達分析。
1.2.6高溫脅迫處理 采用1.2.5表面消毒方法消毒轉基因以及野生型擬南芥,并分別播種到1/2 MS培養基上,室溫生長7 d,對照組一直置于22 ℃培養箱中生長,處理組采用浸入法將培養皿置于42 ℃水浴鍋中熱處理1 h后分別取樣進行GUS染色和熱誘導表達分析,取3個獨立的株系,每個株系處理30個植株。
1.2.7GUS染色分析 將正常與高溫處理之后的轉基因以及野生型擬南芥置于新鮮配置的GUS染色液中37 ℃染色2 h,用75%的乙醇脫色3~5次至背景色為白色,在相同曝光條件下,比較各材料GUS染色強度來判斷啟動子是否受誘導。
1.2.8熒光定量分析 RNA提取及反轉錄參照1.2.4的方法,采用GoTaq Real-Time PCR Systems (Promega,A6001)檢測pMBF1c:GUS與野生型擬南芥中GUS基因以及AtMBF1c基因在正常以及高溫誘導條件下的表達,引物組合為:qGUSF,5′-CGGTCAGTGGCAGTGAAGGG-3′與qGUSR,5′-CGAGGTACGGTAGGAGTTGG-3′,AtMBF1c基因:qAtMBF1cF,5′-TGCCGAGCAGATACCCAGGAGC-3′;qAtMBF1cR,5′-TAACCGTTTGAACCGCGACACC-3′,內參基因為Actin2,引物組合為Actin2 F,5′-AGCGCTGAGGCTGATGATATTCAAC-3′,Actin2 R 5′-TCTAGAAACATTTTCTGTGAACGATTC-3′,操作方法按照試劑盒說明書,反應程序為94 ℃ 2 min,94 ℃ 15 s,58 ℃ 15 s,72 ℃ 30 s,讀取熒光信號,45個循環,72 ℃ 5 min,25 ℃ 5 min。采用2-ΔΔct方法分析GUS和AtMBF1c的表達變化,每個樣品3個生物學重復,每個生物學重復3個技術重復。
1.3 數據分析
采用Excel 2010分析實驗數據,采用t測驗分析表達差異顯著性,閾值設為P<0.05,采用Excel 2010與Power Point 2010作圖。
2 結果與分析
2.1 紫花苜蓿MsMBF1c側翼序列的克隆與分析
紫花苜蓿是草牧業與生物固氮領域的重要農作物之一。其同源種蒺藜苜蓿(Medicagotruncatula)作為豆科牧草的模式植物,基因組已經完成。然而由于紫花苜蓿異源四倍體的基因組特點,其基因組的研究仍相對滯后。本研究前期通過同源克隆的方法從紫花苜蓿基因組克隆獲得MsMBF1c基因,并發現其顯著受高溫誘導,然而其啟動子序列仍未知。借鑒染色體步移技術方法,采用酶切連接的方法利用EcoRⅠ、BamHⅠ、HindⅢ、XhoⅠ、SmaⅠ酶切紫花苜基因組DNA,并用相應接頭連接后形成擴增模板。根據已知的MsMBF1c編碼序列和接頭序列信息設計了2對巢式PCR引物,經過兩輪巢式PCR擴增,獲得MsMBF1c的側翼序列片段,回收在第一輪與第二輪PCR反應中都有擴增,且第二輪PCR產物略小于第一輪產物的片段(圖1),測序分析獲得側翼序列信息。
2.2 MsMBF1c啟動子序列結構與生物信息學分析
利用Sequencer 4.0將分離側翼序列的測序結果與MsMBF1c基因進行比對,獲得一條與MsMBF1c序列有重疊的1748 bp片段,采用PlantCARE軟件(Plant Cis-Acting Regulatory Element, http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)與AGRIS軟件(The Arabidopsis Gene Regulatory Information Server,http://agris-knowledgebase.org/)對該序列結構預測發現,MsMBF1c的啟動子有植物耐熱、抗旱、ABA信號傳導以及環境脅迫緊密相關模體(表1),說明MsMBF1c可能參與植物耐熱等環境脅迫應答。已經將該序列上傳到GeneBank數據庫,其登錄號為:MH760587。
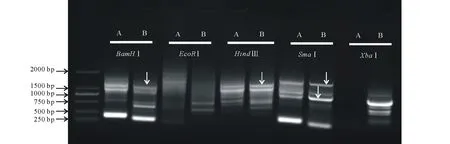
圖1 MsMBF1c啟動子序列的分離Fig.1 Isolation of the promoter sequence of MsMBF1c 每一個橫線指示一種酶切連接方法。A:第一輪巢式PCR電泳結果;B:第二輪巢式PCR電泳結果。箭頭表示回收用于測序分析的候選啟動子片段。Every horizontal line stands for a digest-ligation method. A: The results of electrophoresis of the first round of nested-PCR; B: The results of electrophoresis of the second round of nested-PCR. The arrowhead indicate the potential promoter fragments recycled for sequence analysis.
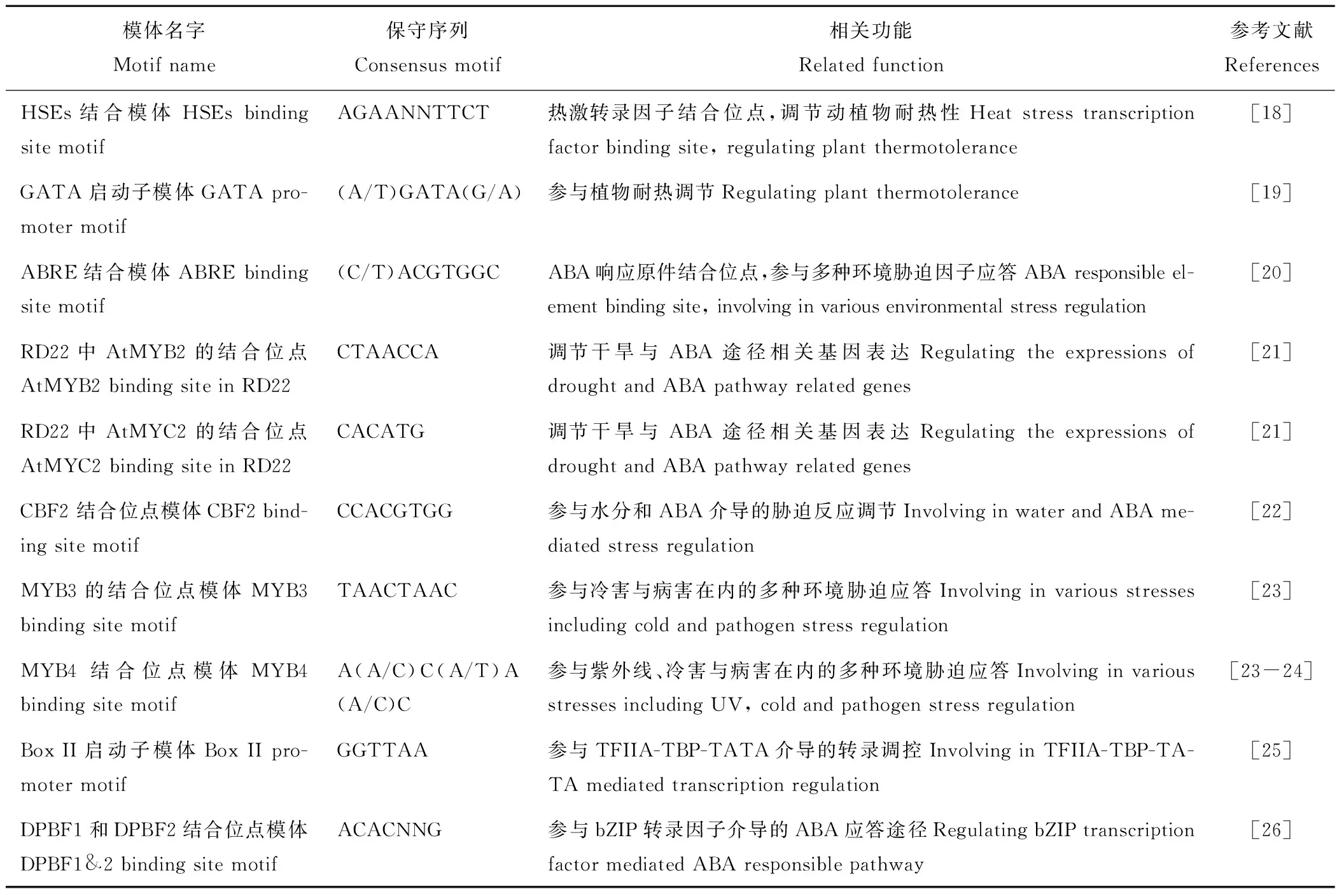
模體名字 Motif name保守序列Consensus motif相關功能Related function參考文獻ReferencesHSEs結合模體HSEs binding site motifAGAANNTTCT熱激轉錄因子結合位點,調節動植物耐熱性Heat stress transcription factor binding site, regulating plant thermotolerance[18]GATA 啟動子模體GATA pro-moter motif(A/T)GATA(G/A)參與植物耐熱調節Regulating plant thermotolerance[19]ABRE結合模體ABRE binding site motif(C/T)ACGTGGCABA響應原件結合位點,參與多種環境脅迫因子應答ABA responsible el-ement binding site, involving in various environmental stress regulation[20]RD22中AtMYB2的結合位點AtMYB2 binding site in RD22CTAACCA調節干旱與ABA途徑相關基因表達Regulating the expressions of drought and ABA pathway related genes[21]RD22中AtMYC2的結合位點AtMYC2 binding site in RD22CACATG調節干旱與ABA途徑相關基因表達Regulating the expressions of drought and ABA pathway related genes[21]CBF2 結合位點模體CBF2 bind-ing site motifCCACGTGG參與水分和ABA介導的脅迫反應調節Involving in water and ABA me-diated stress regulation[22]MYB3的結合位點模體MYB3 binding site motifTAACTAAC參與冷害與病害在內的多種環境脅迫應答Involving in various stresses including cold and pathogen stress regulation[23]MYB4 結合位點模體MYB4 binding site motifA(A/C)C(A/T)A(A/C)C參與紫外線、冷害與病害在內的多種環境脅迫應答Involving in various stresses including UV, cold and pathogen stress regulation[23-24]Box II啟動子模體Box II pro-moter motifGGTTAA參與TFIIA-TBP-TATA介導的轉錄調控Involving in TFIIA-TBP-TA-TA mediated transcription regulation[25]DPBF1和DPBF2結合位點模體DPBF1&2 binding site motifACACNNG參與bZIP轉錄因子介導的ABA應答途徑Regulating bZIP transcription factor mediated ABA responsible pathway[26]
2.3 MsMBF1c啟動子克隆與遺傳轉化
根據測序結果設計引物,克隆獲得MsMBF1c啟動子全長序列,利用HindⅢ與BamHⅠ限制性酶切外源片段與pBI121載體,并進行連接,獲得pBI121-pMsMBF1c::GUS雙元載體。采用農桿菌介導的花器官轉化法將pBI121-pMsMBF1c::GUS雙元載體轉化野生型擬南芥,以50 mg·L-1的卡那霉素進行篩選,共獲得了24株陽性轉基因植株,分別采用抗性基因NPTF+NPTR與基因特異引物pMsMBF1cF+GUSR組合檢測轉基因植株。24株植株用抗性基因檢測呈陽性,其中,20株植株用基因特異引物檢測呈陽性(圖2)。
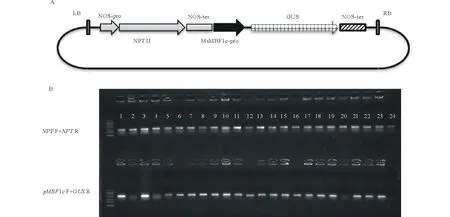
圖2 pBI121-pMsMBF1c::GUS雙元載體構建與轉基因擬南芥分子鑒定Fig.2 pBI121-pMsMBF1c::GUS vector construction and detection of positive transgenic Arabidopsis plants A:pMsMBF1c-GUS報告基因表達載體示意圖;B:轉基因擬南芥分子檢測示意圖,上排為抗性基因檢測(NPT),下排為基因特異性引物檢測(pMBF1c F+GUS R)。A: A diagram showing pBI121-pMsMBF1c::GUS reporter vector; B: Results of molecular detections of transgenic Arabidopsis, antibiotic resistant gene premiers were showed in the upper line (NPT F+NPT R) and the gene specific premiers were showed in the down line (pMBF1c F+GUS R).
2.4 MsMBF1c熱誘導表達分析
為進一步驗證pMsMBF1c啟動子的功能,本研究分析了正常與高溫條件下野生型擬南芥與pBI121-pMsMBF1c::GUS轉基因植株(抗性及特異性分子鑒定為陽性的植株)中GUS基因與內源的AtMBF1c的表達變化。在正常條件下,野生型與pBI121-pMsMBF1c::GUS轉基因植株中AtMBF1c的表達基本相當,而在高溫脅迫后,AtMBF1c基因的表達都顯著被誘導,并且誘導程度相似,分別上升了3.9與4.8倍。在正常以及熱誘導條件下的野生型中都沒有檢測到GUS基因的表達,而在pBI121-pMsMBF1c::GUS轉基因植株中,GUS基因顯著受熱誘導,與正常條件相比,表達量增加了5.4倍(圖3)。
2.5 MsMBF1c啟動子熱誘導GUS染色分析
動植物中MBF1家族基因的功能與耐逆性尤其與耐熱調節顯著相關,挑選3個獨立的轉pBI121-pMsMBF1c::GUS株系與野生型擬南芥一起,采用β-葡萄糖醛酸糖苷酶(β-glucuronidase,GUS) 組織化學染色分析其在正常與42 ℃高溫誘導后的表達變化。在正常和高溫誘導條件下,野生型均不能被染色(無GUS基因表達)。在正常條件下, 3個pBI121-pMsMBF1c::GUS轉基因植株呈淡藍色,pMsMBF1c啟動子低豐度驅動GUS表達,42 ℃高溫處理1 h后,3個pBI121-pMsMBF1c::GUS植株呈深藍色,說明pMsMBF1c啟動子顯著受高溫誘導,驅動下游報告基因GUS的表達(圖4)。
3 討論
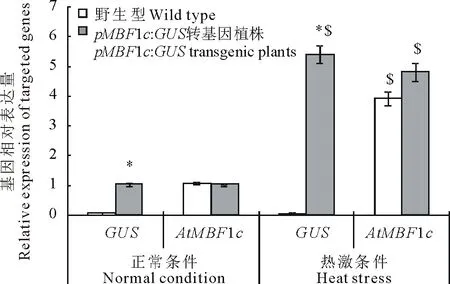
圖3 AtMBF1c與GUS報告基因在正常與高溫脅迫條件下的野生型及轉基因擬南芥中的表達變化分析Fig.3 Expression analysis of AtMBF1c and GUS, the reporter gene, in wild type and transgenic Arabidopsis plants 每個樣品3個生物學重復, Actin 7為內參基因, *表示轉基因植物與野生型之間差異在P<0.05達顯著水平,$表示相同材料在正常和熱激條件下差異在P<0.05達顯著水平。Each sample had 3 biological repeats, Actin 7 were used as an internal reference, *stands for a significant difference at P<0.05 between different materials (wild type and transgenic plants), while $ stands for a significant difference at P<0.05 between different treatment (normal and high temperature stress conditions) within the same material.
植物為光合自養型生物,不能像動物那樣采取移動的方式來應對不利的環境脅迫,因此需要通過調節不同基因的表達以提高自身的適應性[27]。植物基因的表達是由不同啟動子驅動,可以分為組成型、組織特異型和誘導型等幾種,根據生長發育和應對脅迫需要,選擇性的啟動或關閉基因的表達,節約植物新陳代謝所需的能量[1]。在模式植物擬南芥[15, 28]與小麥(Triticumaestivum)[29]等農作物中,MBF家族基因都被報道能夠參與植物耐熱調節,然而還沒有對該基因的啟動子進行研究的報道,因此本研究對紫花苜蓿MsMBF1c的啟動子的熱誘導性進行系統研究,為更好的利用MBF基因進行耐熱性改良奠定基礎。
植物耐熱調控網絡已經有了很好的研究,前人建立了一個以HSF-HSP為核心的耐熱調控網絡[18]。其中受HSF調節的基因在其啟動子區域都存在一個核心的保守順式作用元件HSE[13]。在MsMBF1c啟動子序列中,發現了該功能保守的元件,說明MsMBF1c可能受HSF直接作用參與植物耐熱調節,最近在匍匐剪股穎(Agrostisscabra)耐熱轉錄組中,HSF、HSP與MBF1基因都顯著受熱誘導[30],能夠從側面印證本研究的推論。除了HSF-HSP調控網絡外,植物耐熱途徑還存在第二信使途徑、植物激素調節途徑以及其他調解途徑等,這些途徑之間相互獨立,但也通過一些關鍵基因相互鏈接到一起[31]。最近的研究發現GATA啟動子模體能夠參與植物耐熱調節[19],在MsMBF1c啟動子序列中共有3個拷貝的GATA啟動子模體,說明MsMBF1c的表達可能受耐熱核心基因HSF與GATA相關途徑的耐熱相關基因的共同調節,前人研究發現HSP70能夠通過GATA途徑調控紫杉醇誘導的細胞程序死亡[32],而HSP70也是耐熱調節途徑的關鍵基因[33]。
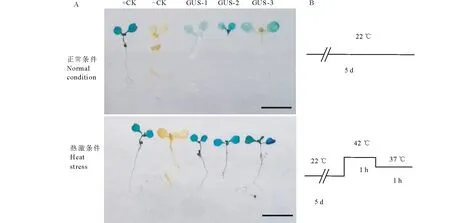
圖4 GUS染色顯示高溫誘導MsMBF1c啟動子驅動報告基因表達Fig.4 GUS staining of pBI121-MsMBF1c::GUS transgenic plants showed the MsMBF1c promoter could be induced by high temperature A:pBI121-MsMBF1c::GUS轉基因植物在正常與高溫條件下報告基因GUS染色結果;B:正常與高溫處理示意圖。+CK: pBI121空載體;-CK: 野生型擬南芥;GUS-1,-2,-3:pBI121-MsMBF1c::GUS轉基因株系-1,-2,-3, 標尺=5 mm。A: GUS staining of pBI121-MsMBF1c::GUS transgenic plants under normal and high temperature conditions; B: A diagram showing the normal and high temperature stress conditions. +CK: pBI121 vector; -CK: Wild type Arabidopsis seedlings; GUS-1,-2,-3: pBI121-MsMBF1c::GUS transgenic plants -1,-2,-3, bars=5 mm.
MBF1c基因不僅能顯著受高溫誘導,而且在部分植物中已經被應用于耐熱性改良[14]。本研究采用熒光定量的方法分析了pBI121-MsMBF1c::GUS轉基因株系和野生型擬南芥中AtMBF1c的表達也能顯著受高溫誘導,與前人的結果一致[34]。熒光定量表達分析MsMBF1c啟動子的熱誘導性發現高溫條件下GUS的表達也顯著的高于正常條件(圖3),組織化學染色(GUS)印證了熒光定量PCR的結果(圖4)。
植物耐逆調控網絡一直都是植物育種與植物功能基因組研究的熱點,并已發現一批功能保守的轉錄因子,它們具有多種功能能夠調節植物耐逆性,這些轉錄因子是連接其他轉錄調控因子、植物激素等信號分子與功能基因的橋梁與紐帶[35],例如MBF家族基因。小麥的TaMBF1c能夠促進水稻和酵母的耐熱性[29]。水稻MBF1不僅能夠調節植物營養生長與滲透脅迫,而且還參與水稻抗稻瘟病調節[36]。南極洲苔蘚(Polytrichichastrumalpinum)MBF1c基因能顯著增強擬南芥對高鹽脅迫的耐受性[37],然而在牧草類作物中,還沒有關于MBF參與其他耐逆性的報道。本研究中,在MsMBF1c啟動子中共發現了5個ABA應答元件(ABRE、MYB2、MIC2、CBF與DPBF),分別參與干旱(水分調節)等環境脅迫調節反應,說明MsMBF1c可能參與了ABA介導的植物耐逆性調節。因為ABA是植物耐逆調控的關鍵激素之一,特別是在抗旱反應中對氣孔的開閉起到至關重要的作用[38]。此外,在MsMBF1c啟動子中還發現了MYB3與MYB4兩個參與冷害與病害調節順式作用元件,說明MsMBF1c除了參與抗旱耐熱反應外,還可能參與其他耐逆性調節。
4 結論
在無參考基因組信息的作物中,酶切連接是一個經濟有效的獲取已知序列側翼序列的方法,對小宗農作物重要基因啟動子序列的獲得與功能分析有重要促進作用。紫花苜蓿MsMBF1c啟動子序列中有多個保守的參與植物耐熱調節及其他抗逆調節的順式作用元件,其可能是紫花苜蓿耐逆(熱)調節的關鍵作用因子。從生物信息分析、基因表達分析與組織化學染色等多個角度說明MsMBF1c啟動子能顯著受高溫誘導激活,因此可作為高溫誘導型啟動子用于科研與生產實踐。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
